Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2010) Volume 1, Issue 1
Bacterial infection is a major cause of high mortality in juvenile sea turtles at the Sea Turtle Conservation Centre, Chonburi Province, Thailand. The attempt is to identify sources of bacterial accumulation and contamination in order to reduce the number of infections. The aim of this study was to determine the effects of live rocks and captivity parameters on microbiological water quality. Microbiological results indicated that the presence of live rocks in juvenile green turtle containers resulted in a reduction of haemolytic bacteria in seawater. Additionally, the high stock density of animals in the captivity has influence on the increased bacterial levels in the water, while temperature of the water was found to have an influence on bacterial growth. The results of this study implied that the environments of captivity have great impact on the bacterial levels in the water, which should be considered for the management of sea turtles as well as other aquatic animals
Keywords: Seawater quality, sea turtles, bacterial assessment, live rocks, captivity environments.
Green sea turtle (Chelonia mydas) is one of seven sea turtle species that have been listed as endangered species in Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) (http://www.cites.org/eng/app/Appendices-E.pdf, p. 25). Increasing the number of animals in nature is one major objective of conservation programmes in several countries. In maintenance of sea turtles for research and conservation, public display, or other purposes, several factors need to be taken into account: rearing facilities, seawater system, and environmental parameters [1]. Presently, knowledge about the basic biology of sea turtles is limited. As a result, sea turtle husbandry in many facilities has been carried out on a trial and error basis. Therefore, information about the effects of environmental parameters on sea turtles is urgently needed for the effective management of the animals in captivity.
Water quality is critical for aquatic animal health and welfare, particularly for captive animals. For sea turtle, bacterial infection is one major problem that causes a high rate of disease and death in both free-living and captive sea turtles [2-6]. It has been shown that bacteria in water together with other parameters have significant impact on the survival rates of fish [7, 8]. In closed seawater systems e.g. aquaria, both biotic and abiotic environmental factors have great effect on the bacterial population in the water [9]. Several bacterial species associated with captive sea turtles have been identified including saprophytes and potential pathogens [2,5]. However, factors associated with bacterial concentration and diversity in captivity are yet to be investigated.
Live rock, a term commonly used in the aquarium trade, is a dead coral skeleton usually covered by encrusting coralline algae and harboring various invertebrate organisms [10]. Live rock is popular for closed marine ecosystems as it can maintain water quality via the nitrogen removal of bacteria colonizing the rock and helps to stabilize the water chemistry [11]. Nevertheless, the information on the association between live rocks and heterotrophic bacterial growth in seawater is very limited. Therefore, the objective of this study was to determine the effects of live rocks, stock density, and other variables including pH and temperature of the water on the seawater quality in juvenile green turtle captivity. In this investigation, aerobic bacterial populations and water quality parameters of the seawater were compared among different captive environments.
Experiment conditions and rearing protocol
The experiments were performed at the Sea Turtle Conservation Centre, Sattahip, Chonburi Province. Three different conditions in juvenile green turtle captivity based on stock density, captivity size, and presence of live rocks were used in the study (Table 1). The uncured live rocks with random sizes of dead coral skeleton fragments encrusted with coralline algae collected from Sattahip Bay, Chonburi Province, Thailand (12° 40’ North, 100° 54’ East) were placed directly in the captivity to cover the entire bottom.

Table 1: Details of rearing conditions and environments used in the study.
The turtles were maintained in smooth-surfaced containers and fed twice a day with fish meat stored in a -20°C freezer. The seawater in containers 1 and 3 was changed twice a day, after morning feeding time and in the late afternoon by direct pumping from the sea near the conservation centre to the basins without any treatment. For container 2, there was no water changing but the water was aerated continuously using air pump. To analyze the bacterial levels, seawater of each basin was collected at a 10 cm depth from the surface by using a 15 ml sterile tube and kept on ice until examination. The sampling of each treatment was at every 2 weeks interval from February 2009 to June 2009, in duplicate, yielding 14 water samples. Temperature and pH of the samples were measured at every sampling.
