Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2015) Volume 4, Issue 3
Essential oils are commonly used as natural remedies in many areas of the world. However, studies investigating the effects of essential oils at the cellular and molecular level are lacking. This work was undertaken to evaluate the effects of essential oils on mRNA expression in human cells. The liver HepG2 cell line was treated with 31 different essential oils, and then assessed for gene expression changes using RNA-seq. Several well characterized transcription factors were identified as being differentially regulated upon general treatment with an essential oil. Additionally, genes and pathways unique to a particular essential oil, or subset of essential oils, were identified. Importantly, genes and pathways identified through this study suggest that essential oils induce pathways that have the potential to promote cell growth and self-defense, or induce cell death, depending on the context. Follow-up experiments further validating and verifying the biological activity of essential oils in multiple cell types need to be performed.
Keywords: RNA-seq, Essential oils, HepG2 cells, Transcriptome, Isolation
Evaluation of natural plant products as potential remedies for common ailments is a fast growing segment of product research and development. One natural product segment that currently has considerable interest is the use of essential oils (EOs). EOs is natural aromatic compounds isolated from plants. Although much anecdotal evidence exists for the usefulness of these products, the research to provide actual mechanisms of action and scientific support for these products is severely lacking. Gene expression studies, followed by molecular pathway analysis, are a common method to provide data for hypothesis generation and testing of the effects of a test substance. The contemporary method to investigate changes in the transcriptome is through RNA-seq. RNA-seq is a technology that uses the capabilities of next-generation sequencing to reveal a snapshot of RNA species and quantity from a genome at a given moment in time. We set out to evaluate the effects of a large panel of essential oils on the transcriptional landscape of human cells. Although there are many relevant cell types that could be examined in such a study, we chose the well characterized HepG2 liver cells for a model, as the liver is designed as a removal organ for foreign substances and HepG2 cells show dramatic changes in transcriptional regulation when treated with products of interest [1,2].
Essential oils
Thirty one essential oils (peppermint [Mentha piperita], bergamot [Citrus Bergama], wintergreen [Gaultheria procumbens], grapefruit [Citrus x parasisi], basil [Ocimum basilisum], rosemary [Rosmarinus officinalis CT cineol], orange [Citrus sinensis], melaleuca [Melaleuca alternifolia], clove [Syzygium aromaticum], lemon [Citrus limon], balsam fir [Abies balsamea], birch [Betula alba], chamomile [Chamaemelum nobile], cinnamon [Cinnamomum verum], cypress [Cupressus sempervirens], eucalyptus globulus [Eucalyptus globulus], eucalyptus radiate [Eucalyptus radiate], frankincense [Boswellia carteri], ginger [Zingiber officinale], helichrysum [Helichrysum italicum], lavender [Lavandula angustifolia], lemongrass [Cymbopogon flexuosus], marjoram [Origanum majorana], myrtle [Myrtus communis], oregano [Origanum compactum], sandalwood [Santalum album], spruce [Picea mariani], tangerine [Citrus reticulate], thyme [Thymus vulgaris], vetiver [Vetiveria zizanoides], ylang ylang [Cananga odoratac]) were procured through Améo (Lehi, UT, USA).
Cell culture
HepG2 cells were grown in RPMI media supplemented with 10% fetal bovine serum, and 1% Penicillin/Streptomycin solution in a humidified incubator with 5% CO2. HepG2 cells were plated in 6-well plates and allowed to attach overnight. Cells were then treated with EOs and incubated for a period of 6 hours.
RNA preparation and sequencing
RNA was isolated from cells using the mirVana RNA preparation kit (Life Technologies, Carlsbad CA). mRNAs were then purified using Poly(A) selection from the total RNA sample, and then fragmented. First strand cDNA was synthesized using random priming, followed by the synthesis of the second strand of cDNA. The resulting doublestrand cDNA was end repaired, phosphorylated and A-tailed. Library preparation was performed according to the manufacturer’s protocol (Illumina, San Diego CA) for library preparation. Raw fasta files from the sequencing instrument were quality trimmed to remove poor quality ends and adaptors removed. Tophat2 was used to align the cleaned reads to the human genome (UCSC assembly Hg20). Ht Seqcount was used to count reads mapped to exons [3]. Then, differential gene expression analysis was conducted using edgeR [4]. All zero hits genes were removed, 38179 genes has at least I hit count. Those without 1 cap were removed. A total of 14224 genes remained. Differential gene expression analysis was performed using edgeR. Heat maps were drawn with CIMminer using normalized RPKM values.
