Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2013) Volume 2, Issue 1
Consumption of untreated drinking water, raw milk, undercooked poultry meat and handling of raw poultry meat are the main sources of campylobacteriosis. Treatment of campylobacteriosis becomes very difficult due to the increasing pattern of antimicrobial resistance of these microbes. Thus, objectives of this study were to determine the cytotoxic potential of Campylobacter jejuni (C. jejuni) isolated from fecal samples of poultry on Chinese Hamster Ovary cell (CHO) and also to determine the effect of Bitter Melon (Momordica charantia) extract (BME) at 25%, 5%, 2.5% and 0.5% concentrations to reduce the cytotoxic effect of C. jejuni on CHO cells. Thirty-two C. jejuni strains isolated from poultry fecal samples were analyzed using Bolton broth as pre-enrichment broth and modified Charcoal–Cefoperazone Deoxycholate agar as a selective media, then characterized for the presence of 16S rRNA gene using polymerase chain reaction. Of the thirty-two strains isolated, twelve (37.5%) were positive for the presence of 16S rRNA gene. Minimum percent (0.17%) of cell adhesion of C. jejuni strain ATCC 29428 on CHO cells was observed at 25% BME and highest percent (8.3%) of cell adhesion was observed at 0.5% BME C. jejuni strain FCA 40 released minimum percent (44%) of Lactate Dehydrogenase (LDH) from CHO cells in the presence of 25% concentration of BME, where as maximum percent (73%) of LDH release was observed with 0.5% concentration of BME on CHO cells. Results of this experiment also showed that percent alkaline phosphate (AP) release is ranged from 100% (ATCC 29428 with 0.5% BME) to 67% (ATCC 29428 with 25% BME). Thus, results of this study showed that BME significantly reduced the cytotoxic effects of C. jejuni on CHO cells, and it can be utilize as a natural therapeutic agent.
Keywords: Foodborne; Campylobacter; jejuni; Chinese hamster ovary cell; Bitter melon extract; Cytotoxic effect Foodborne
Campylobacteriosis is one of the most common diarrheal illness in the United States caused by Campylobacter spp. Campylobacteriosis is estimated to affect over 2.4 million persons or 0.8% of the population and causes 124 deaths each year in the US [1]. CDC reported that approximately one in every 1,000 reported Campylobacter illnesses leads to Guillain-Barré syndrome and as many as 40% of Guillain- Barré syndrome cases may be triggered by campylobacteriosis in the US (CDC, 2010). This infection may be sporadic in occurrence or can occur as an outbreak usually in autumn and spring [2]. Sporadic infection occurs due to consumption of untreated drinking water [3], raw milk [3,4], milk from bottles pecked by birds [5], undercooked poultry [3,6-9], and handling of raw poultry meat [3,10]. C. jejuni causes more than 90% of campylobacteriosis cases in most developed countries [11]. The species most commonly associated with human infection is Campylobacter jejuni (80–90%), followed by Campylobacter coli (5–10%), but other Campylobacter species are also known to cause human infection [12,13]. It has been reported that overlapping of serotypes of C. jejuni was observed in humans, poultry and cattle, which indicated that food of animal origin have a significant contribution in transmitting Campylobacter jejuni to human being [14].
Campylobacter species has resistance to several antibiotics, which leads to increasing infection and prolonged illness in humans. This resistance to several antibiotics makes it difficult to treat the infection. The use of fluoroquinolones in poultry was approved in Europe, but there was a rapid increase in the C. jejuni strains in humans in early 1990s [15,16], which showed that the fluoroquinolones susceptible strains also became resistant when it was administered in chicks [17]. The strains, which were isolated from dysenteric samples, were found to be more invasive and cytotoxic than the strains which were grown by in vitro method [18]. The important step in the pathogenesis of Campylobacteriosis is reported to be the adhesion to and invasion of the surface of the mucosal surfaces [19]. C. jejuni does not exhibit invasive property when tested with Sereny-Anton test. However, its invasive property is proved by its presence in blood and leucocytes of infected patients and also through many experiments performed in vivo [20].