Bacterial investigation
Enumeration of aerobic bacteria in seawater samples was carried out using spread plate technique. The seawater was subjected to a series of 10-fold dilutions using sterile 0.9% NaCl before 1 ml of each non-diluted (100) and diluted sample (10-1 to 10-6) was plated on nutrient agar, supplemented with 1% NaCl (NA+1%NaCl) and with 3% NaCl (NA+3%NaCl) for heterotrophic marine bacteria [12]; blood agar (BA) for pathogenic haemolytic bacteria; Mannitol salt agar (MSA) for halophiles and for differentiating pathogenic and nonpathogenic Staphylococcus spp. [13]; MacConkey agar (Mac) for gram-negative bacteria [14]; Eosin methylene blue agar (EMB) for gram-negative enteric and coliform bacteria [15,16]; Xylose-lysine deoxycholate agar (XLD) for Salmonella and Shigella spp. [15]; and Thiosulfate citrate bile salt sucrose agar (TCBS) for Vibrio spp. [17-19]. All plates were incubated at 25C for 48 h and estimation of the bacterial number was according to the colony-forming unit (CFU) method. Confirmation of bacterial identification was performed on randomly selected colonies using standard bacterial taxonomy procedures according to the 8th edition of Bergey’s Manual of Determinative Bacteriology [20].
Statistical analysis
The average values are shown in Table 2. Statistical analysis was carried out to evaluate differences in the mean abundance of each bacterial group, according to bacterial counts from each medium, among three treatments as well as between sampling periods by one-way ANOVA. When a significant difference (p<0.05) of the bacteria levels was observed, the means were analysed by Tukey’s HSD test.
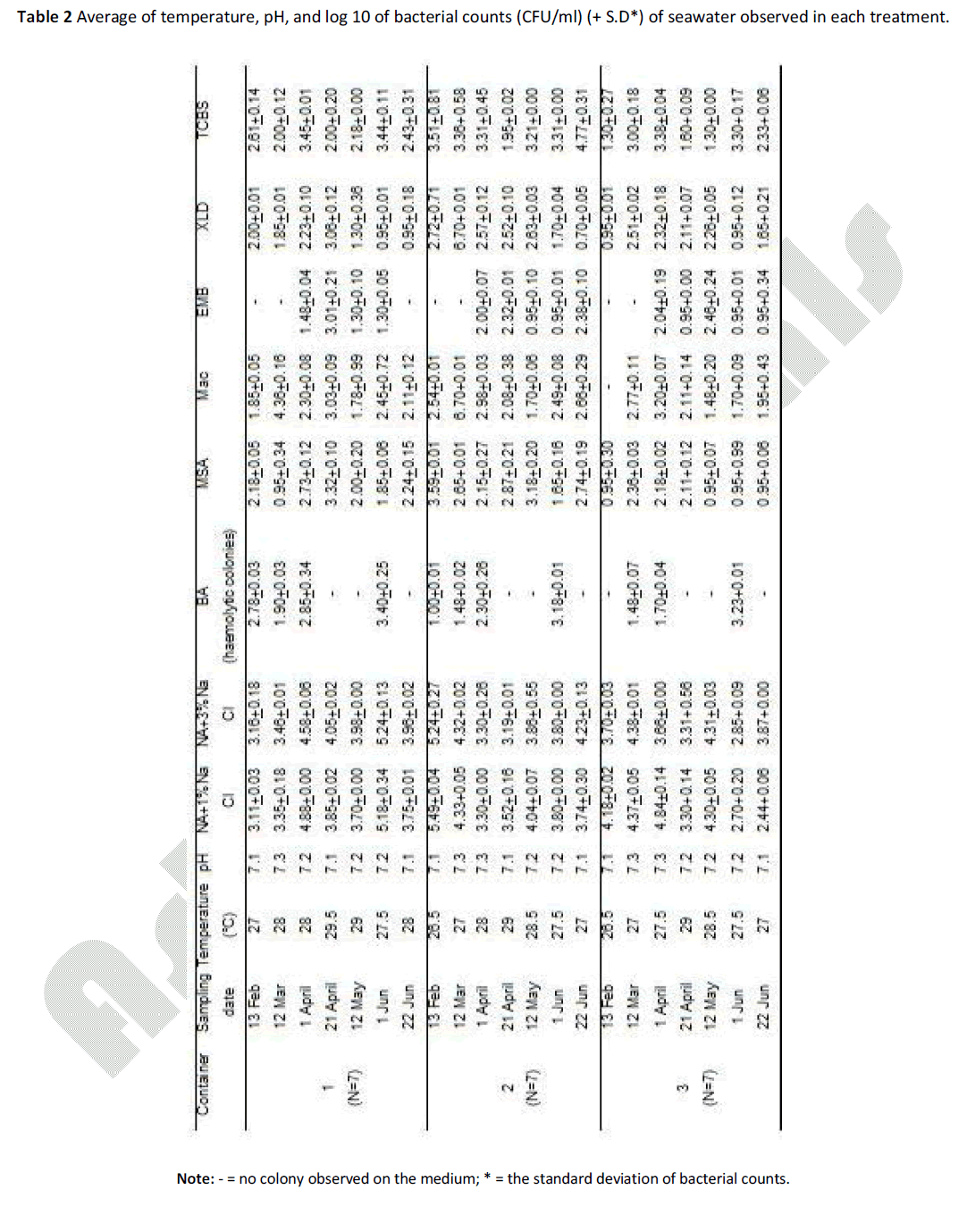
Table 2: Average of temperature, pH, and log 10 of bacterial counts (CFU/ml) (+ S.D*) of seawater observed in each treatment.
Abiotic characteristics of seawater
