Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2012) Volume 1, Issue 7
Drought is a major abiotic factor that limits agricultural crop production. Antioxidant activity of some plants may be changed by water stress. In this study, a pot experiment was conducted to assess the effect of drought on the antioxidant activity of thyme (Thymus vulgaris L.). Different irrigation regimes (2, 4, 6, 8 and 10 day intervals) were applied for about 6 months and the antioxidant capacity was evaluated by using two different assays; the FRAP (ferric reducing antioxidant power) and DPPH (2,2-diphenylpicrylhydrazyl) scavenging assays. Although in some other species water stress might increase the antioxidant capacity, but the experimental results showed that long-term drought caused
a reduction in the antioxidant activity and total phenolic content of thyme. While all plants died with 10 day irrigation interval, with 2 (control), 4, 6, and 8 days irrigation intervals, IC50 values of the DPPH assay was 87.82, 105.86, 118.05 and 133.60 μg/ml respectively. A higher value of IC50 showed a lower antioxidant activity, which indicated that severe water stress (8 days) significantly decreased the antioxidant activity of thyme (P≤0.05). FRAP values showed a trend to reduction by increasing the irrigation intervals, but this trend was not significant.
Keywords: Antioxidant activity; Thymus vulgaris L; Total phenol content; Water deficit
Plants are subjected to several harsh environmental stresses that adversely affect their growth, metabolism and yield. Drought, salinity, low and high temperatures, flood, pollutants and radiation are important stress factors limiting the productivity of crops [1]. Although the general effects of drought on plant growth are fairly well known, the primary effects of water deficit at the biochemical and the molecular levels are not well understood [2-4]. At the whole-plant level, the effect of stress is usually perceived as a decrease in photosynthesis and growth and is associated with alterations in C and N metabolism. At the molecular level, the negative effects of stress on leaves may be in part a consequence of the oxidative damage to important molecules, as a result of the imbalance between production of activated O2 and antioxidant defenses [5]. It is has been shown that among the aromatic and medicinal plants, the plants which are from Lamiaceae family have marked antioxidant properties [6].
Within Lamiaceae species, examples of new antioxidants include phenolic diterpenes, phenolic carboxylic acids, biphenils and flavonoids isolated from rosemary, sage, oregano and thyme. In many cases, phenolic compounds have antioxidant activities more effective than α-tocopherol, and activity comparable to that of synthetic antioxidants, Bylated Hdroxyanisole (BHA) and Butylated Hydroxytoluene (BHT) [7]. Thyme (Thymus vulgaris L.) like some other wild Thymus species possesses a wide range of biological activities including expectorant, spasmolitic, sedative [8], antifungal [9] and antioxidant properties [10]. In most cases, investigations on biological activities were carried out on extracts, essential oils or pure compounds isolated from dried herbs of Thymus species. Madsen et al. [10] showed that total phenol content in the extracts of different Lamiaceae species (including thyme) linked linearly with the antioxidant activity, as measured by oxygen depletion, but not with the free radical scavenging effects. The results obtained showed that investigated extracts contained components with at least two different antioxidative mechanisms.
Drought generally increased the levels of phenolic compounds in the shoots and roots of Hypericum brasiliense. Therefore, the increase in phenolic compounds seen in H. brasiliense plants stressed by water deficit condition might also induce an antioxidant response to ROS production [11]. It has been shown that water deficiency induces various physiological and metabolic responses like stomatal closure, decline in growth rate, solute and antioxidant accumulation and expression of stress specific genes [12].
According to the previous work, in Pisum sativum L. cv. Lincoln, the increase in catalytic Fe and the decrease of antioxidant protection may be involved in the oxidative damage caused by severe water deficiency [13] .The results also have showed that drought stresses could induce oxidative stress in liquorices (Glycyrrhizin uralensis Fisch) seedlings, as indicated by the increase level of lipid peroxidation [14].
The aim of the present study was to find out the effects of long-term irrigation treatments on the antioxidant activity and total phenolic content of thyme under greenhouse conditions.
Chemicals
In this work, the antioxidant capacity was evaluated by using two different assays, the FRAP (ferric reducing antioxidant power) method and the 2, 2-diphenylpicrylhydrazyl (DPPH) assay.
Acetic acid glacial, ferrous sulfate heptahydrate, ferric chloride, folin-cioacalteu reagent, methanol, sodium acetate, sodium carbonate and 2,4,6-tripyridyl-s-triazine (TPTZ) were purchased from Merck (Darmstadt, http://www.merck.de). Catechin hydrate, 2,2-diphenyl- 1-picrylhydrazyl (DPPH) and hydrochloric acid 32% were obtained from Sigma-Aldrich (St. Louis, http://www.sigma-aldrich.com). Trolox (water soluble equivalent of vitamin E) and quercetin were from Acros Organics (Geel, http://www.acros.com).