Determination of EO concentration for pathway induction
Each particular EO is composed of a unique compliment of constituents, and thus potentially vary in their effects on living cells. Therefore, the first step was to determine a dose high enough to elicit a biological response, yet low enough to avoid the induction of cell death. A dose finding study was performed to determine the concentration of EO that would decrease viability by 20% at a treatment length of 72 hours (IC20). This empirically determined concentration was carried out for each essential oil and used in the subsequent expression studies (recorded in Table 1).
| Essential oil | Oil%vol/vol | Essential oil | Oil%vol/vol |
|---|---|---|---|
| Peppermint | 0.050 | Eucalyptus radiata | 0.030 |
| Bergamot | 0.032 | Frankencense | 0.019 |
| Wintergreen | 0.050 | Ginger | 0.121 |
| Grapefruit | 0.100 | Helichrysum | 0.046 |
| Basil | 0.148 | Lavender | 0.051 |
| Rosemary | 0.029 | Lemongrass | 0.012 |
| Orange | 0.074 | Marjoram | 0.009 |
| Melaleuca | 0.014 | Myrtle | 0.006 |
| Clove | 0.010 | Oregano | 0.004 |
| Lemon | 0.020 | Sandalwood | 0.006 |
| Balsam fir | 0.010 | Spruce | 0.016 |
| Birch | 0.035 | Tangerine | 0.060 |
| Chamomile Roman | 0.013 | Thyme | 0.011 |
| Cinnamon Bark | 0.003 | Vetiver | 0.026 |
| Cypress | 0.034 | Ylangylang | 0.052 |
| Eucalyptus Globulus | 0.005 |
Table 1: Essential oil IC20 concentrations in HepG2.
General effects of essential oil treatment on HepG2 cells
Following the determination of the EO IC20 concentrations, HepG2 cells were treated with EOs for a period of 6 hours. Following treatments, cells were harvested and RNA was isolated for expression studies. RNA was prepped and underwent next-generation sequencing as described in the materials and methods section. RPKM values were determined and set relative to the untreated control samples.
To gain a sense of the general transcriptional changes taking place in EO treated cells, the median relative expression values across all EOs were calculated for each gene/transcript and rank ordered. This analysis identifies transcripts that are most differentially regulated in a majority of the EOs tested. Overall, there were 209 transcripts identified as up-regulated by 2-fold or greater, and 129 transcripts that were down-regulated by 2-fold or greater. The 50 most up-regulated transcripts and 50 most down-regulated transcripts are listed in Table 2. Several well-known proteins of the AP-1 complex are notably upregulated in the EO treated cells. AP-1 is a heterodimeric transcription factor composed of proteins belonging to the c-Fos, c-Jun, ATF and JDP families. It regulates expression in response to a variety of stimuli, such as stress, growth factors, cytokines, and microbial infections [5]. AP-1 pathway signaling is contextual and can promote differentiation, proliferation, and/or apoptosis [6]. We observed general up-regulated expression of FOS, FOSB, FOSL1, and JUN in EO treated cells, which could be contributing to cell growth, or cell death. A study by Dosmorov, et al. examined the effects of frankincense and sandalwood on cultured bladder cells. Although they reported that the two oils activated unique pathways, it was noted that both oils activated FOSB, JUN, JUNB, EGR1, and ATF3 [7]. Furthermore, another study found that the EO and active fraction from coriander (Coriandrum sativum L.) was found to up-regulated c-Jun, c-FOS, c-JUN/C-FOS and AP-1 [8]. Therefore, our results further corroborate and extend these findings to include HepG2 cells and a much larger breadth of EOs with this property.
