Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2014) Volume 3, Issue 1
Keywords: Trichoderma, Cellulase, Xylanase, Wheat bran, Cellulose powder
Research on micro-organisms that utilize xylan, Cellulases and on the involvement of enzyme system, is important from ecological and economic point of view. In a time of rising fuel prices, utilization of abundantly available biomass as an alternative resource of energy is one of the thrust area of research. Currently researchers throughout the world focus on the utilization of renewable, economical and abundantly available biomass for the production of useful products. Cellulose is the most common organic polymer, representing about 1.5 ×1012 tons of the total annual biomass production through photosynthesis especially in the tropics, and is considered to be an almost inexhaustible source of raw material for different products [1]. It is the most abundant, cheapest and renewable biopolymer on earth and the dominating waste material from agriculture [2]. A promising strategy for efficient utilization of this renewable resource is the microbial hydrolysis of lignocellulosic waste and fermentation of the resultant reducing sugars for production of desired metabolites or biofuels [3]. Cellulose is generally degraded by a multi-complex enzyme called cellulases. Synergistic mechanism among three types of enzymes namely endoglucanase, exoglucanase and β-glucosidase are required for the complete enzymatic hydrolysis of cellulose biopolymer [4].
Cellulases are produced by a number of microorganisms such as bacteria, yeast and fungi [5]. However, the most extensively studied cellulases are those produced by efficient lignocellulose degrading fungi, particularly Trichoderma [6]. The cellulase produced by Trichoderma is resistant to chemical inhibitors and stable in stirred tank reactors at pH 4.8, 50°C for 48h or longer [7]. Trichoderma has the capacity to secrete enzymes in high yields and this property can be exploited when using Trichoderma as an industrial host for homologous and heterologous enzyme production [8]. Besides, enzymes produced from Trichoderma possess high activities of both endoglucanase and exoglucanase but low β-glucosidase activity [9]. Trichoderma harzianum and Trichoderma azad BGLs is subjected to produce inhibition and though it is sufficient to support growth on cellulosic material; it is often supplemented with Aspergillus BGLs for complete biomass saccharification at industrial level [10]. Nevertheless, it is the best cellulase available today.
The major bottle neck against comprehensive application of cellulases in biofuel industry is the high cost of the enzyme production. Therefore, extensive research is needed to enhance the enzyme activity so that less enzyme is needed for the complete hydrolysis of biomass. Cellulose quality, temperature, carbon sources, incubation period and pH of the medium are important parameters for the optimized production of cellulase enzymes [11]. Considering the importance and application of cellulases, present study was aimed to isolate and screen the fungal isolates for the cellulolytic ability. Moreover, in the present study three different substrates such as wheat bran, birch wood xylan and cellulose powder were used and the effect of various parameters like pH and temperature were also studied on cellulase production.
Chemical reagents
All chemicals and reagents used were of analytical grade. Different carbon sources such as birch wood xylan, Salicin, Carboxy-methyl cellulose (CMC) and glucose were obtained from Hi-media, Mumbai India; whereas wheat bran was purchased from local market of Kanpur, Uttar Pradesh, India.
Isolation of fungal culture
To isolate cellulase producing microorganisms, soil sample from decomposing sites of Biocontrol Laboratory of C.S. Azad University of Agriculture and Technology, Kanpur was taken. The decomposing residue brought in the laboratory was kept at 4°C ± 1. The cellulosehydrolytic fungus was isolated by direct agar plate method of Warcup [12] using Czapek Dox Agar medium. The medium consists of: 3 g/L NaNO3, 1 g/L K2HPO4, 0.5 g/L MgSO4.7H2O, 0.5 g/L KCl, 0.01 g/L FeSO4.7H2O, 30 g/L sucrose, 15 g/L Agar powder. Streptomycin was also added to the molten medium after autoclave. The pH of the medium was kept at 5.5. For isolation, 0.5g of soil sample was transferred with the help of sterile flattered tip needle and was dispersed in a drop of sterile water in the bottom of petridish. The sterilized Czapek Dox agar medium was then poured into the petridish. The plated sample was dispersed in the plate by rotating the medium before its solidification. After this plates were kept in incubator at 28°C ± 1 for their growth.
Purification of fungal isolate
The contaminated cultures of fungal isolates were purified and derived through single spores, using dilution plating [13]. To ensure purity, cultures were derived from single spore in each case. The bacterial contamination was prevented by using antibiotic streptomycin (50mg/100 ml) in the medium during isolation and purification of fungi. After ensuring purity, the cultures were maintained in PDA slants at 4°C.
