Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2016) Volume 3, Issue 4
In order to study the effect of external ethylene on postharvest display and activity of some senescence-related enzymes in potted miniature roses, this experiment was carried out in a completely randomized design, using three treatment levels with three replications. In glass chambers, potted miniature rose plants were exposed to ethylene concentrations of 0, 5 and 10 μL L-1 for 12 h. The results of study showed that the application of ethylene affected morphological and physiological properties and activity of antioxidant enzymes, as well as laccase enzyme in treated plants. Stress of ethylene in miniature roses caused to increase the percentage of leaf abscission and laccase enzyme activity. Studying defensive mechanism by measuring antioxidant enzymes showed that after using stress of ethylene activity of peroxidase enzyme was gradually decreased. Ethylene concentration of 10 μL L-1 caused to increase activity of peroxidase enzyme in primary days after exposing to ethylene stress but enzyme activity was gradually decreased till the final days. In other side, increasing peroxidation rate of membrane lipids under ethylene stress condition shows plant inability to eliminate or balance oxygen free radicals. Activity of laccase in stressed plants was increased significantly during time, concomitant with decreasing chlorophyll and proline content.
<
Keywords: Abscission; Chlorophyll; Copper-containing oxidase enzyme; Lipid peroxidation; Proline
Miniature roses (Rosa hybrida ) are well known as one of the world’s most popular ornamental plants grown world widely as potted and plants. Postharvest display quality and life of flowers are key factors, which are considered in acceptance of miniature rose in international marketing trade especially for exporting to distance markets. In commercial view, yellowing of leaf and falling leaf, flower and bud organs decrease popularity of potted plant especially roses [1]. Ethylene, gaseous plant hormone, has most important role in plant developmental stages as well as regulating the aging process in plant organs. Moreover, the plant organs are active in self-regulation of ethylene production [2]. When plants produce ethylene or are exposed to external source of ethylene, receptors perceive ethylene and consequently the signal is transferred, resulted in downstream genes activation. So, ethylene induces expression of related genes involved in senescence of plant organs, leads to changes in physiological characteristics, leaf yellowing and organs abscission [3-4]. The main effects of external and/or internal ethylene on potted roses include leaf yellowing and abscission, acceleration of bud and flower shedding, flower premature aging and decreasing of flower vase life [5]. However, response of potted roses to external ethylene treatment is different dependent on their varieties [4,6-7]. Some anti-ethylene agents such as 1-MCP can suppress ethylene action by occupation of place of ethylene on receptors, resulting in improving postharvest display of rose plants [7,8].
Changing oxidation levels are considered as important events of environmental stresses [9]. Oxygen free radicals could act as marker molecules in commencing and activation of defensive responses of organisms against environmental stress [10]. It was reported that defensive system of enzymatic and non-enzymatic anti-oxidants in plants regulates the rate of oxygen free radicals [11]. Under stress conditions and/or during senescence, oxidative stress is occurred along with the loss of activity of antioxidant enzymes. The plant response to these inferior conditions depends on their ability in regulation of gene expression and function of resistance-related proteins [12]. Peroxidase and copper-containing oxidase enzyme, laccase, play important roles in polymerization of monolignols in plants [13,14]. Both enzymes were found widely in various organisms which play several roles such as lignifications, auxin metabolism and key roles in response to stress and degradation reactions of hydrogen peroxide [15,16]. Peroxidase catalyzes hydrogen peroxide reduction by receiving electron from several donor molecules such as phenols and lignin precursors [17]. Laccase [(EC 1.10.3.2), Parady phenol di-oxygen oxidoreductase] is multi-copper oxidases and known as blue oxidase which while reducing oxygen molecule lead to oxidation a wide range of organic compositions such as phenols, poly phenols and also some mineral compositions by one-electron transportation [18]. Although it was reported that laccase involves in improving wound, absorbing iron and responding to stresses [19,20], it was showed that laccase gene expressed under ethylene treatment in rose [3] and citrus leaves [21].
The aim of this research was to study the physiological response of potted rose cv. Sanaz-e-Zard to short-term exposing to ethylene and evaluation of laccase and peroxidase enzymes activity as well as proline content. According to our knowledge, there is no report relating to expression of laccase enzyme and relationship between laccase and anti-oxidant enzymes. Due to stress condition induced by ethylene, evaluation of anti-oxidant enzymes involved in defensive system could shed light in physiological and bio-chemical behavior of potted miniature rose plants to ethylene.
