Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2013) Volume 2, Issue 2
Keywords: Streptomyces albolongus, Streptomyces aburaviences, Extracellular proteases, Culture conditions
Proteases play a colossal role in biotechnology and are widely used in tanning industry, in the manufacturing of biological detergents, meat tenderization, peptide synthesis, food industry, pharmaceutical industry, and in bioremediation processes [1-3]. Although protease can be obtained from several sources, including plant, animal and microorganisms, microbial proteases are preferred in view of their rapid growth, ease of cultivation purification and genetic manipulation. A large number of microorganisms have been reported for protease production [4-8].
Microbial proteases can be produced using many processes like solid-state fermentation and submerged fermentation [9,10]. Cultural conditions (physical, chemical and nutritional factors) play significant role in the production of extracellular proteases by microorganisms [11-13]. Physical factors include aeration, temperature, pH and incubation time. In addition to these physical factors, nutritional factors such as the sources of carbon and nitrogen also significantly affect protease production.
Researchers have examined bacteria and fungi from various habitants to obtain suitable proteases. At present, majority of commercially available proteases are secreted by Bacillus spp., although there have been increasing reports of the potential use of proteases of fungal origin [14-16]. Actinomycetes as a source of naturally occurring extracellular protease is overlooked and information on proteases from actinomycetes has been limited. However, the industrial demands of proteolytic enzymes stimulate the search of new enzyme sources with extended range of applications. Among the actinomycetes, several species of the Streptomyces are among the most important industrial microorganisms because of their capacity to produce numerous bioactive molecules, particularly antibiotics. Streptomyces species are heterotrophic feeders, which can utilize both complex and simple molecules as nutrients. In addition to antibiotics, Streptomyces species liberate several extra cellular enzymes [17]. They produce variety of extra cellular proteases that have been related to aerial mycelium formation and sporulation [18]. In view of the above, the present work was under taken to study the effects of culture conditions on the production of extracellular proteases by S. albolongus and S. aburaviences, and also aimed at optimization of media composition, which has been predicted to play a significant role in enhancing the production of protease.
Microorganism
Two isolates of actinomycetes, S. albolongus (A5) and S. aburaviensis (RB20) were collected from the laboratory stock culture of the Department of Microbiology, University of Chittagong. For the growth and preservation of the isolates, nutrient agar was used.
Screening of the isolate for proteolytic activity
The organisms’ ability to produce extracellular protease enzyme were determined by growing the isolates in solid medium containing protein sources like gelatin, skimmed milk casein and boiled egg albumin. Secondary screening of the isolates was done by measuring the protease activity in liquid medium by quantitative method. For this purpose, the isolates were inoculated into three liquid medium, viz. (i) Peptone yeast extract-dextrose broth (Yeast Extract 1%, Peptone 2%, Dextrose 2%) [19] (ii) Tryptone-yeast extract-dextrose broth (Tryptone 1%, Dextrose 0.1%, Yeast extract 0.5%) [20] (iii) Gelatin-yeast extract- glucose broth (Gelatin 1%, glucose 1%, Yeast Extract 0.2%, K2HPO4 0.3%, KH2PO4 0.1%, MgSO4.7H2O trace) [21] at pH 7.0 and were incubated at 35 ± 2°C for 5 days [22].
Measurement of enzyme activity
After incubation, the broth cultures were filtered with Whatman grade 1 filter paper. Then the filtrates were centrifuged at 8,000 rpm for 15 minutes at 4°C. The supernatant was used as crude enzyme. The enzymes were stored at 4°C with few drops of toluene/Sodium Azide to avoid bacterial contamination. Protease assay was done by using the method described by Meyers and Ahearn [23], which is a modified method of Hayashi et al. [24]. Briefly, 3 ml of crude enzyme, 3 ml of citrate phosphate buffer and 3 ml of 1% (w/v) casein was taken in a 25 ml test tube, and the tube was placed in a water bath at 35°C for 1 hour. Enzyme-substrate reaction was stopped by adding 5 ml of 20% (w/v) trichloroacetic acid (TCA). After one hour, the solution was filtered by Whatman grade 540 (ash less) filter paper. From the filtrate, 1 ml enzyme-substrate mixture was taken into a test tube and 2 ml of 20% Na2CO3 was added to it. To this mixture, 1 ml of Folin-Ciocalteu reagent was added and the contents of the tube were mixed well immediately. After 30 minutes, 6 ml distilled water was added to the tube and absorbance of the solution was measured at 650 nm in a Vis-UV spectrophotometer (LaboMedInp). The amounts of amino acids released were determined using a standard curve plotted from known concentration of tyrosine. The enzyme activity was expressed in Unit. One unit of enzyme was defined as the amount of enzyme that releases 1 g of tyrosine from substrate (casein) per hour, under assay conditions.
