Virology & Mycology
Open Access
ISSN: 2161-0517
ISSN: 2161-0517
Research Article - (2017) Volume 6, Issue 2
Studies were conducted in vitro to determine the antagonistic potential of Trichoderma harzianum for biological control of Fusarium moniliforme isolated from rotted Dioscorea rotundata tubers. The experiments were performed at Advanced Plant Pathology Laboratory, Federal University of Agriculture, Makurdi, Nigeria. Fungi organisms isolated and identified from the rotted white yam tubers were Botryodiploidia theobromae, Aspergillus flavus, Aspergillus niger, Aspergillus ochraceus, Fusarium moniliforme, Fusarium oxysporum, Penicillium purpurogenum and Pestalotia sp. Pathogenicity tests done confirmed F. moniliforme as one of the organisms responsible for the rot in yam tubers in this location. The antagonist was introduced same time with the pathogen, two days before the inoculation of the pathogen and two days after the inoculation of the pathogen. Plates were incubated for 192 hours and measurements of mycelial radial growths were done at intervals of 24 hours beginning from the third day of inoculation. The results of in vitro dual culture interactions between T. harzianum and F. moniliforme showed that T. harzianum has potentials to significantly (P ≤ 0.05) inhibit the growth of F. moniliforme irrespective of the time of introduction of antagonist and duration of incubation. Mean Percentage growth inhibition was highest (58.70%) when the antagonist was introduced 2 days before inoculation of the pathogen, followed by (52.54%) introduction of the antagonist same time with the pathogen while the least percentage growth inhibition (34.33%) was recorded when T. harzianum was introduced 2 days after inoculation of F. moniliforme. Minimum inhibition concentration showed moderately effective to effective control depending on the time of introduction of the antagonist. In conclusion, biological control agents should be used since they are biodegradable, eco-friendly, less expensive and target specific. The introduction is done before the arrival of pathogenic organisms in order to achieve highest level of effectiveness in controlling post-harvest rots causing organisms.
<Keywords: In vitro; Antagonist; Biological control; Fusarium moniliforme; Dioscorea rotundata ; Pathogenicity test; Inhibition
Yams (Dioscorea spp.) are tuber crop belonging to the family of Dioscoreaceae [1]. Yams are widely cultivated in West and Central Africa, in Asia and South American countries [2]. Yam is the most important tuber crop cultivated in West Africa in terms of area coverage and a key staple food, particularly in Nigeria [3]. More than 90% of the global yam production constituting about 38.92 million metric tonnes fresh tubers are produced in West Africa annually [4,5].
Yam tubers have very high food value and are major sources of carbohydrate, minerals such as calcium, phosphorus, iron and vitamins such as riboflavin, thiamine and vitamins B and C [6,7]. In as much as yam is produced on a high scale in the country, it is nonetheless affected by pathogenic organisms which is responsible for causing rot in yam tubers. Yams rots usually start in the soil and progress in storage; this may occur when the infected tubers do not yet have any sign of external symptoms [6].
Fungi causing rot are of particular importance because they reduce vigour and subsequently cause a reduction in tuber yield and quality [8]. In yams, a record of about 50% reduction of the total stored tubers has been reported within the first 6 months of storage [9]. Rot of yam tubers may be caused by a wide range of pathogenic organisms including fungi, bacteria and viruses at all stages of growth and also during storage of tubers [8,10].
These pathogenic fungi include Aspergillus flavus , Aspergillus niger, Botryodiplodia theobromae, Colletotrichum spp, Fusarium oxysporum, Fusarium solani, Fusarium moniliformes Geotrichum candidum, Penicillium chrysogenum, Pennicillium digitatum, Rhizoctonia sp, Penicillium oxalicum, Rhizopus nodosus [11-15]. A total of 30 different fungi have been reported to be associated with the storage rots of yam [16,17].