C. jejuni is believed to account for approximately 90% of all cases of human Campylobacter infections, with C. coli, which is usually associated with pigs, accounting for 3-4% of cases [21]. Case-control study in the United States has shown that 48% to 70% of sporadic infections were due to consumption of chicken infected with C. jejuni [22]. However, consumption of contaminated chicken may not only be the reason for Campylobacteriosis in human. Feces of beef cattle are also known to be a rich source of C. jejuni. The infection can also occur due to drinking of contaminated water with C. jejuni [23]. The disease occurs due to the consumption of enterotoxins produced in the contaminated food by C. jejuni. Within 2-3 days of consumption of contaminated food, symptoms such as fever, abdominal pain and diarrhea are observed followed by the gastroenteritis, septicemia, proctitis, arthritis, meningitis, abortion and Guillain-Barre’ syndrome [23]. Treatment for Campylobacteriosis becomes difficult because of the antimicrobial resistance of these microbes. Antimicrobial resistance in Campylobacter is an emerging threat to public health. In a study, 38.8% of 297 C. jejuni samples were resistant to antibiotics like ampicillin, streptomycin, and erythromycin samples [8]. In the same study resistance to tetracycline, a character transferred through plasmid was the most common antibiotic resistance observed in around 23.8% of all the C. jejuni. In another study, 59% of C. jejuni were found to be resistant to ciprofloxacin, 23% to tetracycline and 17% to ampicillin from 69 samples collected from humans suffering with diarrhea [24].
Differentiation between various species of C. jejuni is very important, especially between C. jejuni and C. coli. The main reason is that these species are the most frequently isolated from the diarrhea. These two species share common clinical manifestation, which makes it further more difficult to differentiate. Microbiological, biochemical and serological detection and identification are time consuming and cumbersome procedures. Thus, an easier and efficient way of differentiating microbes with close similarities is by performing molecular identification.
Adherence and invasion of Campylobacter species into a host cell does not mean that host cell is infected. After the adherence and invasion of C. jejuni, the host cell defense will try and kill the microbe. A host cell is said to be infected either by observing derailment in metabolic changes or by the cell damage or cell death. Cell death or damage is indicated by the release of various enzymes such as lactate dehydrogenase and alkaline phosphatase. Thus, in this study cell association, adhesion and cell cytotoxicity assays were performed by estimating the lactate dehydrogenase and alkaline phosphatase release from Chinese hamster ovary cells. Cytotoxic effects of various Campylobacter jejuni strains isolated from cells fecal samples of poultry in the presence of 25, 5, 2.5 and 0.5% ethanol extract Bitter Melon.
The C. jejuni samples were collected from various poultry farms using sterile tongue depressors. Approximately 2-3 g of fecal samples was collected into tubes containing Clair-Blair semi-solid medium. This medium were supplemented with 5% lysed horse blood and shipped in refrigerator maintained at 8-10ºC to the Food Microbiology Laboratory at Alabama A&M University.
Isolation and Growth of Campylobacter jejuni
Within an hour of sample collection, plating was performed for identification of C. jejuni. Plating was performed using modified Campy- Cefex [25] and modified charcoal cefoperazone deoxycholate agar [26]. The samples were grown in Preston Broth enrichment media (Oxoid, Ltd.), and then were incubated for 24 h at 42ºC under microaerophillic conditions. Microaerophillic condition was provided in Anaxomat (Spiral Biotech, Norwood, MA) jar glassing System. These conditions include 10% CO2, 5% O2, and 85% N2 air gas mixture. Then 0.1 ml of this enriched sample were swabbed onto mCC (modified campy cefex) and mCCDA (modified charcoal cefeperozone deoxycholate agar) plates. These plates were incubated for 24 h at 42ºC under microaerophillic conditions. Plates were then screened for typical Campylobacter colonies. Colonies that showed typical morphology and motility under phase contrast microscopy were considered presumptive positive. Tryptic soy broth (Difco, Detroit, MI) supplemented with 30% glycerol (v/v) and 5% lysed horse blood were used to store the presumptive positive samples at -80ºC for further use.
In this study, C. jejuni strains were isolated using modified protocol. Brain heart infusion broth (BHI) supplemented with crude 0.5, 2.5, 5.0, and 25% Bitter Melon extract were used for culturing all C. jejuni strains. The ethanol extract of Bitter Melon used in this experiment was provided by Dr. S. Rao Mentreddy, Medicinal Plant Laboratory of Alabama A&M University. Different concentrations of Bitter Melon extract were prepared using sterile distilled water. BHI broth (5 mL) in a 13×100 mm culture tube was inoculated with approximately 10 μl of culture taken from a frozen glycerol stock. Each tube containing the starter culture was incubated at 37ºC for 24-48 h without agitation under modified microaerophillic conditions (85% Nitrogen, 10% carbon dioxide and 5% oxygen).