| 50 most down-regulated transcripts following addition of EO | 50 most up-regulated transcripts following addition of EO | ||
|---|---|---|---|
| LRRC25 | RP11-326C3.13 | FOS | ZFP36 |
| NFE2 | ALDH8A1 | EGR1 | LINC00659 |
| LRRN4 | FOXJ1 | FOSB | IL11 |
| RP11-549B18.1 | RP11-350N15.5 | SLC7A11 | DNAJB9 |
| NANOS1 | DUSP5P1 | DUSP5 | MCL1 |
| LIM2 | C1orf210 | RP11-499O7.7 | PLK3 |
| SOWAHB | GALNT6 | HMOX1 | KDM7A |
| AC006273.5 | AC012065.7 | GDF15 | HBEGF |
| KLHDC7A | EPN3 | CXCL8 | FOSL1 |
| MIR135A1 | RP11-1008C21.2 | RASD1 | CSGALNACT2 |
| FAM111B | RP11-110I1.12 | DUSP1 | LIF |
| TMPOP2 | CEP83-AS1 | SPRY4 | GCLC |
| LRRC26 | NTN4 | EREG | EID3 |
| TMPO-AS1 | RP11-572O17.1 | CPEB4 | TNFSF9 |
| CTD-2555C10.3 | AC011242.6 | EGR4 | TRIB1 |
| C17orf82 | EPO | MXD1 | RP11-325K4.3 |
| RP11-568A7.2 | TMEM139 | PPP1R15A | CEBPB |
| RP5-836N17.4 | RP13-890H12.2 | MIR3189 | DNAJB4 |
| ZNF517 | RP11-529E10.6 | JAG1 | RP11-345L23.1 |
| SOWAHD | FAM84B | RP11-325K4.2 | NR1D1 |
| E2F2 | TIGD3 | HSPA6 | LURAP1L |
| AC141928.1 | FZD8 | SERPINE1 | JUN |
| CTB-193M12.4 | FTOP1 | GCLM | CCL20 |
| CYP1A1 | PRR15 | ERRFI1 | RP5-1142A6.9 |
| TUSC8 | HOXA3 | AREG | MMP3 |
Table 2: General expression effects of EOs on HepG2 cells.
To gain more insight into the molecular pathways being affected by EO treatment, functional annotation by the DAVID gene ontology tool [9] was performed. Using the 338 transcripts differentially regulated at greater than 2-fold relative to untreated cells, it was revealed that pathways related to the regulation of transcription had the lowest p-values and were predicted to be the most significant (Table 3). When taking only the down-regulated genes into account, the DAVID ontology tool produced similar GOTERM results compared to the combined list in Table 3, with several genes involved in transcription noted. These down-regulated transcripts include: E2F2, E2FB, BATF2, CBX2, EPO, FOXJ1, HOXA3, HOXA4, NFE2, OTX1, SALL2, UHRF1, and ZNF517. When taking only the up-regulated genes into account, the biological process GOTERM for response to organic substances ranked the highest. The up-regulated genes identified as important for this process were DNAJB4, FOSL1, KLF10, PLIN2, ADM, ASNS, DUSP1, EGR1, ERN1, GCLC, GOT1, HSPA6, HMOX1, IGFBP1, ITGA2, JUNB, JUN, LDLR, MMP3, MCL1, PCK1, PPP1R15A, SOCS3, THBS1, TRIB1, FOS, and MYC.
| DAVID (6.7) Top GETERM biological processes | P-Value | |
|---|---|---|
| 1 | Regulation of transcription from RNA polymerase II | 1.10E-08 |
| 2 | Regulation of transcription | 1.20E-08 |
| 3 | Regulation of RNA metabolic process | 2.10E-08 |
| 4 | Regulation of cell proliferation | 6.40E-08 |
| 5 | Regulation of transcription, DNA-dependent | 7.90E-08 |
| 6 | Positive regulation of macromolecule biosynthetic process | 2.40E-07 |
| 7 | Response to organic substance | 4.50E-07 |
| 8 | Positive regulation of cellular biosynthetic process | 5.90E-07 |
| 9 | Positive regulation of biosyntheic process | 7.80E-07 |
| 10 | Positive regulation of macromolecule metabolic process | 1.30E-06 |
Table 3: Gene ontology predictions from differentially regulated genes.