The use of synthetic chemicals such as borax, captan, thiobendazole, benomyl, bleach (sodium hypochlorite) has been found to significantly reduce storage rot in yam but chemicals have been proven to have detrimental effects on the environment as they are non-biodegradable and extremely toxic [18-20]. Plant extracts have equally been used to control yam diseases in different parts of Nigeria [14,21,22].
The use of biological control method using Trichoderma viride , T. harzianum , Pseudomonas syringae , Pseudomonas chlororaphis , Gliocladium roseum , and Bacillus subtilis is generally favoured as a method of plant disease management [7,23-25]. Biological control method has been preferred in some cases because it is selective with no side effect and is cheap.
Resistance to biological control is rare and biological control agents are self-propagating and self-perpetuating [23,26]. Biological control agents have no phytotoxic effects, do not create any pollution problems as it is eco-friendly, promote plant growth, solubilize phosphorus and micronutrients and hence increase soil fertility, compete aggressively with other microorganisms, suppressive soil borne pathogens and are target specific with no adverse effect on the host [27-31].
The antagonists also produce antibiotics or toxin at low concentration which poison or kill other microorganisms, it also inhibits pathogens by secreting enzymes like glucanase, cellulase, chitinase protease etc which disintegrate the cell wall of pathogens [30,32].
It is therefore, necessary to search for a method of control of yam rot fungi organisms that will be affordable, durable, target specific and more importantly free from pollution of the ecosystem. In view of this, the in vitro inhibitory potential of Trichoderma harzianum introduced at different times with Fusarium moniliforme isolated from rotted white yam tubers was carried out in the present study.
Study Area
The research was carried out at the Advanced Plant Pathology Laboratory, Federal University of Agriculture, Makurdi, Nigeria.
Source of T. harzianum isolate
T. harzianum was obtained from yam Pathology Unit of University of Ibadan, Oyo State, Nigeria. Stock cultures of the isolate were maintained on slants of acidified potato dextrose agar (PDA) in McCartney bottles and stored for subsequent use.
Source of rotted and healthy yam tubers
Rotted yam tubers of Pepa and Hembankwase white yam varieties (Dioscorea rotundata) showing varying degrees of dry rot symptoms were obtained from yam farmers from various storage barns in Tor- Donga, Benue State, Nigeria.
The tubers were carefully and neatly packaged in sterile polyethylene bags to avoid wounding, tubers were taken to the laboratory for isolation and identification of pathogens two days after collection. Potato Dextrose Agar (PDA) was the medium used for isolation. The healthy yam tubers collected were used for pathogenicity test. Test fungus in this study was F. moniliforme.
Isolation and identification of fungi pathogens
Diseased yam tissues were washed under running tap water to remove surface soil, debris and other contaminants. Tissue pieces were cut out from the leading edge of lesion, approximately 2 × 2 mm with sterile scalpel and placed in 5% sodium hypochlorite for 2 minutes, then washed in four successive changes of sterile distilled water and dried on sterile filter paper [14].
The dried infected tissues were aseptically plated on Petri dishes containing acidified sterile potato dextrose agar (PDA) and the plates were incubated at ambient room temperature (30 ± 5°C) for 7 days. Fungi organisms were identified following sub-culture of growing mycelia after 7 days of incubation when pure cultures were obtained [14]. Microscopic examination and morphological characteristics were noted and compared with already established authorities [33,34].
Determination of frequency of occurrence of isolates
To determine how often the isolates occur, records of organisms isolated were kept on periodic basis. Since isolation and characterization were carried out at monthly interval, the number of times each fungi pathogen was isolated in a month was expressed as a percentage of the total of all the different organisms over the period, which was calculated as follows [26];
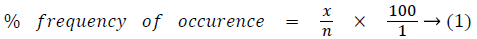
where,
x=number of times of occurrence of the individual isolates over the period
n=total number of micro-organisms isolated in the study over the period