In Table 2, the average water temperature was from 26.5°C to 29.5°C; the highest temperature was during April and May 2009 (between 28°C and 29.5°C). The pH of the seawater from all containers showed low fluctuation, ranging between 7.1 – 7.3 (Table 2), which was similar to the natural seawater pH.
Comparisons of microbial levels in seawater among containers
The results from statistical analysis revealed no significant difference of heterotrophic marine bacterial numbers in the water on NA + 1% NaCl and 3% NaCl media between three containers (ANOVA, p>0.05), usually around 3 to 4 log CFU/ml (Table 2). Microbial counts of the water from every container observed in all 6 selective media generally ranged from 1 to 3 log units of CFU/ml (Table 2).
No significant difference of bacterial levels in the water from 3 containers was observed in all selective media (ANOVA, p>0.005). Comparisons of microbial counts of each media between 3 containers indicated that the seawater of container no. 3 always contained low microbial levels. Generally, container no. 2 showed the highest values of bacterial counts (in 4 cases out of 7 samples), except for BA medium where the highest bacteria counts of haemolytic colonies were from the water in container no. 1. It is also noteworthy that for TCBS medium, the Vibrio counts from the water of container no. 2 (approximately 3 log units) were in most cases 1 to 2 order of magnitude higher than the counts from container no. 1 and 3 (around 1 to 2 log units).
Comparisons of microbial levels in seawater between sampling periods
Significant differences in bacteria counts averaging from three container water between sampling dates were obtained from BA, XLD, and TCBS media by using one-way ANOVA and Tukey’s HSD test (p=0.000, p=0.026, and p=0.028, respectively) (Fig. 1). On BA and TCBS media, haemolytic and Vibrio counts in the water of all three containers during April and May obviously decreased (Fig. 1a and c), while decrease in Shigella and Salmonella levels (on XLD medium) was seen in the seawater samples collected in June.