Polymerase Chain Reaction Identification of Campylobacter Isolates
DNA from the samples was extracted. The samples were initially grown for about 24-48 h at 42ºC under microaerophillic conditions. DNA extractions were performed using PrepMan™ Ultra (Applied Biosystems, Foster City, CA). PCR assay were carried out using extracted DNA samples for 16s ribosomal RNA. 16s ribosomal RNA gene is highly conserved within a species and among species of the same genus, and hence can be used as the new gold standard for speciation of C. jejuni. [27].
DNA amplification for 16s ribosomal RNA
DNA amplification for 16S r RNA was performed on a total of 50μl. The reaction mix consisted of 30.9 μl of deionized water, 5 μl of 10X PCR buffer II (50mM KCl, 10mM Tris-HCL, pH 8.3), 4 μl of 25 mM MgCl2, 4 μl of each of 200 μM deoxynucleotide triphosphates dATP, dCTP, dGTP and dTTP, 0.3 μl of each forward and reverse primer (Table 1), 0.5 μl of Taq DNA polymerase (Perkin-Elmer, Norwalk, Conn.) and 5 μl of 20 ng template DNA. Amplification was carried out with in 30 cycles. Denaturation was carried out at 94ºC for one minute, annealing for one minute at 60ºC, and an extension for 1 minute at 72ºC. After the last cycle, an extra extension temperature of 72ºC for 5 minutes was carried out and the samples were held at 4ºC until it is used. The PCR product was detected by electrophoresis on a 2% agarose gel and then visualized under UV light after staining with ethidium bromide.
| Primer Name | Sequence (5'-3') | Gene | Base pair |
|---|---|---|---|
| C412 F | GATGACACTTTTCGGAG | 16SrRNA | 816 |
| C1228 R | CATTGTAGCACGTGTGTC | 16SrRNA | 816 |
Table 1: Gene sequence of Primer used for 16S r RNA for performing Polymerase Chain Reaction.
Cell Association/Adhesion Assay
The cell association assay was performed as described by Francis et al. [28]. CHO cells were seeded in 24-well tissue culture plates with 0.5 ml of Dulbecco,s Modified Eagle Medium (DMEM) medium supplemented with 10% fetal bovine serum containing 105 cells per ml and incubated overnight at 37ºC with 7% CO2. Prior to cell association assay, the medium covering the cells was replaced with fresh pre-warmed DMEM-10 and incubated for a minimum of 30 min at 37ºC in 7% CO2. The cells were inoculated with the same numbers of bacteria for different Bitter Melon concentrations. One hundred micro liters of each culture was added to each well at a multicity of infection (MOI) of 100:1. The initial concentrations per well ranged from 3.1×107 to 5.8×107 cells per milliliter. The plates were incubated for 2 hour at 37ºC with 7% CO2. After incubation, monolayers were washed three (3x) times with sterile phosphate buffered saline (pH 6.8) to remove any non-associated bacteria. The cells were lysed by incubation at 25ºC with 1% Triton-X for 10 minute. BHI broth (0.8 ml) was added to each well and serial dilutions were performed to determine cell concentration per monolayer. This represented the number of bacteria cell-associated and intracellular. Cell associated bacteria were plated onto plate count agar (PCA) to determine the number of associated cells. Cell association percentages were calculated as follows:
(Number of cell associated bacteria / the number of bacteria originally added)×100.
Cell Invasion Assay
The cell invasion assay was carried out as described by Durant et al. [29]. Bacteria were allowed to adhere to CHO cells as described in the cell association assay. After incubation for 2 hour, the overlying medium was removed and replaced with pre-warmed DMEM-10 containing 100 μg of gentamicin per ml. The cell monolayer was washed 3 times with sterile PBS (pH 6.8). The cells were then lysed by incubation at 25ºC with 0.2 ml of 1% Triton-X for 10 min. Brain heart infusion (BHI) broth was added to each well and serial dilutions were performed to determine the cell concentration (cells/ml) per monolayer. This represented the number of bacteria that were internalized since gentamicin killed extracellular bacteria while intracellular bacteria were viable MacBeth and Lee [30]. Intracellular bacteria were enumerated by plating for viable bacteria onto plate count agar. The percent of invasion were determined as follows:
% Invasion = Mean number of internalized bacteria/Mean number of bacteria originally added × 100
Lactate Dehydrogenase Assay
After the development of a monolayer, CHO obtained in 24 well plates were washed two times with serum free Dulbecco’s modified Eagles medium (DOF) and exposed to C. jejuni bacteria at MOI of 100:1 for 16 hour. The experiment was carried out in replicates of 2 wells per treatment. After incubation, cells were centrifuged (300×g, 3 min) using a Beckman bench top centrifuge and cell free supernatant fluid (0.1 ml) was collected and LDH assay was conducted using a cytotoxicity kit (Boehringer Mannheim, Anaheim, CA) according to Manufacturer instructions. After 3 minutes of incubation at room temperature, the plates were read at 490/655 nm using a BioTek Synergy™ (Winooski, VT) micro plate reader. The cytotoxicity percentage was calculated by following formula:
Cytotoxicity (%) = Experimental Value – Low Control/High Control-Low Control×100.