Stock cultures of the isolates were maintained on slant of acidified potato dextrose agar (PDA) in McCartney bottles for subsequent studies.
Pathogenicity test
Pathogenicity test was done to confirm the infectivity of F. moniliforme from the healthy looking white yam tubers collected in the study area. Culture of F. moniliforme was used to inoculate yam tubers. A five-millimeter diameter cork borer was used to remove 4 mm tissue from the healthy D. rotundata tuber surfaces aseptically [35].
A 5 mm diameter mycelial agar plug of a 5 day-old culture of the fungus was used for inoculation. These fungal plugs were put in the holes created in the yam tubers. Petroleum jelly was used to seal the edges of the replaced yam tissues [36].
The same procedure was used for the control experiments except that discs of uninoculated PDA were placed in the holes created in the tubers [37]. The inoculated yam tubers were placed at room temperature in a completely randomized design for 14 days under sterile condition to determine rot [38].
On appearance of rot symptoms, a sterilized and flamed knife was used to cut open the inoculated yam tubers from the point of inoculation. The tissues at the margins of the healthy and diseased parts were surfaced-sterilized, excised and plated onto PDA for incubation at ambient room temperature (30 ± 5°C) for 7 days. At the end of this period, morphological characteristics and growth patterns observed were compared with the one initially obtained.
Evaluation using dual culture on agar plates
The assay for antagonism was performed on Potato Dextrose Agar (PDA) on Petri dishes by the dual culture method [39]. 5 mm diameter mycelial plugs of T. harzianum and F. moniliforme were both inoculated on the same dish with the antagonist (T. harzianum) and pathogen (F. moniliforme) 6 cm from each other. The antagonist was plated same time with the pathogen, two days before the inoculation of pathogen and two days after inoculation of pathogen on Petri dishes containing 15 ml of sterile PDA.
Paired cultures were incubated at room temperature (30 ± 50°C) for 192 hours. In the control plates, PDA plugs were used against the pathogen instead of the antagonist. Treatments comprised times of introduction of antagonist which were replicated three times for each treatment and arranged in completely randomized design [38].
Measurement of radial mycelia growth
Measurement of radial mycelia growths of the antagonist and the fungus in dual culture and the fungus in control plates were done after three days of inoculation on a 24 hour interval beginning from the 72nd hour up to the 192nd hour of incubation at ambient room temperature (30 ± 50°C) [25]. Percentage Growth Inhibition (PGI) of F. moniliforme was calculated based on the method of Korsten and De Jager [40].
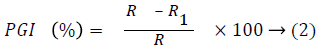
where,
PGI=Percent Growth Inhibition
R=the distance (measured in mm) from the point of inoculation to the colony margin in control plate,
R1=the distance of fungal growth from the point of inoculation to the colony margin in treated plate in the direction of the antagonist.
The percent growth inhibition was determined as a guide in selecting the minimum inhibition concentration (MIC) that will be effective in controlling the rot-causing fungus for the three treatments. Antagonist was also rated for inhibitory effects using a scale by Sangoyomi [41] as:
≤ 0% inhibition (not effective),
>0-20% inhibition (slightly effective)
>20-50% inhibition (moderately effective),
>50-<100% inhibition (effective)
100% inhibition (highly effective)
Data analysis
Data collected were subjected to Analysis of variance (ANOVA) using GenStat Discovery Edition 12 for ANOVA and means separation, Minitab Release 17 for descriptive statistics and Graph Pad Prism 6 for trend graphs. Statistical F-tests were evaluated at P ≤ 0.05. Differences among treatment means for each measured parameter were separated using Fisher’s least significant difference (FLSD) [42].
Isolation and identification of F. moniliforme
Colony of F. moniliforme on PDA grew rapidly with white aerial mycelium (Figure 1). Tan to orange spore masses of irregular shape and size were occasionally present. Abundant microconidia were formed and were hyaline, usually one-celled but occasionally twocelled, oval to club-shaped, and slightly flattened at each end (Figure 2).