Biomass yield
The actinomycetes cultures were filtered through Whatman grade 1 filter paper. The filter paper was dried in oven at 80°C to a constant weight. The amount of biomass was calculated by subtracting the weight of filter paper. The yield was expressed as mg/g of protein.
Optimization of culture conditions
Broth cultures were carried out at various culture conditions such as temperature (10, 27, 37, 45°C), initial pH of the culture media (4, 5, 6, 7, 8, 9), incubation time (1, 2, 3, 4, 5, 6, 7 days), to optimize the culture conditions for the production of extracellular proteases. To determine the effects of aeration, inoculated mediums were incubated at both stationary and shaking condition (100 rpm), keeping other experimental conditions at optimum.
Effects of carbon and nitrogen sources
The production of extracellular proteases under different carbon and nitrogen availability was studied in the liquid culture medium. Four carbon sources (glucose, lactose, fructose, galactose), five organic and two inorganic nitrogen sources (Gelatin, Yeast Extract, peptone, Beef Extract, Pulse, KNO3, (NH4)2HPO4,) were added to the medium, and the effect of this carbon and nitrogen sources on the production of protease was recorded. To ascertain optimum percent of carbon and nitrogen sources, the study was carried out with 0.5 to 2.5% carbon and 0.5 to 2.5% nitrogen sources, keeping other experimental conditions at optimum.
Two isolates of actinomycetes, Streptomyces albolongus and Streptomyces aburaviences were tested for proteolytic activity. Primary screening was done by the boiled egg albumin degradation, gelatin hydrolysis and skimmed milk casein hydrolysis method. Both the isolates showed clear zone of hydrolysis in gelatin agar plate and casein agar plate after 3 days of incubation at 37°C (Figures 1A and 1B). Complete degradation of egg albumin was observed after 7 and 9 days of incubation at 37°C for S. albolongus and S. aburaviensis, respectively (Figure 1C). The isolates were allowed to grow in three liquid media and maximum enzyme activity were found in Gelatin- yeast extract- glucose broth and tryptone-dextrose-yeast extract broth for S. albolongus and S. aburaviensis, respectively (Table 1).

Figure 1: Photograph showing clear zone of hydrolysis A. casein hydrolysis by S. albolongus (A-1) and S. aburaviensis (A-2), B. Gelatin hydrolysis by S. albolongus (B-1) and S. aburaviensis (B-2) and C. Egg albumin degradation.
| Culture medium | Protease activity (U/ml) Isolates of actinomycetes | |
|---|---|---|
| S. albolongus | S. aburaviensis | |
| Yeast Extract 1%, Peptone 2%, Dextrose 2% | 173.07 | 160.02 |
| Tryptone 1%, Dextrose 0.1% Yeast extract 0.5% | 326.92 | 474.58 |
| Gelatin 1%, glucose 1%, Yeast Extract 0.2%, K2HPO4 0.3%, KH2PO4 0.1% MgSO4 trace | 631.86 | 253.43 |
Enzyme-substrate reaction pH and temperature was 6.5 and 35°C, respectively.
Table 1: Protease activities of the isolates in different liquid medium.
Effects of incubation period on the production of proteases
S. albolongus showed maximum enzyme production (789.14 U/ml) after 5 days of incubation and highest protease production (567.99 U/ml) by S. aburaviensis was recorded after 4 days of incubation (Figure 2). For both isolates, highest biomass yield (140 mg/g substrate for S. albolongus and 150 mg/g substrate for S. aburaviensis) were observed after 6 days of incubation, Table 2 associated with surface and sedimentary growth with white sporulation in case of S. albolongus and grayish sporulation in case of S. aburaviensis. The pH of the culture filtrates was ranged from 5.26 to 6.14 and 6.10 to 6.90 for S. albolongus and S. aburaviensis, respectively.