Screening for cellulolytic activity
Screening for cellulolytic activity was made using PDA media supplemented with 5% carboxy-methyl cellulose (CMC). After certain incubation the plates were stained by Congo red to see the cellulolytic activity of isolated strains. The cellulase activity of each culture was determined by measuring the zone of clearance on agar plate [14].
Preparation of enzyme source and enzyme assay
Isolated fungal strain was grown in Mandel and Reese medium [7] supplemented with different carbon sources (1% of xylan, wheat bran or cellulose powder) at 28°C for 7 days. After cell growth reached early stationary phase, the culture broth was centrifuged at 9500 rpm for 10 minutes (4°C) and the supernatant was used as extracellular source of enzyme. Cellulase activity was determined according to the methods described by the Mandels [15]. For endoglucanases the reaction mixture consists of 0.5 ml. of 1% carboxy methyl cellulose (CMC) in citrate buffer (pH 4.8), 0.5 ml of enzyme sample, diluted with citrate buffer (pH 4.8). The reaction mixture is incubated at 50°C for 30 minutes. 3ml of DNS reagent was added and the tubes were placed in boiling water bath for 5min. The tubes were cooled and the reducing sugar (glucose) was determined. Similarly, the exoglucanase activity was determined by using 6 × 1 cm Whatmann no. 1 filter paper strips, cut into small strips. The strips were incubated with 1 mL of 0.1 M citrate buffer (pH 4.8) containing 0.5 mL culture filtrate at 50°C and the reducing sugars released was determined after 60 minutes intervals by DNS method [16].
For β-glucosidase activity, the reaction mixture, consisting of 0.5 mL of 1% salicin solution, 0.5 mL of 0.1 M citrate buffer (pH 4.8) and 0.5 mL of enzyme sample, was incubated at 50°C for 30 minutes. The tubes were cooled and the reducing sugar (glucose) was determined. This was determined by measuring the absorbance at 540nm using Lambda 25 UV/Vis Spectrophotometer. In accord with the International Union of Biochemistry, one enzyme unit (U) equals to 1μmol of glucose released per minute.
Xylanase activity
Trichoderma species was maintained at 4°C after growing for 7 days in a culture medium containing 0.2% malt extract, 0.2% yeast extract, 2% glucose and 2% agar (MYG medium) at 28°C. Sporulation medium was prepared by using Trisodium citrate (0.5g), KH2PO4(0.5g), NH4NO3 (0.2g), (NH4)2 SO4 (0.4g), MgSO4 (0.02g), peptone (0.1g), yeast extract (0.2g), glucose (0.2g), and agar – agar (2.5g) and the volume was made upto 100ml with double distilled water at 5.5 pH and 28°C temperature. Inoculation medium (100ml) was prepared in the same way but without agar –agar. A loopful culture of Trichoderma species from sporulation medium was transferred aseptically into the Erlenmeyer flask, containing 100ml Vogel’s medium prepared as per standard method. The flasks were incubated on rotary shaker at 150 rpm at 28°C for 5 days. Liquid state cultures were harvested by centrifugation at 10,000 rpm for 20 min. at 4°C, and the resulting supernatant was used as crude enzyme preparation. Xylanase activity of this crude enzyme was measured as per method described by Khanna and Gauri [17] using Xylose as standard. The unit (1U) of enzyme activity was defined as the amount of enzyme required to liberate 1 m mole min-1 of xylose from the appropriate substrate under the standard conditions. Trichoderma species was grown in Vogel’s medium with different carbon sources like glucose, maltose, CMC, sucrose, corn-cobs, wheat bran, oat splet xylan and birch wood xylan to check their effect on the expression of xylanase. The crude xylanase activity was measured in pH range of 4.0 to 8.0 with different buffers. Sodium acetate buffer was used to maintain pH of 4-5.5 and citrate phosphate buffer was used to maintain pH of 6-8. The reactions were carried out at 40, 50, 60 and 70°C for determination of optimum temperature and xylanase assay was carried out.