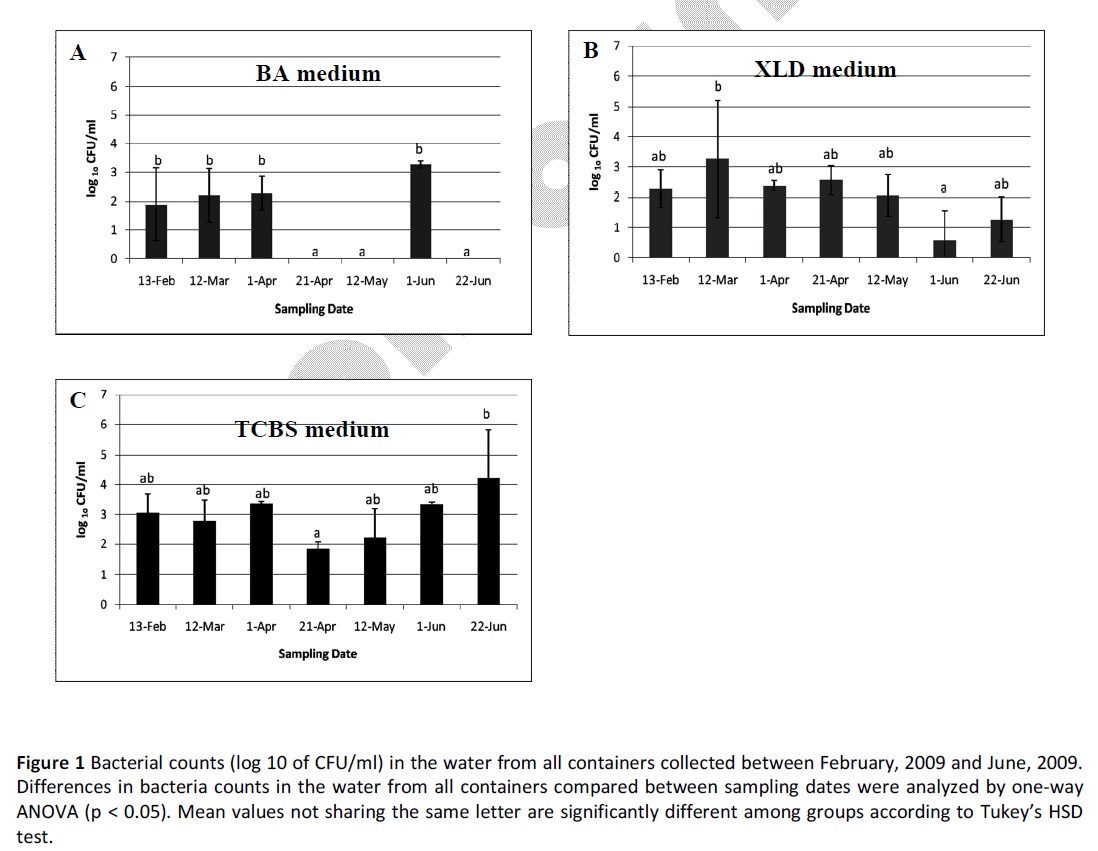
Figure 1: Bacterial counts (log 10 of CFU/ml) in the water from all containers collected between February, 2009 and June, 2009. Differences in bacteria counts in the water from all containers compared between sampling dates were analyzed by one-way ANOVA (p < 0.05). Mean values not sharing the same letter are significantly different among groups according to Tukey’s HSD test.
The experiments conducted in this study showed that the presence of live rocks in juvenile green turtle containers maintained low levels of haemolytic bacteria in seawater, implying its ability for bacterial growth inhibition. This finding may have implications for further development of live rock utilization in aquaculture, possibly for regulating the bacterial compositions of the community, which give benefits to the system such as maintaining low levels of some bacteria that have pathogenic properties to the animals (see below). We also observed that stock density of animals in captivity is another parameter affecting the bacterial levels in seawater with subsequent disease problems.
It is clear that both live rock and captivity environments have no effect on the pH of seawater in the juvenile green turtle containers. In Table 2, the results indicated that replacing live rocks and coral skeletons in the captivity clearly reduced bacteria growth in the water (container no. 2) on BA medium, which is generally used to distinguish normal from pathogenic bacteria based on the haemolytic characteristic of red blood cells, compared to the container no. 1 that had the same size and stock density. Although live rock has been noted for its ability of nitrification and denitrification, there has been no available information on the involvement of such activity in growth reduction of heterotrophic marine bacteria. To our knowledge, this is the first study on the live rock effect on bacterial growth in seawater and it is interesting to further elucidate the best use of live rock in aquaculture application.
On the other hand, the water from container no. 2 always showed a high number of bacteria counts for other media, including MSA, Mac, XLD, especially for TCBS. Some facultative anaerobic halophilic bacteria such as Vibrionaceae can use nitrate as alternative electron receptor for oxidative metabolism. Therefore, the bacteria colonizing live rocks may directly supply nitrate to the Vibrio population enhancing the growth rate of this bacterial genus, and this might explain high levels of Vibrio spp. in this container. Alternatively, the live rocks themselves may have no any effect on the bacterial growth and perhaps high bacteria levels in the container are simply due to accumulation of the bacteria in the live rock and coral skeleton structures.
