Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2014) Volume 2, Issue 1
Probiotic bacteria are proposed for prevention of Clostridium difficile associated diarrhea. The aim of this in vitro study was to evaluate the influence of five Lactobacillus plantarum strains to the survival of C. difficile reference strains (M13042 and VPI 10463) and clinical isolates (n=12) using co-culturing and micro-titre plate assay. Changes in bacterial growth were assessed over the time period of 48 h. Quantitative analysis of C. difficile population revealed that there was a significant decrease of C. difficile in co-culture compared to the control (p=0.01). Susceptibility against L. plantarum was C. difficile strain specific, while L. plantarum was not affected by the presence of C. difficile. Reference strains were more sensitive to inhibition than most of the clinical strains (M13042 strain vs eight clinical strains, p=0.03; VPI vs six clinical strains, p=0.04). Fluoroquinolone resistant C. difficile strains were less inhibited by L. plantarum than sensitive strains (p<0.05). In the micro-titre plate assay experiment the inhibition of C. difficile was not related to any particular C. difficile strains however, inhibitory activity was affected by treatment of supernatants. Supernatants of tested lactobacilli inhibited the C. difficile growth from 72% to 82% if nonneutralized (p=0.001); 43% to 68% if neutralized (p=0.003) and 92% to 99% (p=0.001) if supernatant was neutralized and heated as compared to controls.
Keywords: Clostridium difficile ; Lactobacillus plantarum; Antibiotic associated diarrhea; Probiotics.
Clostridium difficile, an anaerobic toxigenic bacterium, causes a severe infectious colitis that leads to significant morbidity and mortality worldwide. In North America and Europe C. difficile has been a well-established pathogen for decades. Prerequisite for colonisation and infection by C. difficile is weakening of colonization resistance by suppression of indigenous intestinal microbiota, usually due to administration of antibiotics. Both enhanced bacterial toxins and diminished host immune response contribute to symptomatic disease. C. difficile has been a well- established pathogen in North America and Europe for decades [1].
Use of probiotic organisms to reduce and alleviate antibiotic-associated diarrhea has started to receive increasing interest during recent years [2]. Probiotics are live microorganisms that are available over the counter and represent a low-cost, well-tolerated, safe, non-antibiotic based strategy that may have efficacy as adjunctive treatment of infections without the attendant risks of promoting antimicrobial resistance [3-5]. Along with conventional antibiotic therapy, administration of probiotics to manage C. difficile associated diarrhea is drawing increasingly more attention [2,3,6,7]. Since C. difficile infection develops after suppression of indigenous microbiota, restoration of colonization resistance with beneficial bacteria seems to be the most natural way for prevention and treatment of this infection.
Various possible mechanisms of antagonism against C. difficile by lactic acid bacteria have been suggested: pH reduction, competition for nutrition, production of antimicrobial substances, blockage of receptors, immunomodulation etc. [8-12]. The growth of C. difficile may be affected by low pH conditions due to organic acid secretion by lactic acid bacteria. Lactobacilli can also influence the cytotoxicity of C. difficile [13,14].
A great number of in vivo and in vitro studies have been published to evaluate the effect of probiotics against C. difficile; however the results are controversial and only partially successful [2,15-18]. This could be related to individual gut microbiota of macroorganisms and its suppression range, specific virulence factors of particular C. difficile strains and probiotic properties of used lactobacilli.
The aim of our work was to determine the antimicrobial effect of L. plantarum strains to clinical C. difficile isolates.