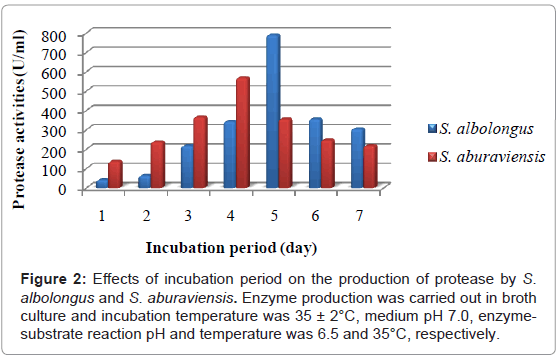
Figure 2: Effects of incubation period on the production of protease by S. albolongus and S. aburaviensis. Enzyme production was carried out in broth culture and incubation temperature was 35 ± 2°C, medium pH 7.0, enzymesubstrate reaction pH and temperature was 6.5 and 35°C, respectively.
| Incubation Period (in days) | Biomass yield (mg/gm of substrate) | |
|---|---|---|
| S. albolongus | S. aburaviensis | |
| 1 | 4 | 12 |
| 2 | 40 | 30 |
| 3 | 60 | 36 |
| 4 | 80 | 102 |
| 5 | 136 | 134 |
| 6 | 140 | 150 |
| 7 | 132 | 121 |
Table 2: Effects of incubation period on biomass yield.
Effects of medium pH on the production of protease
Figure 3 represents the protease activities of the isolate S. albolongus and S. aburaviensis at different initial pH of culture medium. S. albolongus showed highest enzyme production (795.32 U/ml), having medium pH 7.0, but highest biomass yield (160 mg/g substrate) was recorded with medium pH 8.0 (Table 3). The pH of the culture filtrate ranged from 5.12 to 7.88. The isolate exhibited surface and sedimentary growth with white sporulation at medium pH 7.0. On the other hand, S. aburaviensis showed highest enzyme production (612.63 U/ml) and maximum biomass yield (180 mg/g substrate) at pH 8.0. The pH of the culture filtrates ranged from 5.19 to 7.93. Biomass characteristics of the isolate were varied with the pH of the medium.
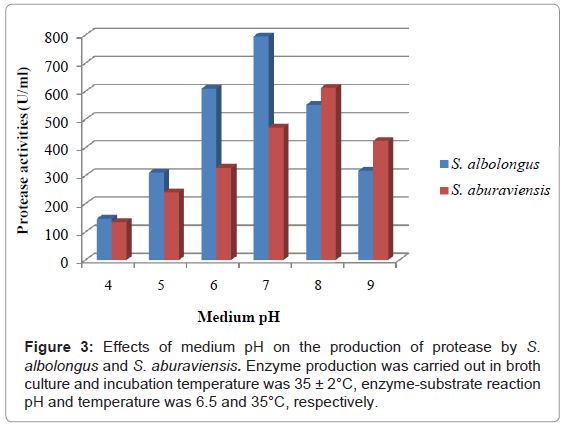
Figure 3: Effects of medium pH on the production of protease by S. albolongus and S. aburaviensis. Enzyme production was carried out in broth culture and incubation temperature was 35 ± 2°C, enzyme-substrate reaction pH and temperature was 6.5 and 35°C, respectively.
| Medium pH | Biomass yield (mg/gm of substrate) | |
|---|---|---|
| S. albolongus | S. aburaviensis | |
| 4 | 44 | 9 |
| 5 | 80 | 29 |
| 6 | 122 | 115 |
| 7 | 141 | 155 |
| 8 | 160 | 180 |
| 9 | 106 | 127 |
Table 3: Effects of medium pH on biomass yield.
Effects of incubation temperature on production of proteases
Both S. albolongus and S. aburaviensis showed maximum protease production (747.24 U/ml and 619.50 U/ml respectively) (Figure 4) and maximum biomass yield (152 mg/g substrate and 174 mg/g substrate respectively), at 37°C associated with surface and sedimentary growth with sporulation. Table 4 represents biomass yields of the isolates at different incubation temperature. The pH of the culture filtrates was ranged from 7.10 to 7.89 for S. albolongus and 7.52 to 7.79 for S. aburaviensis.