Characterization
Effect of pH, temperature and kinetics of enzyme activity: Effect of pH on enzyme activity was determined by incubating enzyme in buffers of different pH during standard enzyme assays. The buffers used were 0.1M sodium citrate (pH 4 and pH 4.8) and 0.2M phosphate buffer (pH 5.0 and 5.5). For estimation of the optimum temperature of the enzyme, the activity was determined by carrying out the standard assay at several temperatures between 28°C and 32°C at pH5.5. Kinetics of enzyme production was studied for culture of T. harzianum Thazad by growing them in synthetic media supplemented with cellulose powder; incubating at 28° C and withdrawing samples of cell-free culture supernatant at 0,1,2,3,4 and 5 days intervals. Enzyme activity was measured by cellulase assay.
Effect of various carbon sources on cellulases production: Different strains of Trichoderma species were grown in Mandel’s and Reese medium with different carbon sources viz., cellulose powder, birch wood xylan and wheat bran to check their effect on the expression of cellulases. All samples were analyzed in duplicates and the mean values calculated.
Isolation and identification of fungal strain
A total of two isolates of fungi were isolated from rhizospheric soils of legume crop and identified by Indian Type Culture Collection (ITCC), IARI, New Delhi. Identification of fungal strain was made on the basis of morphological and microscopic characteristics by using standard reference manuals [18,19] (Figure 1 & Figure 2).
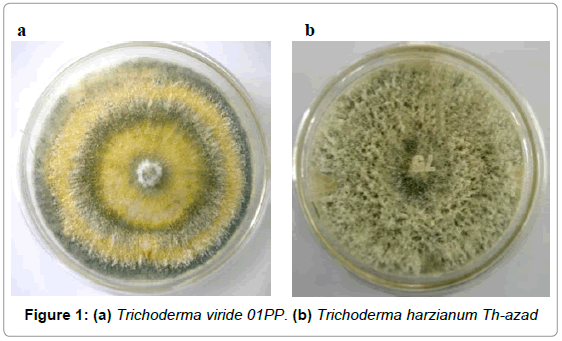
Figure 1: (a) Trichoderma viride 01PP. (b) Trichoderma harzianum Th-azad

Figure 2: Conidia and Conidiophore branching in Trichoderma viride 01PP and T.harzianum Th-azad using lactophenol cotton blue (100X).
Screening of fungal isolate for cellulolytic activity
Screening of fungus was conducted by using Congo red test as a preliminary study for identifying cellulase producer. The fungal isolates which showed a maximum clear zone on the cellulose and xylose containing medium after staining with Congo red was taken. The diameter of hydrolytic zones was measured which was found to be 10mm and 19mm. (Figure 3a & 3b).
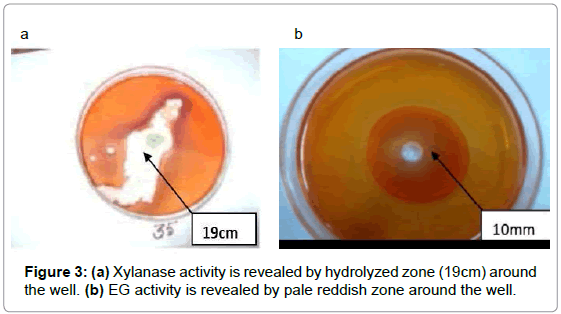
Figure 3: (a) Xylanase activity is revealed by hydrolyzed zone (19cm) around the well. (b) EG activity is revealed by pale reddish zone around the well.
Cellulase assay
CMCase, FPase and BGL assays were carried out respectively for measurement of endoglucanase, exoglucanase and β-glucosidase activity (Table 1). The result shows that culture of T.harzianum Thazad shows higher cellulase activity (EG-5.89 IU/ml, EXG-4.32 IU/ml and BGL-6.08 IU/ml; pH 5.5) than the T. viride 01PP with wheat bran as sole carbon source. The results reveal that T. harzianum Th-azad shows highest endoglucanase and exoglucanase activity than T. viride 01PP. Earlier it has been reported that endoglucanase was induced by CMC but repressed by glucose [20]. It was also found that Wheat bran, carboxy-methyl cellulose and cellulose powder was preferred substrate for EG and EXG production [21].