The 338 transcripts identified as generally differentially regulated among the EOs were examined using hierarchical clustering (Figure 1) to determine which oils group together with respect to differential gene expression. There were 2 main clades observed, one that clustered with the 2 untreated controls, and one that had more pronounced expression changes relative to the controls. However, it should be noted that differential expression was similar for most of the transcripts among the 31 essential oils tested, supporting the idea that there is a general response to essential oils in HepG2 cells.
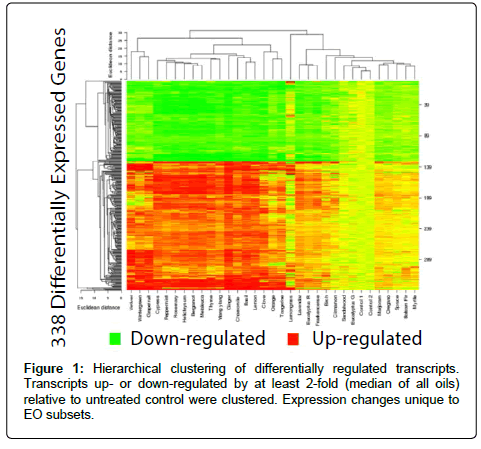
Figure 1: Inhibitory effect of the Morinda citrifolia 70% aqueous-ethanolic fruits (Mc.Cr.F) (A) and leaves extract (Mc.Cr.L) (B) on spontaneous and high K+- induced contractions and concentration-response curves of Ca++ in isolated rabbit jejunum. The symbols represent mean ± S.E.M, n= 4-6.
After determining a general sense of transcripts differentially regulated among EO treated cells, the next step was to further investigate expression profiles for changes that are unique to particular oil or to a subset of EOs. To determine the variation among the EO treatments the standard deviation of relative transcription levels was calculated for each gene. There were 242 genes that had greater than a 2-fold standard deviation. HSPA6, FOS, PCK1, EGR1, and FOSB were the 5 genes that showed the most variation among the EOs. Compared to the 338 genes identified as generally differentially expressed, 101 genes overlapped, meaning that 141 genes exhibited high variation between samples, but were not identified as up- or down-regulated. Table 4 provides an example of some interesting genes observed from the 141 genes identified. Interestingly, there are many similarities in expression patterns when compared with the hierarchical clustering performed using the set of differentially expressed genes. Vetiver, wintergreen and grapefruit treatments induced expression of CXCL2, CXCL3, HSPA1B, and DDIT3 to a much greater extent than the other EOs. Cypress, peppermint, rosemary, helichrysum, bergamot, and melaleuca formed a group in the previously performed differential expression hierarchical clustering, and also exhibited a unique pattern in this analysis comparing genes identified as variably expressed but not considered generally differentially regulated (Table 4). These 6 EOs exhibited among the lowest expression levels of the stress inducible genes CXCL2, CXCL3, and HSPA1B. In contrast, these 6 EOs also exhibited among the highest levels of UCN2 and IGFBP3. UCN2 and IGFBP3 are negative regulators of growth signaling and play a role in stress adaptation [10,11].