Alkaline Phosphatase Activity
After the development of a monolayer, CHO obtained in 24 well plates were washed two times with serum free Dulbecco’s modified Eagles medium (D0F) and exposed to C. jejuni bacteria for 16 hour. After incubation, cells were centrifuged (300×g, 3 min) and cell supernatant (0.1 ml) was placed in a 96-wells microtitre plate for AP assay. Alkaline phosphatase substrate buffer (100 μl, 0.1 M Tris, 0.1 mol l-1 NaCl, 5 mmol l-1 MgCl2, pH 9.5) containing p-nitrophenyl phosphate (PNPP, 1 mg/ml) (Sigma Chemical, St. Louis, MO) was added to each well. After 3 minutes of incubation at room temperature, the plates were read at A405/595 nm using a BioTek Synergy™ plate reader (Winooski, VT). The AP percentage was calculated by the following formula.
AP % = Experimental Value – Low Control/High Control-Low Control×10.
Statistical Analysis
Complete Random Block Design was used for cell association, cell invasion, lactate dehydrogenase and alkaline phosphatase assay. Data were analyzed using SAS software. Mean differences were tested for statistical significance by ANOVA and means were separated using Tukey’s studentized range test. Means differences were considered significant at P ≤ 0.05.
Bacterial Strains
The ability of the C. jejuni to utilize catalase, oxidase and production of sodium hippurate were determined. The results of this test showed that twelve out of thirty-two isolates were positive for C. jejuni (Figure 1) and these isolates were considered presumptive positive and were further characterized genotypically using PCR.

Figure 1: Detection of 16S rRNA gene from Campylobacter jejuni isolated from poultry samples using polymerase chain reaction. Lanes from left to right are M-100 bp DNA ladder, 1)-Campylobacter jejuni ATCC 29428 (positive control), 2-6) E. coli 0157:H7 (negative control), 10-3, 1, 10-2, 10-5, 14-2 7) Campylobacter jejuni isolate #17-1, 8-9) 7, 10-1 10)- Campylobacter jejuni isolate #14-5, 11)- Campylobacter jejuni isolated #1-3 and 12) Campylobacter jejuni isolate # 8, 14 and 15) 11-4 and 4.
PCR identification
Twelve (38.7%) isolates were positive by PCR for 16s rRNA, indicating the presence of the gene encoding a highly conserved region of the Genus Campylobacter. These isolates positive for the 16s rRNA gene were further characterized for their ability to produce cytotoxic effects on CHO cell and also determine the role of Bitter Melon extract on the virulence and cell mediated cytotoxicity of C. jejuni in cell culture.
Cell adhesion assay without Bitter Melon extract
Results of this experiment showed that percent cell adhesion ranges from 0.34-31.9% by the C. jejuni isolates 14-5 and 17-1 respectively (Figure 2), when inoculated on CHO cells. Highest percentage of cell adhesion was observed in C. jejuni isolates 17-1 (31.9%) followed by the isolate 2-F (31%) and then 5A (20.7%). Lowest percentage of cell adhesion was observed in C. jejuni isolate 14-5 (0.34%). Control strain of C. jejuni ATCC 29428 showed 12.2% cell adhesion. C. jejuni isolates 1230 and 950 showed similar percentages of cell adhesion (8.6%). Thus, these results showed that C. jejuni isolates 17-1 and 14-5 had the maximum and minimum capacity to adhere the CHO cells respectively (Figure 2).