Plant material and irrigation treatments
Thymus vulgaris L. transplants were provided from Shiraz Plant Propagator Co. and transferred to greenhouse of College of Agriculture, Shiraz University, Shiraz, Iran, and planted in pots. After 3 months, 5 irrigation treatments, i.e. 2 (control/field capacity), 4, 6, 8 and 10 day intervals were applied for about 6 months. The aerial parts of the studied plants were harvested, and then air dried at room temperature for 2 weeks. The plant samples transferred to the laboratory of Medicinal and Natural Products Chemistry Research Center, Shiraz University of Medical Sciences, Shiraz, Iran, and were tested for antioxidant activities.
Extraction
Methanolic extracts of plants were prepared as follows: 2 g of dried aerial parts of plant (3 samples for each treatment) after being defatted by petroleum ether was macerated in 200 ml methanol/distilled water (90/10) for 2 days, with one change of solvent after 1 day. The extract was filtered and then completely concentrated in a rotary evaporator (EYELA SB-1000 and Heidolf LABOROTA 4000). Extract powders were weighed to calculate the yield and kept at -20°C, until use. Shortly before experiment, the powders were dissolved in methanol at the desired concentration and tested for antioxidant activity and total phenolic content.
Antioxidant capacity determined by Ferric Reducing Antioxidant Power (FRAP)
The FRAP assay was conducted in accordance with Chaitanya et al. [2]. Briefly, the FRAP solution was prepared freshly by mixing 10 ml of acetate buffer 300 mM, pH 3.6, 1 ml of ferric chloride hexahydrate 20 mM dissolved in distilled water and 1 ml of 2,4,6-tripyridyl-s-triazine (TPTZ) 10 mM dissolved in HCl 40 mM. 20 μl of plant extract dissolved in methanol at the concentration of 1 mg/ml was mixed with 180 μl of the FRAP solution in triplicate. Absorbance was determined at 595 nm after 6 min of incubation at room temperature by a microplate reader (Bio-Rad. Model 680). Quercetin was tested at the final concentration of 10 μM and used as the reference compound. FRAP values expressed as μM quercetin equivalent for each gram dry weight of plant (DW) were calculated according to the following formula [15].
FRAP value= (ΔAP/ ΔAQ) × Y × 1000
Where ΔAP and ΔAQ are absorbance changes of the FRAP solution in the presence of the plant extract and quercetin, respectively and Y is the extraction yield. The experiment was performed 3 times and their averages were expressed as the final results.
Antioxidant capacity determined by DPPH assay
The ability to scavenge DPPH free radicals was determined based on the method of previous work [16], with minor modifications. Four different concentrations of the plant extract in methanol (400, 200, 100 and 50 μl) were incubated with a methanolic solution of DPPH 100 μM in glass tubes. Trolox and quercetin at final concentration of 0 μM were used as standard. After 30 min incubation at room temperature in the dark, the absorbance at 517 nm was measured by a spectrophotometer (PERKIN-ELMER, Lambda 1, uv/vis). The percent of inhibition (% I) for each concentration was calculated by using the absorbance (A) values, according to the following formula [15,17].
%I= [(ADPPH – AP)/ ADPPH]
Where ADPPH and AP are the absorbance of the DPPH solution containing methanol and plant extract respectively. The dose-response curve was plotted by using the software CurveExpert for windows version 1.3 and IC50 values for extract were calculated. These values were divided by the extraction yield (Y) to calculate the IC50 value for dry plant. The experiment was performed 3 times and their averages were used as the final results [17].
Determination of total phenolic content
Total phenolic content in plant extract was determined using the Folin-Ciocalteu colorometric method [18]. 200 μl of the plant extract dissolved in methanol (1 mg/ml) was taken into glass tubes, in duplicate. 200 μl of catechin 0.25 mg/ml dissolved in methanol was used in duplicate as standard. Then, 2.5 ml Folin-Cioacalteu reagent (diluted 10 times in deionized water) was added and the samples were incubated at room temperature for 5 min, and were vortex mixed at least 2 times. Then, 2 ml Na2CO3 7.5% dissolved in distilled water was mixed and tubes were closed, were covered with aluminum foil and put on a shaker (speed=60) for 90 min at room temperature. Absorbance of samples were measured by a spectrophptometer at 760 nm blanked against deionized water. Phenol concentration of samples was determined according to this formula [17].
Phenol content (mg catechin/g D.W) = (As/Ac) ×R×250
Where As and Ac are the absorbance of the sample and catechin, respectively, and R is the percent of extraction yield. The experiment was performed 3 times and their averages were expressed as the final results.
Statistical analysis
A completely randomized design with factorial arrangements was used for the experiments. Analysis of variance was carried out using SAS software. Means were separated using Tukey’s test at P=0.05. Regression analyses were performed by Sigmaplot 2002 for Windows version 8.0.