| Relative expression values (Log 2) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| CXCL2 | CXCL3 | HSPA1B | DDIT3 | RGS2 | UCN2 | IGFBP3 | PLK2 | CYR61 | |
| Vetiver | 7.5 | 6.0 | 5.1 | 5.7 | 2.9 | 1.3 | 0.5 | 2.6 | 2.5 |
| Wintergreen | 5.6 | 6.2 | 4.6 | 2.9 | 4.7 | 0.6 | -0.2 | 3.4 | 3.1 |
| Grapefruit | 5.9 | 5.1 | 3.8 | 4.8 | 2.9 | 1.6 | -0.2 | 3.7 | 1.3 |
| Cypress | 0.3 | 0.9 | 0.0 | 1.7 | -0.4 | 3.1 | 2.7 | 1.0 | 0.9 |
| Peppermint | -0.3 | 0.5 | 0.1 | 0.6 | 0.7 | 2.8 | 2.3 | 0.3 | 2.3 |
| Rosemary | -0.7 | 0.0 | -0.3 | 0.6 | 0.4 | 2.9 | 1.8 | 0.9 | 2.7 |
| Helichrysum | 0.2 | 0.0 | 0.0 | 1.5 | 0.6 | 3.5 | 2.4 | 0.4 | 1.8 |
| Bergamot | -1.0 | 0.4 | -1.9 | 0.5 | 0.1 | 3.1 | 2.0 | 0.6 | 2.2 |
| Melaleuca | -0.2 | 0.2 | -1.7 | 0.8 | 0.7 | 3.4 | 2.8 | 0.7 | 2.6 |
| Thyme | 0.6 | 2.1 | -1.0 | 2.8 | 0.7 | 1.1 | 2.0 | 0.1 | 1.6 |
| Ylangylang | 0.7 | 1.0 | 0.0 | 1.8 | 0.8 | 2.7 | 2.0 | 0.1 | 0.9 |
| Ginger | 1.8 | 1.9 | -0.4 | 3.4 | 0.7 | 3.8 | 2.3 | 1.1 | 2.4 |
| Chamomile | 2.1 | 3.6 | -0.3 | 4.5 | 0.8 | 0.4 | 1.3 | 1.0 | 1.6 |
| Basil | 2.8 | 3.9 | 2.7 | 3.3 | 2.2 | 1.6 | 0.6 | 2.6 | 2.0 |
| Lemon | 0.1 | 0.4 | 2.1 | 3.5 | 0.8 | 1.8 | 1.2 | 1.0 | 0.2 |
| Clove | 3.5 | 3.3 | 2.4 | 3.8 | 1.0 | -0.5 | -0.2 | 0.3 | 0.2 |
| Orange | 2.1 | 2.9 | 2.1 | 3.7 | -0.8 | 0.4 | -0.2 | 0.2 | 0.6 |
| Tangerine | 0.0 | 2.4 | 0.2 | 3.8 | -0.8 | 0.6 | 0.5 | -0.4 | 0.3 |
| Lemongrass | 2.6 | 3.7 | 4.9 | 2.7 | 5.0 | -0.2 | -0.2 | 2.7 | 1.5 |
| Lavendar | -1.0 | 0.0 | 0.7 | 0.6 | -0.3 | 1.7 | 1.4 | 0.4 | 0.9 |
| Eucalyptus R. | -0.4 | 0.0 | 0.7 | 0.5 | 0.5 | 1.0 | 1.4 | 0.3 | 0.4 |
| Frankencense | -1.0 | 0.0 | 0.9 | 0.5 | 0.6 | 0.4 | 1.6 | 0.1 | 0.1 |
| Birch | -0.1 | 0.0 | 0.0 | -0.2 | -0.6 | 0.0 | 0.4 | -0.1 | -0.1 |
| Cinnamon | -1.0 | 0.0 | 1.9 | 0.1 | -0.1 | -0.4 | -0.2 | -0.1 | -0.2 |
| Sandalwood | -1.0 | 0.0 | -0.1 | 0.1 | 0.4 | 0.0 | 0.1 | 0.1 | -0.1 |
| Eucalyptus G. | 0.2 | 0.0 | 0.6 | 0.2 | 0.0 | -0.9 | 0.2 | -0.1 | -0.5 |
| Marjoram | 0.3 | 0.0 | 0.9 | 0.4 | 0.4 | 0.3 | 0.2 | -0.1 | -0.1 |
| Oregano | -0.6 | 0.8 | 0.3 | 0.1 | 0.9 | -0.2 | 0.3 | 0.2 | 0.4 |
| Spruce | -0.4 | 0.2 | 0.1 | 0.2 | 0.1 | 0.2 | 1.4 | 0.0 | -0.3 |
| Balsam Fir | 0.5 | 0.0 | 0.3 | 0.1 | -0.4 | 0.5 | 0.4 | 0.0 | -0.2 |
| Myrtle | 0.2 | 0.0 | 0.3 | 0.5 | 0.0 | 0.4 | 0.0 | 0.2 | 0.0 |
Table 4: Genes with high variation, but not identified as generally differentially expressed.