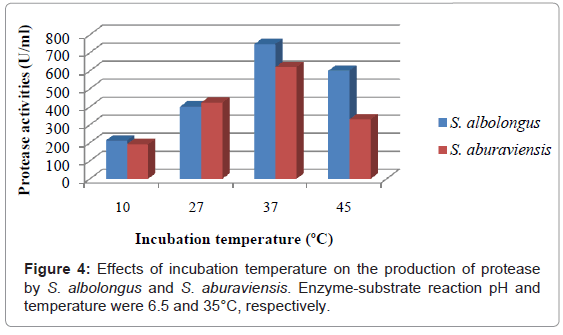
Figure 4: Effects of incubation temperature on the production of protease by S. albolongus and S. aburaviensis. Enzyme-substrate reaction pH and temperature were 6.5 and 35°C, respectively.
| Temperature (0°C) | Biomass yield (mg/gm of substrate) | |
|---|---|---|
| S. albolongus | S. aburaviensis | |
| 10 | 38 | 34 |
| 27 | 72 | 130 |
| 37 | 152 | 174 |
| 45 | 114 | 118 |
Table 4: Effect of temperatures on biomass yield.
Effects of stationary and shaking conditions on the production of proteases
Stationary and shaking conditions have marked influence on protease production. S. albolongus showed maximum protease activity (683.60 U/ml) at stationary condition, whereas shaking at 100 rpm significantly increased protease production (847.28 U/lm) by S. aburaviensis (Figure 5).
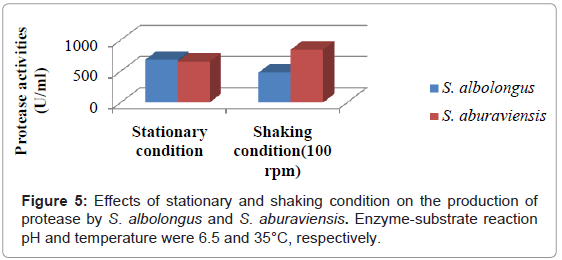
Figure 5: Effects of stationary and shaking condition on the production of protease by S. albolongus and S. aburaviensis. Enzyme-substrate reaction pH and temperature were 6.5 and 35°C, respectively.
Effects of carbon and nitrogen sources on the production of proteases
To investigate the effects of various carbon and nitrogen sources, the isolates were allowed to grow in different media containing four different carbon sources and five organic and two inorganic nitrogen sources. S. albolongus exhibited highest enzyme activity (673.76 U/ml) in glucose and beef extract containing media (Figure 6). Maximum protease (734.95 U/ml) was released when 1% glucose, and 2% beef extract were used as a carbon and nitrogen source in the growth medium (Figure 7). The strain S. aburaviensis showed maximum enzyme activity (484.62 U/ml) in lactose and tryptone containing medium (Figure 8). The isolate S. buraviensis showed maximum protease activity (571.27 U/ml), when 1.5% lactose and 1.5% tryptone were used as carbon and nitrogen source, respectively (Figure 9).
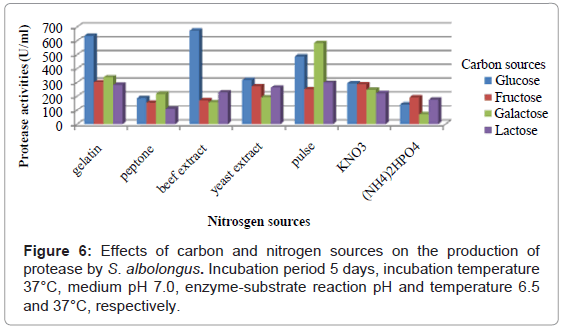
Figure 6: Effects of carbon and nitrogen sources on the production of protease by S. albolongus. Incubation period 5 days, incubation temperature 37°C, medium pH 7.0, enzyme-substrate reaction pH and temperature 6.5 and 37°C, respectively.
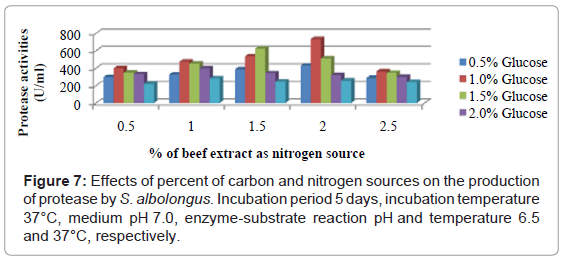
Figure 7: Effects of percent of carbon and nitrogen sources on the production of protease by S. albolongus. Incubation period 5 days, incubation temperature 37°C, medium pH 7.0, enzyme-substrate reaction pH and temperature 6.5 and 37°C, respectively.