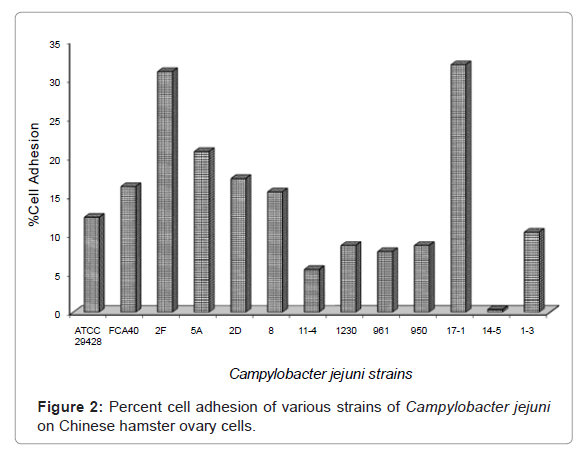
Figure 2: Percent cell adhesion of various strains of Campylobacter jejuni on Chinese hamster ovary cells.
Cell invasion assay without Bitter Melon extract
Outcomes of this study showed that percent of cell invasion by C. jejuni isolates ranged from 0.2%-5.2%, when inoculated on CHO cells (Figure 3). Highest percent of cell invasion was observed by C. jejuni ATCC 29428 (5.2%), then isolate 5A and 961 (2.6%). C. jejuni isolates 8, 11-4, 17-1 and 1-3 showed the least cell invasion (0.2%) on CHO cell (Figure 3).
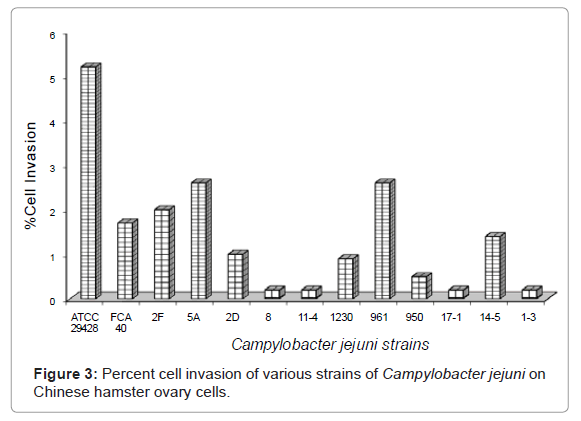
Figure 3: Percent cell invasion of various strains of Campylobacter jejuni on Chinese hamster ovary cells.
Cell adhesion with Bitter Melon extract
Cell adhesion assay was performed using C. jejuni ATCC 29428 and FCA 40 isolate on CHO cells in the presence of Bitter Melon extract at 25, 5, 2.5 and 0.5% concentrations. Minimum percent of cell adhesion of C. jejuni isolate ATCC 29428 on CHO cells was observed at 25% concentration of Bitter Melon extract (0.17%), where as Bitter Melon extract at 0.5% concentration showed high percent of cell adhesion (8.3%) (Figure 4). FCA 40 isolate of C. jejuni showed minimum percent of cell adhesion on CHO cells of 2.6% at 25% concentration of Bitter Melon extract and highest at 0.5% concentration of Bitter Melon extracts (8.8%) (Figure 4). C. jejuni ATCC 29428 and FCA 40 grown in the presence of 25% Bitter Melon extract showed minimum cell adhesion on CHO cells, which indicates that Bitter Melon extract had adverse effects on the growth of C. jejuni and also help to reduce the infectious capability of the organism.
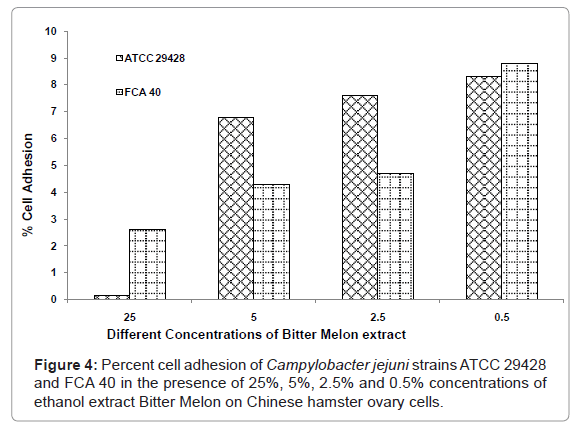
Figure 4: Percent cell adhesion of Campylobacter jejuni strains ATCC 29428 and FCA 40 in the presence of 25%, 5%, 2.5% and 0.5% concentrations of ethanol extract Bitter Melon on Chinese hamster ovary cells.