Figure 1: Culture of F. moniliforme on PDA.

Figure 2: Micrograph showing abundant formation of microconidia (10X).
Macroconidia were formed infrequently and they were hyaline, delicate with thin walls, curved to almost straight, 3-7 septate, and had a foot-shaped basal cell. Chlamydospores were never present in the mycelium or conidia. Perithecia were not observed.
Percentage frequency of occurrence of fungal isolates in Tor- Donga
Eight fungi organisms were isolated and identified from Pepa and Hembankwase cultivars of rotten white yam tuber samples collected from farmers’ barns in Tor-Donga which is one of the largest yam markets in Nigeria. The fungi organisms identified in this location were Botryodiploidia theobromae , Aspergillus flavus , Aspergillus niger , Aspergillus ochraceus , Fusarium moniliforme , Fusarium oxysporum , Penicillium purpurogenum and Pestalotia sp.
Figure 3 shows the percentage frequency of occurrence of the isolated fungi organisms in this location. The result revealed that A. ochraceus was higher in Hembankwase compared with Pepa in February but increased in Pepa above Hembankwase in March, April and May. The occurrence of Pestalotia sp was higher in Pepa in February and March but less in subsequent periods under observation.

Figure 3: Percentage frequency of occurrence of fungal isolates from Pepa and Hembankwase white yam cultivars from February to May 2015 in Tor-Donga.
The occurrence of A. niger and A. flavus were similar in the two cultivars of white yam; the two fungi recorded the highest occurrence in Hembankwase in February and march while Pepa recorded the highest percentage occurrence in April and May. The occurrence of P. purpurogenum and F. oxysporum were less in Hembankwase but high in Pepa in the first two months while in the last two months the frequencies of the two fungi organisms were more in Hembankwase than in Pepa . B. Theobromae was observed to occur higher in Pepa compared with Hembankwase throughout the period of isolation.
F. moniliforme occurred higher in Hembankwase in February and April but was less in March and May while Pepa recorded the lowest percentage occurrence of the fungi organisms in February and April but highest occurrence of F. moniliforme in March and May. Variation of mean percentage frequency of occurrence of each fungus by the respective cultivars in this location after four months of isolation showed no significant difference (P ≤ 0.05) between the cultivars (Table 1).
| Pathogens | White Yam Cultivar | T-Value | P-Value | |
|---|---|---|---|---|
| Pepa | Hembankwase | |||
| B. Theobromae | 17.68±0.56 | 15.09±1.25 | -1.88 | 0.13 |
| A. flavus | 15.51±1.28 | 17.78±0.79 | 1.51 | 0.2 |
| A. niger | 24.51±0.19 | 24.08±1.26 | -0.34 | 0.75 |
| A. ochraceus | 7.29±0.40 | 6.86±0.61 | -0.59 | 0.58 |
| F. moniliforme | 11.65±1.10 | 13.07±0.58 | 1.15 | 0.31 |
| F. oxysporum | 16.14±0.95 | 16.07±1.43 | -0.05 | 0.96 |
| P. purpurogenum | 7.20±0.80 | 7.03±0.53 | -0.17 | 0.86 |
| Pestalotia sp. | 6.52±0.36 | 7.86±0.87 | -0.25 | 0.21 |
Table 1: Variation of mean percentage frequency of occurrence of fungal isolates from Pepa and Hembankwase cultivars of white yam tubers after four months of isolation in Tor-Donga
Pathogenicity test
Pathogenicity test shows that F. moniliforme was able to initiate rot in the healthy looking yam tubers after 14 days of inoculation. Symptoms of infections were seen on the inoculated yam tubers when the tubers were transversely cut open (Figure 4).

Figure 4: Rot caused by F. moniliforme .
The yam tubers that were not inoculated with the mycelia of the test fungus (control experiment) however, did not show any sign of rot indicating absence of reproductive propagules of the pathogen in the bored yam tissues (Figure 5).

Figure 5: Control (no organism inoculated).
Evaluation using dual culture method on agar plates
Results of dual culture method evaluated showed that T. harzianum inhibited the growth of F. moniliforme at different rates depending on the time of introduction of the antagonist. The dual culture interactions showed significant success in biocontrol of the fungus under in vitro conditions. Result of the study revealed that T. harzianum exhibited antagonistic activities on F. moniliforme in all the treatments.
The potential antagonist was seen to grow faster than the fungus in the dual culture. The control plates also grew much faster than their respective dual cultures. In all the treatments, the antagonist continued its growth uninhibited, over grew the pathogen resulting into complete degradation of the fungus and sporulation of the antagonist over the entire plate. There were no zones of inhibitions formed in these interactions.
Mycelia inhibition were however, observed higher when the antagonist was introduced 2 days before inoculation of pathogen (Figure 6) followed by introduction of the antagonist same time with the pathogen (Figure 7) and least when the antagonist was introduced 2 days after inoculation of the pathogen (Figure 8). The results revealed the capability of T. harzianum to reduce mycelia growth of all the tested pathogens when it was grown in dual culture irrespective of the time of introduction of the antagonist and duration of incubation.

Figure 6: Dual culture of T. harzianum and F. moniliforme on potato dextrose agar (left); T. harzianum was introduced 2 days before inoculation of F. moniliforme (2 dbi) and pure culture of F. moniliforme on potato dextrose agar (right).

Figure 7: Dual culture of T. harzianum and F. moniliforme on potato dextrose agar inoculated same time (Th×path) (left) and pure culture of F. moniliforme on potato dextrose agar as control (right).