In this study, FRAP value (Table 1) and total phenolic contents (Table 2) of samples were decreased with increase in the duration of irrigation intervals, but the differences were not significant. Control showed higher values and 8 days interval had lower values. The results of the DPPH assay showed that with water deficit conditions, IC50 of samples were increased significantly (Table 1). IC50 values were increased from control to severe stress condition (8 days interval). In this study, regression analysis showed a correlation between total phenolic contents with two antioxidant assays (Figure 1).
| Treatment intervals (Days) | FRAP value (Days) | IC50 DPPH scavenging(μM/g DW)* (μg DW/ml)** |
|---|---|---|
| 2 (control) | 1617.40a | 87.82a |
| 4 | 1313.27a | 105.86ab |
| 6 | 1343.68a | 118.05ab |
| 8 | 1274.28a | 133.60b |
Table 1: FRAP and DPPH activities in Thymus vulgaris watered with different irrigation intervals.
| Treatment interval (days) | Total phenolic content (mg Catechin/g DW)* |
|---|---|
| 2 (control) | 44.10a |
| 4 | 38.37a |
| 6 | 36.90a |
| 8 | 33.10a |
Table 2: Total phenolic contents of Thymus vulgaris watered with different irrigation intervals.
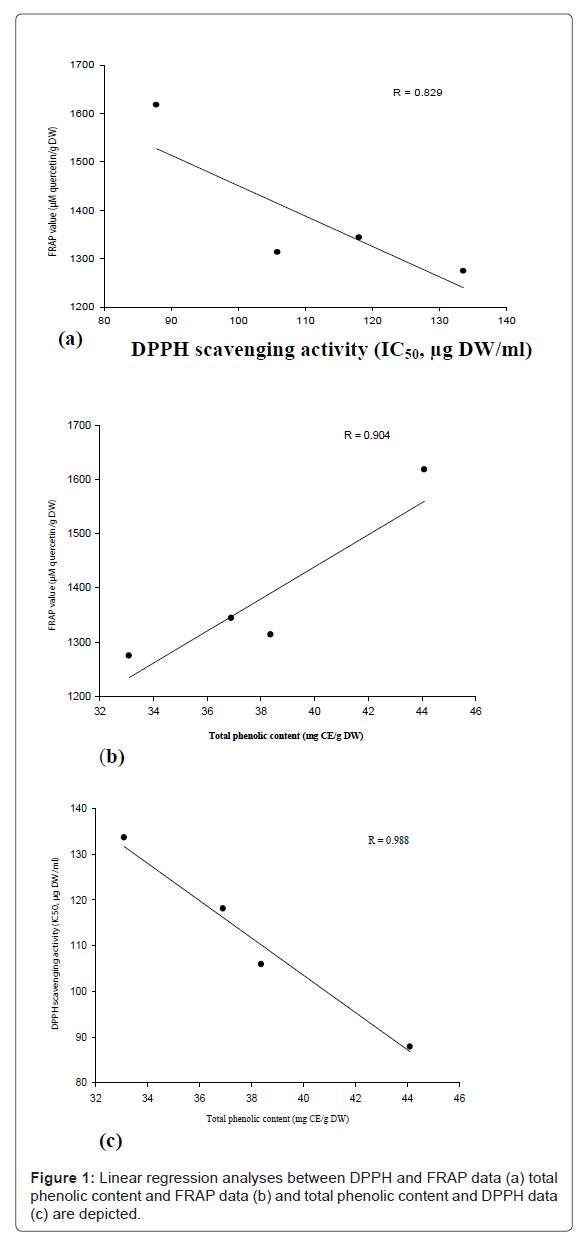
Figure 1: Linear regression analyses between DPPH and FRAP data (a) total phenolic content and FRAP data (b) and total phenolic content and DPPH data (c) are depicted.
It seems that antioxidant defense system is impaired under long-term water deficit conditions. The results of the present study confirmed the data of previous reports on Pisum sativum L. [13,19], Glycyrrhiza uralensis Fisch [14], Sorghum bicolor L. and Helianthus annuus L. under severe drought conditions [20] and Salvia officinalis L. [21], which showed a decline in the antioxidant capacity under drought. However, the data obtained in this investigation were not in agreement with the results on H. brasiliense Choisy [11] and Catharanthus roseus L. [22]. It can be concluded that water deficiency might increase antioxidants levels depending on plant genotypes; however, severe or long-term water deficiency decreased antioxidants activity. Statistical analysis showed a significant linear correlation between the antioxidant activity and the total content of polyphenols in various parts of Allium species, as appraised by the FRAP test both with the DPPH test [4]. Madsen et al. [10] found that total phenol content in the extracts of different Lamiaceae species (including thyme) correlated linearly with the antioxidant activity, as measured by oxygen depletion, but not with the free radical scavenging effects.
Authors wish to thank the cooperation of the personnel of the Laboratory of Medicinal and Natural Products Chemistry Research Center, Shiraz University of Medical Sciences, Shiraz, Iran.