Cell invasion with Bitter Melon extract
Cell invasion assay was performed using C. jejuni ATCC 29428 and FCA 40 isolate on CHO cells in the presence of Bitter Melon extract at 25, 5, 0.5 and 0.5% concentrations. Complete inhibition of cell invasion was observed in C. jejuni ATCC 29428 grown at 25% concentration of Bitter Melon extract. Results of the this study indicated that there was an increasing trend of invasion of C. jejuni ATCC 29428 isolate with the decreased percent of Bitter Melon extract in which it was grown (Figure 5). FCA 40 isolate of C. jejuni grown at 25% concentration of Bitter Melon extract did not show any cell invasion into CHO cells, but FCA 40 isolate grown at 0.5% concentrations of Bitter Melon, showed 5.3% of cell invasion into CHO cells (Figure 5).
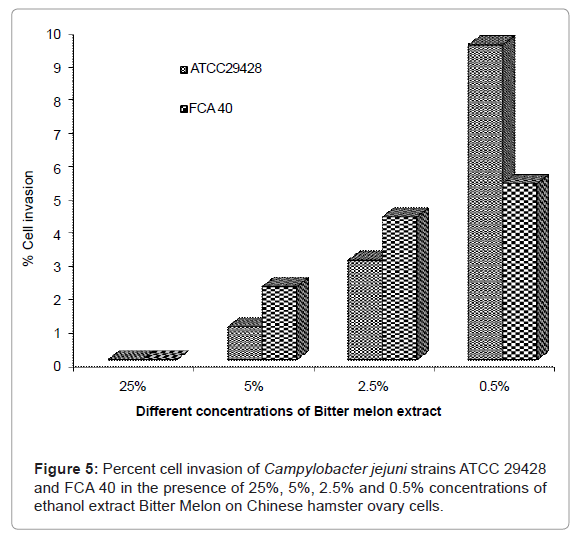
Figure 5: Percent cell invasion of Campylobacter jejuni strains ATCC 29428 and FCA 40 in the presence of 25%, 5%, 2.5% and 0.5% concentrations of ethanol extract Bitter Melon on Chinese hamster ovary cells.
Lactate dehydrogenase assay without Bitter Melon extract
After 16 hour of exposure of CHO cells to C. jejuni ATCC 29428 and FCA 40, LDH assay was performed using a cytotoxicity kit (Boehringer Mannheim, Anaheim, CA). Figure 6 shows the percent LDH released from CHO cells, when treated with various strains of C. jejuni. Maximum percent of LDH release was caused by C. jejuni ATCC 29428 (99%) followed by FCA 40, 8 and 17-1 (91%, 74% and 70% respectively). The lowest percent of LDH enzyme release was caused by C. Jejuni isolates 2F and 14-5 strains (28%). Thus, the strains C. jejuni ATCC 29428 has the ability to cause maximum cell damage releasing highest percent of LDH enzyme followed by FCA 40, 8 and 17-1 (Figure 6). The outcomes of this experiment also showed that C. jejuni strains 2F and 14-5 have the minimum ability to cause CHO cell damage.
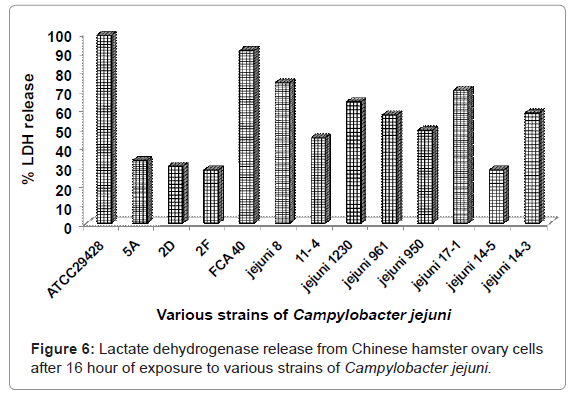
Figure 6: Lactate dehydrogenase release from Chinese hamster ovary cells after 16 hour of exposure to various strains of Campylobacter jejuni.
Lactate dehydrogenase assay with Bitter Melon extract
After 16 hour of exposure of CHO cells to C. jejuni ATCC 29428 and FCA 40 strains grown in the presence of the various concentrations of Bitter Melon, LDH assay was performed using LDH kit. As shown in figure 7, the values of percent LDH release were observed ranging from 97% (ATCC 29428 with 0.5 % Bitter Melon extract) to 61% (ATCC 29428 with 25% Bitter Melon extract). LDH release was lowest percent in the CHO cells treated with 25% of Bitter Melon extract and highest in the CHO cells treated with 0.5% Bitter Melon extract. C. jejuni strain FCA 40 was observed to have minimum LDH release with 25% concentration of Bitter Melon extract (44%) and maximum LDH release with 0.5% concentration of Bitter Melon extract (73%) on CHO cells (Figure 7). The cytotoxic effects of C. jejuni ATCC 29428 and FCA 40 was inhibited due to the presence of Bitter Melon extract at 25% concentration, which resulted in reduce cell damage and less percent of LDH released.