Analysis of expression patterns can also be done in a forward manner by evaluating EOs with similar constituents. Of the 31 EOs tested, 5 were isolated from citrus plants, with limonene being the known major constituent of these oils. Therefore, it is likely that differences exist in transcriptional regulation between the 5 citrus treated samples relative to the other EOs. To explore this possibility, the relative expression for the 5 citrus oils was averaged and compared to the average relative expression of the other 26 treated samples. The difference was calculated and all transcripts that differed by 2-fold or more were further investigated to determine if these differentially regulated transcripts were unique to citrus oils, or alternatively, if citrus oils have a more dramatic effect on general expression changes observed with EO treatment. Overlapping genes depicted by Venn diagram in Figure 2 show that roughly half of the up-regulated genes in citrus were also up-regulated in the other oil treatments. Of the 31 unique elements up-regulated in citrus oils, the metallothioneins and the solute carrier family (SLC) gene families were observed. Metallothioneins are proteins produced at high levels in the liver, and are thought to be protective by binding to heavy metals with high affinity [12,13].

Figure 2: Gene signatures of citrus oils. Venn diagram showing number of genes highly up-regulated (A), or highly down-regulated (B) in citrus oils compared to non-citrus oils. Roughly half of the genes identified as up-regulated in citrus oils were also found up-regulated in the other 26 oils tested. Whereas the majority of down-regulated genes found in citrus were unique to citrus EOs.
Many natural products including flavonoids induce the expression of metallothioneins in HepG2 cells [14]. However, metal toxicity can also induce the expression of metallothioneins [15]. Therefore, it would be interesting to investigate the induced expression of metallothionens by citrus oils more in depth. Of the 25 elements down-regulated in citrus oils, 20 were found to be exclusive to citrus and not found differentially expressed in the other 26 oils. The development associated distalless homeobox 1 (DLX1) and homeobox D1 (HOXD1) genes were among the genes in the list that were unique to citrus. There were 5 overlapping down-regulated genes between the citrus and non-citrus oils. These genes are therefore down-regulated across most EOs, but to a much greater extent in citrus oils. These genes were CYP1A1, MT-TD, TIGD3, LRRC25, and NONOS1. Kalmes et al. examined aryl hydrocarbon receptor translocation to the nucleus and CYP1A1 upregulation following eugenol or isoeugenol treatment in HaCaT cells [16]. However, we observed lower expression of CYP1A1 for all 31 essential oils tested. It is possible that keratinocytes and hepatocytes are different in that regard.
Similar to the citrus oil analysis, lemongrass oil was compared to the other EOs, since it was the most dissimilar oil tested. Lemongrass did not seem to cluster with any other oil, exhibiting several transcripts that were up-regulated upon treatment, which corresponded to downregulated transcripts when treated with the other 30 oils. Compared to the relative expression values of the other 31 oils, 61 genes were found to be up-regulated uniquely in lemongrass. Of the 61 genes, there were several histone subunits, and heat shock proteins. With respect to down-regulated genes identified as unique to lemongrass treated cells, there were 58 genes that exhibited a 2-fold difference relative to the average of the other 30 oils. Of those 58 genes, 54 appeared to be unique to Lemongrass. Interestingly, 14 of the 54 genes identified were mitochondrial genes specific for tRNA. Given that tRNA expression is thought to play a role in apoptosis regulation [17], together with the observation of heat shock proteins up-regulation, we suspect that the lemongrass treated cells were undergoing pressures of stress not observed in the other treated samples. Therefore, it would be important to conduct further studies with a gradient of concentrations for lemongrass to see if lower concentrations give similar results. Interestingly, lemongrass has been reported to have anticancer activity [18], and HepG2 cells might be particularly sensitive to this EO.
RNA-seq is a powerful method to determine mRNA expression levels in cells. We were able to use this method to evaluate the expression changes induced by a comprehensive set of essential oils in HepG2 cells. Gene ontology analysis suggests that AP-1 transcription up-regulation is a general effect of EO treatment in HepG2 cells. Therefore, essential oils might influence growth and rejuvenation of particular types of cells. We have also identified groups of EOs that cluster together based on transcriptional profiles. We are continuing to validate these candidate genes and EO groupings to enable and guide the development and usage applications for EOs.