Plant material and treatment
This research was conducted in a completely randomized design with three replications and three ethylene treatment levels. After harvesting potted miniature rose, they were placed in 200 L glass chambers and fumigated with concentrations of 0, 5 and 10 μL L-1, at laboratory of postharvest physiology, Department of horticultural science, Tarbiat Modares University, potted miniature roses the glass chambers were hermetically sealed for 12 h, and control plants were sealed in ethylene-free air in identical chamber. After 12 h exposing to ethylene, plants were placed in postharvest room under environmental conditions of 20 ± 2°C temperature, relative humidity of 60-65%, 12 h per day of light (15 μmol/m2/s-1). The characteristics of postharvest display including leaf abscission, chlorophyll content, activity changes of enzyme peroxidase and laccase, membrane lipid peroxidation and proline in vegetative organs of potted miniature rose were evaluated during this study. Leaves on secondary branches which were in vegetative phase, no flowering stage were harvested in certain distances. Data analysis was generalized by MSTAT-C statistical software using linear model method (proc glm) where analysis was conducted as plot split experiment in a completely random design. Hormone was regarded as the main factor and sampling time as secondary factor. Comparing averages based on least significant difference (LSD) test was done in five percent probability levels (P<0.05) and figures were drawn using Excel 2007 software.
Evaluation of display quality characteristics
The characteristics of leaf abscission [4] and leaf chlorophyll content was measured based on suggested protocol [22]. Activity change of peroxidase and laccase enzymes was evaluated according to Chance and Maehly [23] and Hoopes and Dean [24]. Lipid peroxidation and leaf proline content was evaluated by the method (Vos et al. and Bates et al. [25,26]).
Chlorophyll content
Measuring chlorophyll was done using suggested method by Lichtenthaler [22] where leaves sample was prepared in every treatment and its repetition once in three days by 8 mm cork borer about 10 samples of disc for. In this method equal number of leaf disc samples were picked up and transported to a 2 mL tube. Then about 1 mL ethanol 80% was added to tubes containing leaf disc and then tubes were stirred. After 10 min, solution containing leaf discs was transferred to 15 mL falcon tubes and reached to equal volume of 15 mL by ethanol 80% and located in warm bath (Ben murray system). By complete discoloring of disc, falcons were taken out of Ben murray. After observing with color of rings and solving their green color in ethanol within falcon, 1 mL of every sample was picked up and read by spectrophotometer system.
Enzyme activity
Peroxidase (POD) enzyme: To extract the peroxidase (POD) enzyme, 200 mg leaf tissue was homogenized in 25 mM Na-phosphate buffer (pH 6.8) followed by centrifugation at 12000 rpm for 30 min at 4°C. For assay, a mixture consisting of 25 mM Na-phosphate buffer (pH 6.1), 28 mM Guaiacol, 5 mM hydrogen peroxide and crude extract was prepared and its absorbance at 470 nm was detected during 1 min using spectrophotometer (BIO-RAD). Enzyme activity was expressed as absorption delta of 470 nm per mg protein [23]. Quantity value of soluble proteins was measured by Bradford method [27].
Laccase enzyme: To extract the laccase enzyme, 500 mg leaf tissue was homogenized in 100 mM sodium acetate buffer (pH 5), followed by centrifugation at 12000 rpm for 10 min at 4°C. The supernatant was used to measure activity of laccase and ABTS 2 mM was selected as precursor. Absorbance at 420 nm was detected during 1 min using spectrophotometer (BIO-RAD). Enzyme activity was expressed as absorption delta of 420 nm per mg protein [24]. Quantity value of soluble proteins was measured according to Bradford [27].
Lipid peroxidation
Damaging membranes was evaluated by measuring rate of MDA as final product of membrane lipids peroxidation. In order to measure MDA, 200 mg leaf tissue was finely powdered in liquid-nitrogen cooled mortar. Then prepared powder was transferred to 15 mL falcon tube and 3 mL Trichloroacetic Acid (TCA) 10% was added. After homogenization, samples were centrifuged at 12000 rpm, for 15 min at 4°C. Then 1 mL of solution was transferred to experiment tubes, added 1 mL Thiobarbituric Acid (TBA) 0.5% and kept in temperature of 100°C for 30 min. MDA was measured by evaluating absorption in 532 and 600 nm wavelengths using constant coefficients [25].