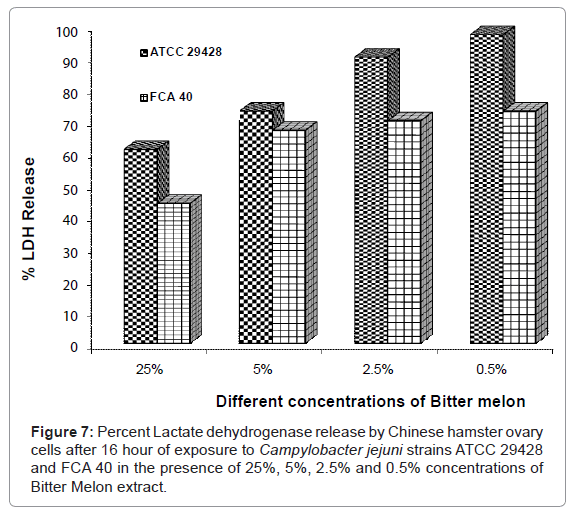
Figure 7: Percent Lactate dehydrogenase release by Chinese hamster ovary cells after 16 hour of exposure to Campylobacter jejuni strains ATCC 29428 and FCA 40 in the presence of 25%, 5%, 2.5% and 0.5% concentrations of Bitter Melon extract.
Alkaline phosphatase assay without Bitter melon
After 16 h of exposure of CHO cells to C. jejuni ATCC 29428 and FCA 40, AP assay was performed using AP substrate at 405/595 nm wave length. Both the strains were observed to have 100% AP release from CHO cells, indicating that both the strains are highly pathogenic and cause complete cell damage.
Alkaline phosphatase assay with Bitter Melon extract
AP assay was performed using CHO cells in the presence of 25, 5, 2.5 and 0.5% concentration of Bitter Melon extract. After 16 h of exposure of CHO cells to C. jejuni ATCC 29428 and FCA 40 grown in the presence of the above mentioned concentrations of Bitter melon, AP assay was performed using AP substrate at 405/595 nm wavelength.
Results of this experiment showed that percent AP release was ranged from 100% (ATCC 29428 with 0.5 % Bitter Melon extract) to 41% (ATCC 29428 with 25% Bitter Melon extract) (Figure 8). Percent AP released from CHO cells was lowest treated with 25% Bitter Melon extract and highest in the CHO cells treated with 0.5% Bitter Melon extract.
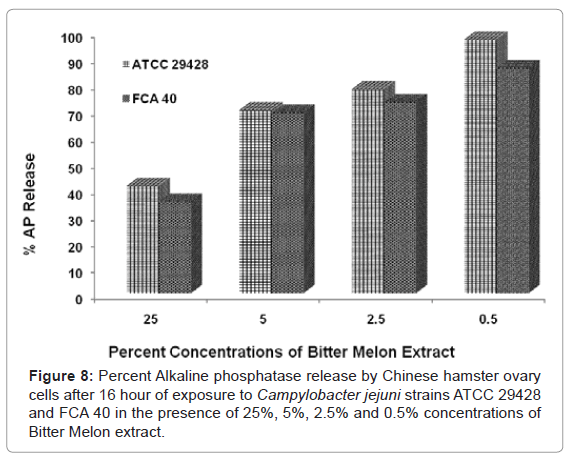
Figure 8: Percent Alkaline phosphatase release by Chinese hamster ovary cells after 16 hour of exposure to Campylobacter jejuni strains ATCC 29428 and FCA 40 in the presence of 25%, 5%, 2.5% and 0.5% concentrations of Bitter Melon extract.
According to World Health Organization, Campylobacter spp. is currently the bacterial foodborne pathogen causing the highest number of gastrointestinal disease in developed countries (www.who.int). Campylobacter jejuni is the most common zoonosis and the main cause of bacterial gastroenteritis in the western world [31]. Several reports showed that annual C. Jejuni infection is about 1% of the US and EU populations [32,33]. Ogundipe et al. [34] reported that the phytochemical extract from the morphological parts of Mallotus oppositifolium exhibited minimum inhibitory concentration and minimum bacterial concentrations of 32.5 micro g/ml and 65 micro g/ ml against Pseudomonas aeruginosa NCTC 6750 and 25 micro g/ml and 50 micro g/ml against Staphylococcus aureus NCTC 6571 respectively [35]. These values obtained were similar to the therapeutic drugs. Mahony et al. [35] demonstrated that turmeric was the most effective against Helicobacter pylori, followed by ginger, cumin, chili, borage, black caraway, oregano and liquorice. Extract of turmeric, borage and parsley showed the ability to inhibit the adhesion of Helicobacter jejuni to the stomach wall. Thus, in this study it observed that the consumption of plants, which have the ability to prevent adhesion of Helicobacter pylori to the stomach wall, would be an excellent alternative for the drugs therapy during infections [35].