In aquatic systems, nutrient levels strongly influence bacterial growth, growth efficiency, and bacterial community composition. Several reports found that changes in nutrient levels exert either positively or negatively on bacterial growth depending on bacterial strains, thereby changes in bacterial compositions [21-23]. Similarly, in this study a lower abundance of haemolytic bacteria in a container with live rocks can be explained by the dynamics of the bacterial community resulting from the coralline algae, invertebrate organisms, and denitrifying/nitrifying bacteria inhabited in the live rocks which may play important roles in the nutrient cycles of the communities such as particular organic and inorganic nutrients [24,25]. Somehow, the nutrient levels due to shifts of these substrate suppliers do not support the growth of haemolytic bacteria in the water of the container.
Haemolytic bacterial and Vibrio spp. numbers in the water of all 3 containers were significantly different when the time of sample collection was considered (Fig. 1a and c). Decrease in these bacterial counts were obviously seen during April and May, the summer season of Thailand, which commonly has the highest average temperature of the year (Table 1). From our preliminary data, we found that incubation of all cultures at a higher temperature (37C) resulted in much slower growth rates of bacteria than at a lower temperature (25°C). Moreover, several reports indicated that the survival and growth rates of some Vibrio species in seawater and animals including Vibrio alginolyticus, the most common bacterial type found in bacteria-infected organs of Thai juvenile green turtles [2], have been strongly influenced by the temperature of the water [26-30]. This implies that temperature, a physical factor, has great impact on haemolytic bacterial and Vibrio spp. growth in the seawater of sea turtle captivity.
As expected, the microbial numbers in seawater of container no. 3 was found to be at low levels as it had very low stock density of animals compared to the other containers (Table 1), where high stock density or over-crowding is known to be a major cause of the health problem of aquatic animals in captivity [31]. The results in Table 2 demonstrated that stock density in captivity is another parameter that directly affects the microbial levels in seawater.
Several bacterial genus and strains have been evaluated as markers of microbiological water quality. By using the filtration method and sediment analysis [15], we could not detect Escherichia coli in the water samples, both from natural environment and from the sea turtle containers (data not shown). This may be because in marine habitats the survival of Vibrio spp. is longer than that of E. coli [32]. For this reason, more microbiological controls are recommended for completion of water quality assessment. Vibrio spp. has been used widely as an indicator for various purposes including aquaculture activities, environmental concern, and pathogenic aspects for humans as well as aquatic animals. For the Sea Turtle Conservation Centre, Thailand, the water from the captivity has moderate numbers of Vibrio spp. (ranging from log cfu/ml 1.30 to 4.77), when compared with the Vibrio counts from the mussel water farm at the Mar Piccolo, Ionian Sea, Italy (log cfu/ml at 1.0 - 2.4) [17]; and at the Gulf of Gaeta (Tyrrhenian Sea), Italy (log cfu/ml 3.03 – 3.95) [18]; and from the aquaria in Spain (log cfu/ml 3.60 - 4.34) [9].
The experiments conducted in this study showed that the presence of live rocks in juvenile green turtle container maintained low levels of haemolytic bacteria in seawater, implying its ability in bacterial growth inhibition. This finding may have implications for further development of live rock utilization in aquaculture.
Stock density of animals in captivity and temperature of the water are other parameters affecting bacterial levels in seawater that could lead to disease problems. The data obtained from this research will be useful fundamental information for further study on the live rock application for aquaria and for the establishment of suitable environments required for aquatic animal husbandry.
The authors declare that they have no competing interests.
TC carried out the study design, bacterial investigations, and drafted the manuscript. PP participated in bacterial investigations and helped to draft the manuscript. KP performed the statistical analysis.
This research was supported by a grant from Faculty of Science, Silpakorn University. We are grateful to the Sea Turtle Conservation Centre along with the Air and Coastal Defense Command, the Royal Thai Navy for their kind support in providing the samples. We also thank S Nimgirawath (Department of Chemistry, Faculty of Science, Silpakorn University) and M Areekijseree (Department of Biology, Faculty of Science, Silpakorn University) for critical discussion on the manuscript.