ΔA = ɛ × C × L
ΔA = Absorption delta (digit of spectrophotometer system)
C: Concentration (rate) of material
L: Cuvette width (1 cm)
Proline content
Leaf tissue of 500 mg was powdered in liquid nitrogen and poured in 10 mL of homogenized sulfosalicylic acid 10% and filtered. Acid ninhydrin reagent was prepared by solving 1.25 g of ninhydrin in 30 mL of glacial acetic acid and 20 mL of phosphoric acid 6 M by heating on stirrer. This reagent stays constant in 4ºC for 24 h. About 2 mL of filtered solution reacts to 2 mL of acid ninhydrin and 2 mL of glacial acetic acid in a tube for 1 h within 100°C bath; then this reaction was quenched into a cold bath. Next, 4 mL toluene is added to the solution and integrated using vortex for 15-20 s. The resulted supernatant, being red in color, was read in 520 nm wavelength [26].
Vase life is the main postharvest characteristic of ornamental plants, which is highly affected by ethylene in rose plants. Research on postharvest physiology of rose cut flowers and potted plants resulted in development of basic knowledge in the field of aging and applicable treatments to prevent postharvest losses.
Our results showed that display quality of potted miniature rose plants was significantly affected by ethylene. Increasing exposedethylene concentration enhanced the rate of leaf abscission, as the higher abscission was observed in 10 μL L-1 ethylene which was significantly (P<0.01) higher than control (Figure 1).
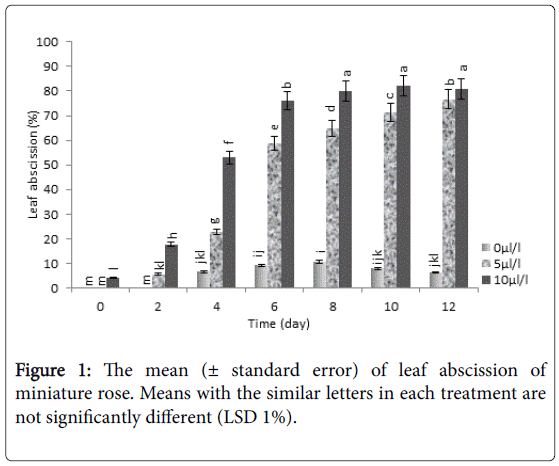
Figure 1: The mean (± standard error) of leaf abscission of miniature rose. Means with the similar letters in each treatment are not significantly different (LSD 1%).
In some plant species, vase life of flower is ended by falling flower pieces with or without senescence [28]. Our observations showed that falling leaf in potted miniature rose is influenced by ethylene and increasing concentration of ethylene gradually raised the percentage of falling leaves. These results are inconsistent with results Ahmadi et al. [4] and Muller et al. [29], on various potted miniature roses. Although some studies showed that ethylene induced higher expression of ethylene receptor genes [30,31], there are controversial reports on expression of receptor genes under ethylene treatment in various organs of miniature rose. Ahmadi et al. [4] reported that expression of ethylene receptor genes (RhETR1/3) and signal transduction genes (RhCTR1/2) was not correlated with ethylene sensitivity of the plant genotypes.
Although ethylene has a regulative act/effect on transcription of and/or translation of expressed genes in cell separation [32], but it is unclear that ethylene is a special inductor of separating cells and causes to develop falling [33]. It is believed that ethylene-dependent or ethylene-independent processes involve falling leaves. As Arabidopsis ethylene sensitive-mutants don't need ethylene to activate process but falling occurs in these mutants; while in mutants by late falling of flower organs fast falling induction was shown in response to ethylene [34].
Our results showed that the lowest chlorophyll content was evaluated at 10 μL L-1 ethylene concentration, which was significantly (P<0.01) lower than control (Figure 2). In potted miniature rose exposed to ethylene, the oxidative stress was induced concomitant with decreasing activity of antioxidant enzymes, premature senescence of leaves, and decreasing significantly chlorophyll content.
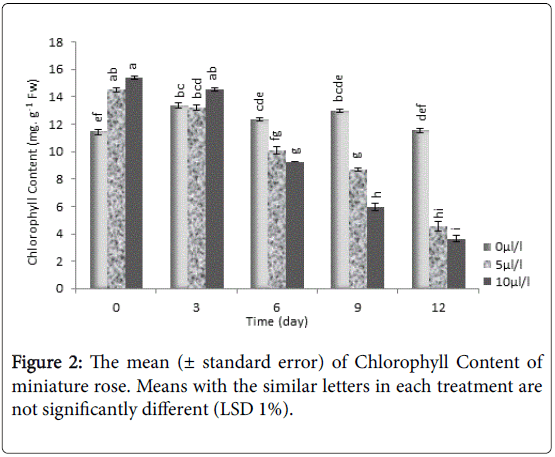
Figure 2: The mean (± standard error) of Chlorophyll Content of miniature rose. Means with the similar letters in each treatment are not significantly different (LSD 1%).