Material
C. difficile and lactobacilli strains were isolated from Estonian and Norwegian antibiotic-associated diarrhoea patients’ stools and have been described in our previous publications [19,20]. For this experiment, we selected 12 C. difficile strains with different antimicrobial resistance patterns belonging to different PCR ribotypes. Among the selected C. difficile strains 6 strains (N; N1; N2; N3; N4; N5 ) were isolated from Norwegian patients and the other 6 strains (E; E1; E2; E3; E4; E5) from Estonian patients. We also included 2 reference strains: C. difficile VPI 10463 (ATCC 43255) and C. difficile M13042 (epidemic strain from Canada belonging to ribotype 027). We selected 5 strains, 4 strains (N11; N27; N33; N44) were isolated from Norwegian patients and 1 strain (E56) was isolated from an Estonian patient. The lactobacilli strains were selected due to their best antagonistic in vitro activity against C. difficile reference strains in our previous screening study (data not shown). All these lactobacilli belonged to L. plantarum species. Clinical C. difficile strains N; N4; N5; E1; E2 and E5 were wild type (minimal inhibitory concentration of moxifloxacin ranged from 0.25 to 1.0 mg/L) and N1; N2; N3; E; E3 and E4 were resistant to moxifloxacin (minimal inhibitory concentration =32.0 mg/L). Clinical C. difficile strains belonged to ribotypes 077, 020, 012, 087, 046, 126, four C. difficile strains ribotypes were not typable.
Co-culturing of C. difficile and L. plantarum strains
The antagonistic activity of L. plantarum against the growth of C. difficile was determined in co-cultivation assay. The quantity of C. difficile was calculated by serial dilution method. C. difficile isolates were enumerated in triplicates. All possible combinations of different C. difficile and L. plantarum strains were studied.
Briefly, an experimental mixture was made as follows: 50 ml of sterile Brain Heart Infusion (BHI) broth (Oxoid Ltd, UK) was inoculated with 0.05 ml of lactobacilli suspension and 0.05 ml of C. difficile suspension, made of 24 h old cultures with final density equal to McFarland 3.0. In positive control BHI was inoculated similarly as described solely with C. difficile or L. plantarum strains. The co-cultures and positive controls were incubated under anaerobic conditions (Anaerobic box Concept 400, The Baker Company, USA) with gas mixture (85% N: 10% CO2: 5% H2) for 48 h at 37°C.
For detection of C. difficile and L. plantarum quantity during the incubation time serial 10-fold dilution in peptone water to 10-7 the made. Quantification of C. difficile and L. plantarum as performed in the beginning of the experiment (0h) and on the 10th, 24th and 48th hour.
C. difficile populations were enumerated by serial dilution and were inoculated onto Fastidious Anaerobe Agar (FAA, Lab M, UK) supplemented with 2% horse blood and incubated in anaerobic conditions for 48 h at 37°C. L. plantarum populations were enumerated by serial dilution and were inoculated onto Man Rogosa Sharpe Agar (Oxoid Ltd, UK) and incubated in microaerobic environment for 48 h at 37°C. Colonies were counted at the dilution at which 1 to 100 well-separated colonies were visible, and viable counts were expressed as log10 CFU/ml (colony forming units per ml).
The inhibition of C. difficile growth at 48 hours was calculated the following way: difference between C. difficile counts in co-culture at 48 and 0 hours minus difference of C.difficile control culture at 48 hours and 0 hours i.e. growth inhibition by lactobacilli = (C. difficile counts in co-cultures at 48 h – counts at 0 h) – (C. difficile counts in controls at 48 h – counts at 0 h).
Antimicrobial activity of Lactobacillus plantarum culture supernatant against Clostridium difficile strains by a microtitre plate (MTP) assay
The antimicrobial activity of L. plantarum supernatant against C. difficile growth was detected according to Kondepudi et al. (2012) by microtitre plate assay with some modifications [21].
Briefly, BHI broth was inoculated with L. plantarum strains and incubated in microaerobic conditions for 24 h. Extracellular cell free supernatants from these cultures were collected by centrifugation (3000 g x for 15 min). The cell free supernatant’s pH was measured and the supernatant was divided into three equal parts: the first part was left acidic, the second was neutralized with 6N NaOH to pH 6.0 and the third part was neutralized and heated at 100°C for 20 minutes. All the supernatants were filter sterilized (0.2 µM, Orange Scientific, Belgium).