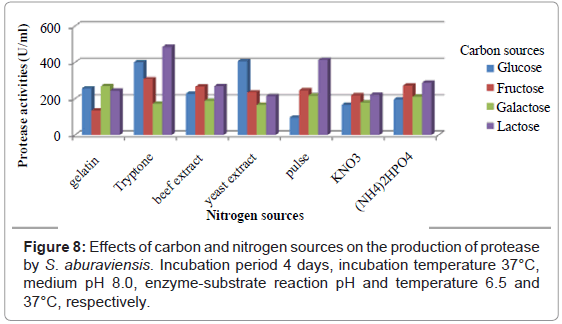
Figure 8: Effects of carbon and nitrogen sources on the production of protease by S. aburaviensis. Incubation period 4 days, incubation temperature 37°C, medium pH 8.0, enzyme-substrate reaction pH and temperature 6.5 and 37°C, respectively.

Figure 9: Effects of percent of carbon and nitrogen sources on the production of protease by S. aburaviensis. Incubation period 4 days, incubation temperature 37°C, medium pH 8.0, enzyme-substrate reaction pH and temperature 6.5 and 37°C, respectively.
The results of primary screening indicated that both S. albolongus and S. aburaviensis have the ability to produce extracellular protease in solid medium under static incubation condition. Then the isolates were allowed to grow in three liquid media and maximum enzyme activity were found in Gelatin-yeast extract-glucose broth and tryptone-dextrose-yeast extract broth for S. albolongus and S. aburaviensis, respectively. There was a gradual increase of protease production up to day 4 for both the isolates. Maximum production was obtained after 5 days of incubation for S. albolongus, and it was 4 days for S. aburaviensis, but maximum biomass was obtained after 6 days for both isolates. Maximum production of extra cellular protease after 120 hours of incubation by Streptomyces sp. 594, which was not growth associated, was reported by Azeredo et al. [25]. Our results were in concurrence with them.
Microorganisms are very sensitive to the concentration of hydrogen ions present in the medium. So, pH is one of the most important factors that determine the growth and the production of protease by microbes. S. albolongus preferred medium with neutral pH for maximum enzyme production, but maximum protease activities S. aburaviensis was obtained in medium having initial pH 8.0. Production of microbial proteases in neutral and alkaline medium pH was also reported by other authors [25-27]. Temperature is an important environmental factor for growth and enzyme production. The best incubation temperature for both the isolates was 37°C. A wide range of temperature (30°C-55°C) has been reported for optimum growth and protease production by Streptomyces sp. [13,25]. Extracellular protease from S. albolongus was optimally produced when incubated at static condition. About nine fold greater protease activity from Streptomyces sp. in stationary culture than produced under shake flask condition was reported by Gibb et al. [28]. Our finding with S. albolongus was in accordance with them. Shaking at 100 rpm markedly increase proteases production by S. aburaviensis. This finding was also reflected in other studies [24,26,28,29].
Microorganisms show a considerable variation in their nutrient requirements. Carbon and nitrogen sources are important variables that affect the growth and products of microbes [30]. Many authors had reported variability of carbon and nitrogen sources with different microorganisms [31-33]. Shafee et al. [30] had reported that maximum protease was produced in medium containing glucose and beef extract, which was similar to our findings with S. albolongus. In case of S. aburaviensis, highest protease production was observed in medium containing lactose and tryptone. Both the isolates produced proteases in response to both organic and inorganic forms of nitrogen, but preferred organic nitrogen for better production. Similar result was reflected in other studies [22,31]. Concentration of carbon and nitrogen sources in the production media significantly affects protease [34-36]. Variability in the production of protease with different percent of carbon and nitrogen sources was also observed in the present investigation. Glucose concentration higher than 1.0 % was found to reduce protease yield from S. albolongus, when 2.0% beef extract was used as nitrogen source. In case of S. aburaviensis protease production was gradually increased with the increase of lactose concentration, and 1.5% was found optimum when 1.5% tryptone was used as nitrogen source.
Streptomyces albolongus and Strepmyces aburaviensis are potential microbes for extracellular protease production, and various environmental and nutritional factors have significant effects on their growth and protease production. In the present investigation, we have determined the optimum conditions for maximum production of extracellular proteases. The optimized media composition and cultural conditions might be implemented in large scale for the production of extracellular proteases by Streptomyces albolongus and Strepmyces aburaviensis.