Chloroplast is one of main place of producing ROS in plants [35] which are active in oxidative stress leading to age in cells [36]. Chloroplasts contain many proteases from which some are encoded by senescence-related genes which are regulated during senescence. It seems that ethylene could accelerate the ROS production in cells by increasing the respiration rate or some catabolic reactions. It was reported that ethylene stimulates activity of effective enzymes such as chlorophyllase, Mg-dechelatase and peroxidase which finally lead to degradation of chlorophyll [37]. Moreover, degradation of chlorophyll could be related to decreasing activity of enzymes such as peroxidase, and superoxide dismutase, although this relationship isn't completely known.
According to Figure 3, the highest activity of laccase was characterized in plants treated with 10 μL L-1 ethylene, which was significantly (P<0.01) difference in comparison to other ethylene concentrations and control too. Laccase activity in leaves was increased under exogenous ethylene treatment of 5 and 10 μL L-1, during running experiment; although this activity for 5 μ LL-1 was lower than 10 μL L-1 ethylene.
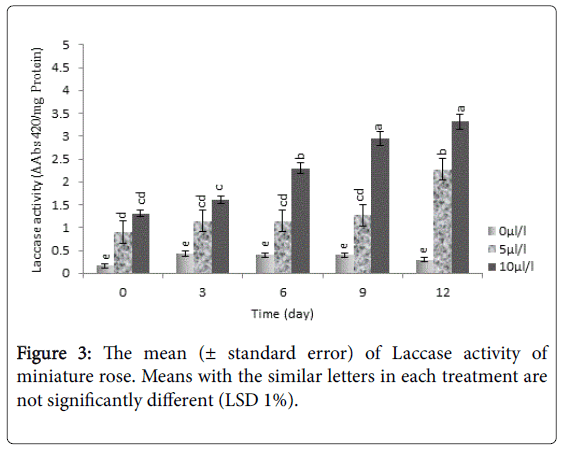
Figure 3: The mean (± standard error) of Laccase activity of miniature rose. Means with the similar letters in each treatment are not significantly different (LSD 1%).
Laccase gene is detected in plants as a multi-gene family [38]. Analyzing of southern blot hybridization showed several copies of the RhLAC gene in the rose species [3], similar to corn laccase that belongs to a multi-gene family [39]. mRNA transcription level of laccase under various stress [20], salinity stress, application of abscisic acid was increased [40]. Application of abscisic acid resulted in accelerating production of ethylene and organ abscission in rose flower [30].
Expressing laccase under treatment of ethylene [3] and other stress [20,40] seems to be related to plant defensive mechanisms in response to pressure conditions [41].
Our results showed that the lowest activity of peroxidase was found in plants treated with 10 μL L-1 ethylene which was significantly (P<0.01) lower than control (Figure 4).
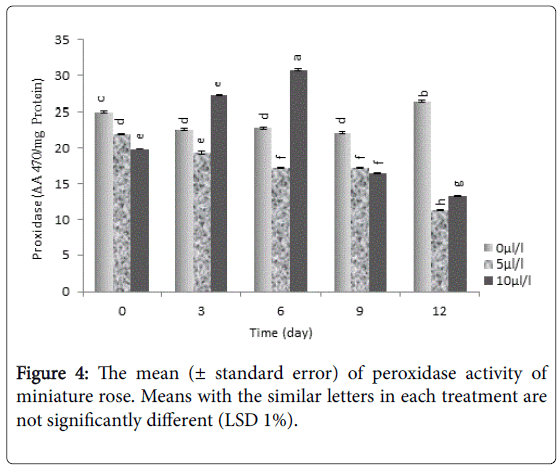
Figure 4: The mean (± standard error) of peroxidase activity of miniature rose. Means with the similar letters in each treatment are not significantly different (LSD 1%).
Activity of antioxidant enzyme and promoting antioxidant capacity of cells are very important factors to control senescence processes [42]. Decreasing activity of antioxidant enzymes in senescence is because of affecting RNA synthesis, which finally causes to decrease protein synthesis and/or inactivation of enzymes [43]. Another agent affecting the activity of antioxidant enzymes during the senescence process could be noted to the increasing of active oxygen species attacking and destroying antioxidant enzymes [44]. Peroxidase enzyme is considered as the most important enzyme reflecting environmental pressures [45], and ethylene could increase the number of reactive oxygen species, lead to the oxidative destruction of cells.