Overnight C. difficile cultures grown on FAA with 2% horse blood supplement were used for the suspension made in BHI broth with a density according to McFarland 3.0. For evaluating the antimicrobial activity of L. plantarum supernatant the following reaction mixes were used: (1) 20 µl of C. difficile cell suspension, 162 µl of Peptone Buffer Salt suspension and 18 µl of BHI broth (as positive control); (2) 20 µl of C. difficile cell suspension, 162 µl of cell free L. plantarum supernatant and 18 µl of BHI broth; (3) 20 µl of C. difficile cell suspension, 162 µl of cell free neutralized L. plantarum supernatant and 18 µl of BHI broth; (4) 20 µl of C. difficile cell suspension, 162 µl of cell free neutralized and heated L. plantarum supernatant and 18 µl of BHI broth. Reaction mixes were incubated under anaerobic conditions for 48 h at 37°C.
The optical density (OD620 nm) was measured in the beginning and at the 48th hour of the experiment. Growth of clostridia (change in optical density values) was measured after 48 h by using an MTP reader (Sunrise Basic, Tecan, Austria). Also growth rates were calculated. The suppressive activity of L. plantarum strains were given as a percentage of inhibition of C. difficile growth, calculated by using the following formula: the % of inhibition of C. difficile growth=100-(ODt X 100/ODc). ODt and ODc are growths of C. difficile in the presence and absence of L. plantarum [21].
Statistical analysis
The suppressive activity of L. plantarum strains and different L. plantarum supernatants were tested with Friedman`s test. All the data was expressed as averages. Changes in C. difficile growth (counts) after 10 h, 24 h and 48 h of incubation were compared by Wilcoxon (signed-rank) test. C. difficile growths (counts) after 48 h of incubation in pairs (L. plantarum+C. difficile vs C. difficile) were compared by Friedman test. The statistical analysis was performed by using Stata program (StataCorp LP, USA).
Co-culturing of C. difficile and L. plantarum strains
The growth of lactobacilli was not affected by the presence or absence of C. difficile, as the counts of L. plantarum in all of the time points (10 h, 24 h, 48 h) were similar in co-cultures as well as in positive control (Table 1). There was no difference in inhibitory activity between the strains of five L. plantarum tested (data not shown). There were differences in counts of C. difficile in co-cultures vs positive control after 48 h but no statistically significant differences after 10 h and 24 h of incubation (Table 1). Susceptibility against L. plantarum antimicrobial activity was C. difficile strain specific. Some clinical strains (E2, E3, E5) were highly sensitive to inhibition of L. plantarum however, some (N1, N2, N3, N4, E4) were inhibited only minimally (p=0.03). Reference strains were more sensitive to inhibition than most of the clinical strains: M13042 strain vs N1-N4; E2-E5 strains (p=0.03) and VPI 10463 strain vs N-N4; E; E4 strains (p=0.04).
| L. plantarum or C. difficilecounts (CFU log10/ml median (range) at different incubation times | |||||
|---|---|---|---|---|---|
| 0 h | 10 h | 24 h | 48 h | ||
| L. plantarumn=5 | Control: LP growth alone | 6.0 (5.2-6.3) |
7.7 (6.3-9.0) |
8.2 (8.0-10.3) |
7.0 (6.0-8.5) |
| Co-culture: LP growth with CD | 5.8 (5.0-7.8) |
7.8 (6.0-9.6) |
8.3 (7.0-11.0) |
7.1 (6.0-9.0) |
|
| C. difficilen=14 | Control: CD growth alone | 4.1 (2.0-6.0) |
6.7 (3.7-8.6) |
5.0* (4.0-9.0) |
7.0# (5.8-8.1) |
| Co-culture: CD growth with LP | 4.0 (2.0-6.3) |
5.0 (2.0-9.3) |
2.0* (2.0-6.5) |
3.0# (2.0-7.1) |
|
Table 1: Growh of L. plantarum and C. difficle strains in different conditions (alone and co-culture) at different incubation times. CD - C. difficile ; LP - L. plantarum. *p=0.058; #p=0.01
Fluoroquinolone resistant C. difficile strains were less inhibited by L. plantarum than the sensitive ones, medians (range): 0.3 (-3.7 to 4.1) vs -0.8 (-4.3 to 2.4); p<0.05 (Figure 1).