Results of this experiment also support the above findings. When CHO cells were infected with C. jejuni without any phytochemicals, the percent cell adhesion ranges from 0.34–31.9% by the C. jejuni isolates 14-5 and 17-1 respectively. Control strain of C. jejuni ATCC 29428 showed 12.2% cell adhesion. When CHO cells were infected with C. jejuni and treated with different concentrations of Bitter Melon extract, it substantially reduced the cell adhesion. When CHO cells were infected with control strains of C. jejuni ATCC 29428 and FCA 40 and treated with 25% of Bitter Melon extract, the cell adhesion were 0.17% and 2.6% respectively compared to control (without Bitter Melon extract were 12.2% and 17% respectively). Truiti Mda et al. [36] reported that ethanol extract of Chaptalia nutans root (7-O-beta-D-glucopyranosylnutanocoumarin) effectively inhibit Staphylococcus aureus and Bacillus subtilis at concentrations of 62.5 g/ml and 125 g/ml, respectively. [36]. Yarrow (Achillea millefolium) is recognized as a powerful medicinal extract showed antibacterial activity against S. Typhimurium and S. aureus with predicted MICs on the order of 10 s of μg/mL or 10 s of mg/mL, respectively [37]. Maldonado et al. [38] reported that E. coli isolated from environmental and food sources where studied for their potential cytotoxicity on African green monkey kidney (Vero, CCL- 81) cells [38]. The food isolates exhibited greater LDH release from the vero cells compared to the environmental isolates. Seventy-one percent (71%) of the food isolates showed high percentage of LDH activity i.e. >50. The outbreak and food isolates exhibited high levels of cytotoxicity. Kang et al. [39] transfected the CHO-K1 with bovine prion protein gene (Prnp) responsible for the bovine spongiform encephalopathy (BSE) and studied cellular changes using LDH assay [39]. Transfected cells showed a relative higher LDH release (6.3%) when compared to wild-type cells (2.7%). The LDH assay suggested that exogenous and endogenous expression of prion protein decreases the cell viability. Kalischuk et al. [40] studied the cytotoxic effect to C. jejuni on human colonic epithelial cell line with a crypt-like phenotype, T84 cells using the LDH enzyme release assay [1]. The C. jejuni treated monolayer showed increased cytotoxicity in 3-hours of inoculation. Gray et al. [41] reported that the toxins of Bacillus cereus and Bacillus thuringiensis using Ped-2E9 cell and CHO cell [41]. They reported that B. cereus and B. thuringiensis exhibiting an alkaline phosphatase activity of >16% were considered cytotoxic strains. Strains with NHE (diarrheal enterotoxin) complex and alkaline phosphatase of >16% were shown to cytotoxic, therefore identified as the pathogenic strains.
Results of this experiment demonstrated that when CHO cells were infected with C. jejuni without Bitter Melon extract, it released high amount of LDH and AP enzymes compared to cells treated with Bitter Melon extract. When CHO cells were infected with C. jejuni ATCC 29428 and FCA 40 (controls) for 16 hour, LDH released was 99% and 91% respectively. Similar trends were observed when CHO cells were infected with various isolates of C. jejuni from poultry samples. When CHO cells were infected with C. jejuni isolates 8 and 17-1 for 16 hour, LDH released was 74% and 70% respectively. When CHO cells were infected with C. jejuni ATCC 29428 and FCA 40 (controls) for 16 hour and treated with 25% Bitter Melon extract, LDH released was significantly lower 61% and 44% respectively compared to controls. These results demonstrated that cytotoxic effects of C. jejuni ATCC 29428 and FCA 40 isolates were inhibited by 25% concentration of Bitter Melon extract, which resulted in reduced cell damage and less LDH released. Same trend was observed, when CHO cells were infected with C. jejuni ATCC 29428 and FCA 40 (controls) for 16 hour and treated with 25% Bitter Melon extract, AP released was significantly lower 41% and 35% respectively compared to CHO cells without Bitter Melon extract (100%).