| S.No. | Species | Substrate | pH | EG activity (IU/ml) | EXG activity (IU/ml) | BGL activity (IU/ml) | Xylanase activity (IU/ml) |
|---|---|---|---|---|---|---|---|
| 1. | T.harzianum Th-azad | Cellulose powder | 4.0 | 3.11 | 0.46 | 3.2 | 12.04 |
| 5.0 | 3.816 | 1.315 | 4.07 | 1.146 | |||
| 5.5 | 4.692 | 2.759 | 6.01 | 14.73 | |||
| Wheat bran | 4.0 | 2.173 | 0.622 | 1.84 | 10.73 | ||
| 5.0 | 5.382 | 3.274 | 4.89 | 1.766 | |||
| 5.5 | 5.89 | 4.32 | 6.08 | 15.96 | |||
| 2. | T.viride 01PP | Cellulose powder | 4.0 | 2.12 | 0.39 | 2.18 | 9.89 |
| 5.0 | 2.89 | 1.03 | 3.11 | 1.104 | |||
| 5.5 | 3.26 | 1.52 | 4.05 | 10.02 | |||
| Wheat bran | 4.0 | 2.10 | 0.46 | 1.82 | 8.46 | ||
| 5.0 | 4.33 | 2.11 | 4.22 | 1.56 | |||
| 5.5 | 3.67 | 3.16 | 5.06 | 12.06 |
Table 1: Comparative analysis between T. harzianum Th-azad and T. viride 01PP for Cellulases (EG, EXG and BGL) and Xylanase production on different carbon sources at various pH.
Xylanase assay
Xylanase activity was measured according to the method of Bailey et al. [22] using 1 ml of 1% (w/v) solution of birch wood xylan. It was found that the culture of T.harzianum Th-azad showed highest xylanase activity (15.96 IU/ml) as compared to other isolate with wheat bran as a substrate. Wheat bran is an inexpensive by product, which contains a lot of xylan. Therefore, it is one of the most popular components of complex media for xylanase production [23-26]. However, the wheat bran particles suspended in the cultivation medium have to be decomposed to form soluble compounds to be used by the fungus and also protects the fungal mycelium from the shear forces. Several substances have been indicated in the literature as suitable carbon sources for xylanase-producing micro-organisms, oat wheat [27], birch wood xylan [28], oat spelt xylan [29], bagasse xylan, wheat bran arabinoxylan [30], wheat bran, sugar cane bagasse and rice bran [31] are few recommended sources.
Characterization
Effect of pH on enzyme production: Cellulase yield by different strains of Trichoderma species were found to depend on pH value. The effect of pH on the crude cellulase and xylanase enzyme production by fungal isolates was examined at different pH values ranging from 4.0 to 5.5. Results illustrated by Table-1, clearly show that cellulase and xylanase production, expressed as enzyme activity, gradually increased as the pH value increased from 4-5.5 and reached its optimum level of pH 5.5 as also reported. Further increase in pH declines the enzyme activity.
Effect of Incubation temperature on enzyme production: Effect of incubation temperature on cellulolytic enzyme production was studied by growing fungal isolates at different temperatures. The optimum temperature for maximal EXG, EG, BGL and xylanase production was found to be 28°C (Figure 4). Further increase in temperature resulted in decrease in enzyme production. The temperature of the fermentation medium is one of critical factor that has profound influence on the production of end product. The optimum temperature of 28°C for enzyme production by Trichoderma species is in accordance with earlier research. Zhou et al. [32] grew Trichoderma viride 01PP at 28°C for optimal cellulases production. β-glucosidase was produced and purified from Trichoderma harzianum type C-4 grown at 28° C [33]. Exo β-1, 3 glucanase was produced from Trichoderma asperullum at 280 C [34]. Similarly cellulases were produced from Aspergillus niger KK2 at 28°C [35]. Similarly, maximum cellulase production in Aspergillus terreus was obtained at a temperature of 28° C [36].
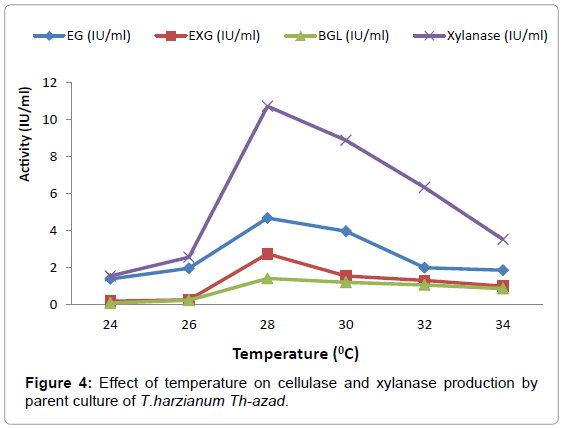
Figure 4: Effect of temperature on cellulase and xylanase production by parent culture of T.harzianum Th-azad.