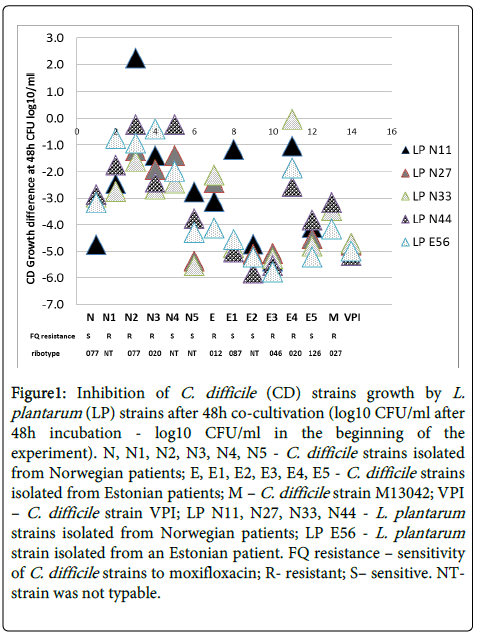
Figure 1: Inhibition of C. difficile (CD) strains growth by L. plantarum (LP) strains after 48h co-cultivation (log10 CFU/ml after 48h incubation - log10 CFU/ml in the beginning of the experiment). N, N1, N2, N3, N4, N5 - C. difficile strains isolated from Norwegian patients; E, E1, E2, E3, E4, E5 - C. difficile strains isolated from Estonian patients; M – C. difficile strain M13042; VPI – C. difficile strain VPI; LP N11, N27, N33, N44 - L. plantarum strains isolated from Norwegian patients; LP E56 - L. plantarum strain isolated from an Estonian patient. FQ resistance – sensitivity of C. difficile strains to moxifloxacin; R- resistant; S– sensitive. NTstrain was not typable.
Antimicrobial activity of L. plantarum supernatant against C. difficile strains
Optical density in the control group and experimental group did not differ after 10 h and 24 h of incubation. After 48 h of incubation optical density in the control group was significantly higher than in all groups with L. plantarum supernatants (p=0.01).
In this experiment inhibition was not related to particular C. difficile strains however, inhibitory activity was affected by treatment of supernatants. Supernatants of tested lactobacilli inhibited the C. difficile growth from 72% to 82% if non-neutralized (p=0.001); 43% to 68% if neutralized (p=0.003) and 92% to 99% (p=0.001) if supernatant was neutralized and heated as compared to controls (Table 2).
| Lactobacillus plantarum strains | |||||
|---|---|---|---|---|---|
| Supernatant of lactobacilli | N11 | N27 | N33 | N44 | E56 |
| Natural (acidic) | 82% | 72% | 76% | 74% | 75% |
| Neutral (pH 6.0 ± 0.15) | 48%* | 67% | 43% | 61% | 68%# |
| Neutral, heated (for 20 min, 100°C) | 99%*× | 97% | 94%× | 92% | 97%# |
Table 2: Inhibition of different Lactobacillus plantarum supernatants to growth of Clostridium difficile strains after 48 h. * p=0.01; # p=0.04; × p=0.001
The highest inhibitions of C. difficile growth were in case of heated neutralized supernatant and the lowest in case of neutralized one: heated neutralized average 96±3% vs non-neutralized 76±4% (p=0.04) and non-neutralized 76 ± 4% vs neutralized 57 ± 11% (p=0.04). There was statistically higher inhibition in heated neutralized supernatants vs neutralized supernatants of N11 and E56 lactobacilli strains. When comparing antagonistic activity of L. plantarum strains, there was relevant difference only between N11 vs N33 strains in case of heated neutralized supernatants (Table 2).
We found that L. plantarum strains were able to inhibit the growth of C. difficile in vitro. L. plantarum is part of indigenous microbiota, but its prevalence in gut may vary in different geographical areas [22]. In our previous study we found some correlation between the absence of L. plantarum strains and presence of C. difficile in the intestinal tract of patients with antibiotic associated diarrhoea [19].