Our evaluation on MDA content showed that the highest amount of MDA was related to 10 μLL-1 ethylene which was significantly (P<0.01) higher than control (Figure 5). This research revealed that rate of inducted lipid peroxidation by ethylene stress increases in plants and increasing concentration of malondialdehyde under ethylene stress probably causes increasing damage of membrane. Increasing peroxidation of membrane lipids in stress conditions showed that plant can't destroy oxygen free radicals and inducing plant's enzymatic defensive mechanisms against oxidative damages. Therefore using anti-ethylene action and/or biosynthesis agents could increase antioxidant capacity of plants against stress.
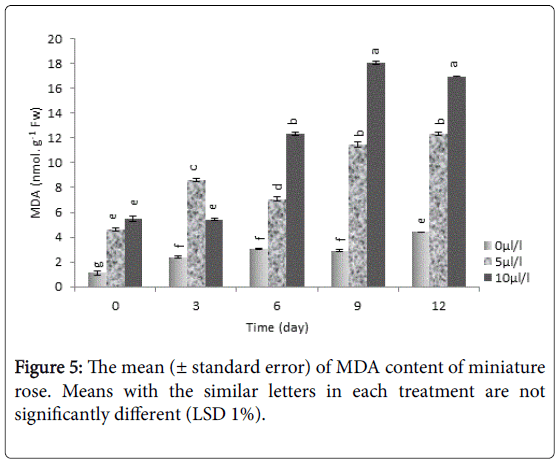
Figure 5: The mean (± standard error) of MDA content of miniature rose. Means with the similar letters in each treatment are not significantly different (LSD 1%).
Similar to our results, increasing of lipid peroxidation during senescence stage in gladiola leaves was reported which leads destroying membrane and reduction of membrane stability [46].
Our results showed that the lowest proline content was related to 10 μL L-1 ethylene which was significantly (P<0.05) lower than control (Figure 6). The amount of proline was decreased during time of running experiment, especially under ethylene treatment. In contrast to our finding, in leaves of Zantedeschia, proline significantly increased during aging time [47]. Data suggests a positive correlation between no proline productions and/or proline oxidation in miniature rose.
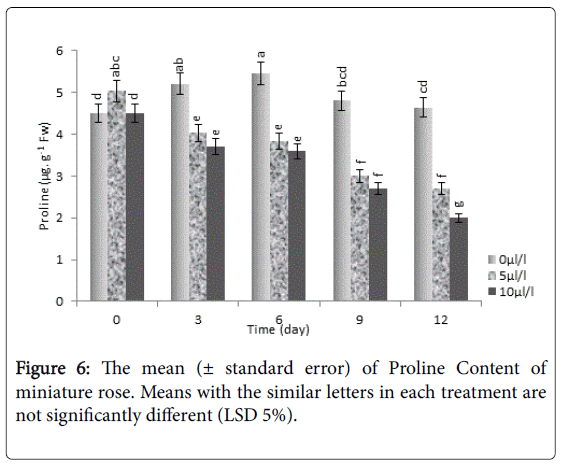
Figure 6: The mean (± standard error) of Proline Content of miniature rose. Means with the similar letters in each treatment are not significantly different (LSD 5%).
Producing and accumulation of proline seems to be an important physiological response to prevent any damage and destroying of protein, enzymes, RNA, DNA and membrane lipids [48]. In senescence stages cell protective ability decreases against oxygen active radicals when cell natural activities and then cell macromolecules such as proteins will be destroyed [49]. Decreasing proline content in plants exposing to exogenous ethylene could be related to the scavenging of reactive oxygen species by proline oxidation under ethylene stress conditions. Moreover, amino acids such as proline often play protective role for chloroplast thylakoids and under pressure membrane systems [50].
Ethylene stress causes to oxidative stress to plant cells by gradual decrease of plant's antioxidant defensive capacity and photosynthesis pigments; decreasing content of proline amino acid leads to variability of cell structure and by increasing peroxidation of membrane lipids causes to accelerate senescence of under-pressure plants.
In conclusion, this research was conducted to evaluate the effects of ethylene on the vase life of miniature rose cv. Sanaz-e-Zard. The results showed that ethylene had significant effect on the physiological and biochemical characteristics. Exogenous ethylene treatment decreased the resistance capacity of tissue/organs by decreasing activity of antioxidant enzymes and increasing peroxidation of membrane lipids. Concomitant increase of laccase activity and leaf abscission could show a relationship between this enzyme and senescence. This is the first report on evaluation of laccase under ethylene and more investigation in this case is required. Moreover, evaluation of isozymes of antioxidant enzymes and changing their activities in order to recognize and study antioxidant system of potted miniature rose and its behavior in ethylene pressure conditions should be considered.