Effect of incubation period on enzyme production: Time course for cellulases production of T.harzianum Th-azad was also investigated. Optimum EXG, EG and BGL production was achieved at 120 h of incubation at pH 5 and 28° C (Figure 5). Further increase in incubation time resulted in decrease in EXG, EG and BGL activities. It may be due to the depletion of nutrients in the medium which stressed the fungal physiology resulting in the inactivation of secretary machinery of the enzymes [37]. Time course of 120 h for optimum cellulases production by T. harzianum Th-azad is in accordance with earlier reports. Cellulase were produced from Aspergillus niger KK2 at 120 h incubation [35]. Similarly cellulolytic enzymes were produced by Aspergillus phoenix at 120 h incubation [38].
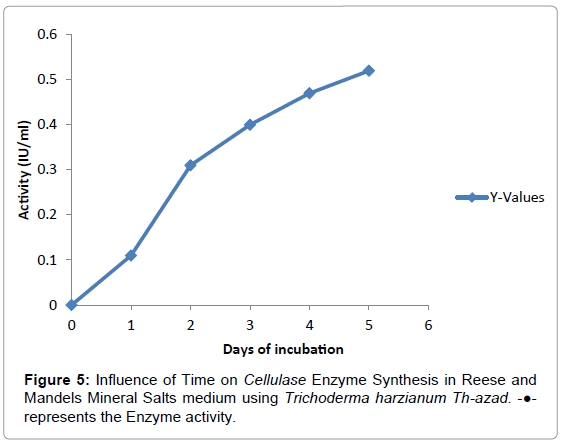
Figure 5: Influence of Time on Cellulase Enzyme Synthesis in Reese and Mandels Mineral Salts medium using Trichoderma harzianum Th-azad. -●- represents the Enzyme activity.
Effect of carbon sources on enzyme production: Data presented in table 1 shows that cellulase and xylanase production by different strains of Trichoderma species was significantly influenced by the type of carbon source in the basal salt medium. In this study two different carbon sources viz., wheat bran, birch wood xylan and cellulose powder were investigated. The result shows that wheat bran is the most effective sole carbon source for cellulase production followed by cellulose powder. The results also revealed that culture of T.harzianum Th-azad shows highest cellulase production than other species. The study also reveals that among the culture of T.harzianum Th-azad shows higher xylanase activity (15.96 IU/ml). Mandel and Reese [7] also reported that maximum yields of cellulase were obtained on 1% cellulose powder carbon substrate using T.viride 01PP. The substrate not only serves as a carbon source but also produces the necessary inducing compounds for the organism [39]. Reduction in the cost of cellulase production can be achieved by the use of cheap and easily available substrates. Earlier it has been reported that endoglucanase was induced by CMC but repressed by glucose [20]. It was also found that wheat bran was preferred substrate for EG production. Niranjane et al. [40] observed highest yields of cellulases on CMC. In this study, we recorded the similar results with very less cellulases activities in the presence of glucose, while wheat bran proved to be the best and cheap substrate for cellulase and xylanase production. Shahid et. al. [41] observed that among various substances used for the maximum xylanase production, birch wood xylan at a concentration of 1% proved to the best carbon source (substrate) for Trichoderma viride as it observed highest xylanase activity at a concentration of 1%.
Trichoderma species did not show xylanolytic activity without the presence of any carbon source.
The fungi as enzyme sources have many advantages that, the enzymes produced are normally extracellular, making easier for downstream process. To identify the cellulase enzymes and to study the activity of the cellulase complexes of Trichoderma species a series of experiments were conducted. In this study comparative analysis and substrate optimization was conducted among various cultures of Trichoderma species isolated from decomposing soil sample. The results clearly show that culture of T. harzianum Th-azad exhibited better activity. The optimum pH of cellulase and xylanase enzyme was found to be pH 5.5 and the optimum reaction temperature and incubation period to be at 28°C and 120 h. During our investigation we also found that the fungal isolates shows better cellulase and xylanase activity with wheat bran as sole carbon source. Finally, it should be noted that wheat bran is a cheaper raw material, since it is the by-product of agricultural wheat production and readily available in large amounts. Therefore, it is concluded that wheat bran can be utilized as a cheap substrate in various industrial sectors for production of various useful products such as biofuels.
The authors are grateful for the financial support granted by the ICAR under the Niche Area of Excellence on “Exploration and Exploitation of Trichoderma as an antagonist against soil borne pathogens” running in the Biocontrol Laboratory, Department of Plant Pathology, C.S.A University of Agriculture and Technology, Kanpur, India.