L. plantarum is able to grow in many different niches and is important for different food and health applications. For example, it is one of the dominant species in fermented foods- sauerkraut, olives, sourdough, and kimchi [23,24]. These bacteria are also applied to preservative processes where they, like many other lactic acid bacteria, can contribute to the production of antimicrobial substances (organic acids, bacteriocins) [25]. Genome sequencing and comparative genomics have revealed a high genomic diversity and flexibility of L. plantarum, which can contribute to its success in diverse niches and applications. L. plantarum, as other lactic acid bacteria, has mosaic modules or cassettes of carbohydrate utilization genes, but L. plantarum seems to be very good in acquiring and shuffling these cassettes and it also allows the optimization of its genome for growth in specific niches [26].
L. plantarum is also used as a probiotic. There have been a growing number of studies about the potential beneficial effects of L. plantarum strains on human health [27]. Although exact mechanisms of these effects are still not defined, some of these could contribute to immunomodulation of the host, competitive exclusion of pathogens, production of antimicrobial substances including bacteriocins and antioxidants [28,29].
In our broth co-cultivation assay for detection of antagonistic activity of several L. plantarum strains, some C. difficile strains were more sensitive to this inhibition than others. This result is generally in concordance with our previous study where C. difficile strain specific inhibition by different lactobacilli species was detected by using growth inhibition assay on agar plates [17]. However, in our previous study C. difficile strains, which were more sensitive to lactobacilli were usually more resistant to various antibiotics (chloramphenicol, tetracycline, rifampicin and erythromycin). On the contrary, in the present study strains resistant to fluoroquinolones were less sensitive to lactobacilli. Relations between these properties are unclear and need future studies.
The variable sensitivity to lactobacilli is one possible explanation of high variation of clinical presentations of C. difficile infections from asymptomatic colonization to lethal disease. This variation could be related to the alteration extent of indigenosus microbiota (presence or absence of particular lactobacilli) on one hand and properties of C. difficile strain (sensitivity to lactobacilli) on the other hand. The strain specific sensitivity of C. difficile could also be one reason for contradictory results of usage of probiotics against clinical or experimental C. difficile infection, since the effect of probiotic strain could be dependent on properties of particular C. difficile strain causing infection [2,15-18].
It should also be taken into account that most of in vitro and animal experiments were done with a few reference strains such as VPI and epidemic 027, which were highly sensitive to lactobacilli according to our results [30,31]. Since clinical C. difficile strains could be more resistant, effect of lactobacilli in clinical settings could be weaker than in experimental studies.
In our inhibition assay with culture supernatant, we found no C. difficile strain-specific effects. Thus, different in vitro assays can give different results and probably in co-cultivation assay other mechanisms beside lactobacilli produced compounds are involved. The neutralization of supernatant did not reduce its inhibitory effect. Thus, lowering the pH of the environment is not the main mechanism in inhibition of C. difficile by lactobacilli. Also heating of supernatant did not reduce its activity thus; some thermostabile compounds may be involved in the inhibition. It is known that L. plantarum strains can produce several thermostabile plantaricins, which inhibit mostly closely related species (L. monocytogenes, other lactobacilli) but can also inhibit several pathogens such as S. aureus, C. perfringens, B. subtilis, E. coli, S. typhimurium [32-34]. Production of plantaricins by our antagonistic strains and their role in inhibition of C. difficile should be evaluated in future studies.
In conclusion, the protective effect of probiotics could vary in different C. difficile infected patients and may depend on the properties of a particular C. difficile strain causing infection. Since sensitivity of C. difficile to lactobacilli is strain-specific, several different C. difficile strains should be included to experimental studies to avoid strain related biases.
This study was supported by grants of Stavanger University Hospital (Project no 500808), Europe financial mechanisms and Norway financial mechanism (EMP 13), Estonian Ministry of Education and Research (target financing No SF0180132s08 and No KOGU-HUMB), Estonian Science Foundation (grant No 7933 and No 9059) and University of Tartu (grant No SARMBARENG). We thank Karolin Toompere for statistical analysis of data.