Figure 8: Dual culture of T. harzianum and F. moniliforme on potato dextrose agar (left); T. harzianum was introduced 2 days after inoculation of F. moniliforme (2 dai) and pure culture of F. moniliforme on potato dextrose agar as control (right).
Table 2 shows the percentage growth inhibition of F. moniliforme increasing with increase in time of incubation from 72 hours to 192 hours irrespective of the treatment. When T. harzianum was introduced same time with F. moniliforme , it was observed that the percentage growth inhibition of the fungus in dual culture with T. harzianum increased steadily from 17.26% at 72 hours to 75.49% at 192 hours respectively. A similar trend was recorded when T. harzianum was introduced two days before inoculation of the pathogenic fungus, with percentage growth inhibition of 33.30% at 72 hours to 81.83% at 192 hours respectively. Percentage growth inhibitions rose from 19.18% at 72 hours to 51.75% at 192 hours when T. harzianum was introduced two days after inoculation of F. moniliforme Duration of incubation for each of the treatments showed significant differences at 24 hour interval. Mean variation of percentage growth inhibition of F. moniliforme tested at different times of introduction of T. harzianum significantly (P ≤ 0.05) inhibited the growth of the fungus (Table 2).
| Duration of Incubation (Hours) | Time of Introduction of T. harzianum | ||
|---|---|---|---|
| Th × Path | Th2dbiPath | Th2daiPath | |
| 72 | 17.26±3.90e | 33.30±16.70c | 19.18±9.54c |
| 96 | 31.30±4.29d | 33.73±5.16c | 41.48±4.95abc |
| 120 | 53.44±2.51c | 56.06±4.01bc | 23.40±10.40bc |
| 144 | 65.93±2.34b | 70.20±1.93ab | 36.43±8.28abc |
| 168 | 71.78±2.59ab | 77.06±1.51ab | 43.71±5.24ab |
| 192 | 75.49±2.04a | 81.83±1.17a | 51.75±3.44a |
| LSD | 9.43 | 22.78 | 22.94 |
| Mean (LSD=13.83) | 52.54±5.32a | 58.70±5.36a | 34.33±3.93b |
Table 2: In vitro Percentage Growth Inhibitions (PGI) of F. moniliforme at different times of introduction of T. harzianum . [Means on the same column with the same superscript are not statistically significant (P ≤ 0.05). Means on the same row (for Mean) with the same superscript are not statistically significant (P ≤ 0.05) by time of introduction of T. harzianum . Th × path=T. harzianum introduced same time with pathogen; Th2dbi path=T. harzianum introduced 2 days before inoculation of pathogen; Th2dai path=T. harzianum introduced 2 days after inoculation of pathogen].
Radial mycelia growth and determination of inhibition
The mycelia growth of both the dual culture and control plates were measured at intervals of 24 hours beginning from the 72nd hour of incubation. It was observed that T. harzianum grew faster than F. moniliform in the dual culture plates. The control plates also grew faster than their respective dual cultures (Figure 9). Minimum inhibition concentration (MIC) shows that T. harzianum was moderately effective to effective in controlling F. moniliforme in culture and varies significantly (P ≤ 0.05) across treatments (Table 3).

Figure 9: Radial growth of F. moniliforme in dual culture with T. harzianum after 192 hours of incubation. Th × path=T. harzianum introduced same time with pathogen; Th2dbi path=T. harzianum introduced 2 days before inoculation of pathogen; Th2dai path=T. harzianum introduced 2 days after inoculation of pathogen.
| Time of Introduction of T. harzianum | Percentage Growth Inhibition (PGI) | MIC (%) | Level of Effectiveness |
|---|---|---|---|
| ThXPath | 52.54a | >50-<100 | Effective |
| Th2dbiPath | 58.70a | >50-<100 | Effective |
| Th2daiPath | 34.33b | >20-50 | Moderately effective |
| LSD | 13.83 |
Table 3: Mean Percentage Growth Inhibition of F. moniliforme in dual culture with T. harzianum showing Minimum Inhibition Concentration (MIC). [Th × path=T. harzianum introduced same time with pathogen; Th2dbi path=T. harzianum introduced 2 days before inoculation of pathogen; Th2dai path=T. harzianum introduced 2 days after inoculation of pathogen; MIC=minimum inhibition concentration (%); ≤ 0% inhibition (not effective); >0-20% inhibition (slightly effective); >20-50% inhibition (moderately effective); >50-<100% inhibition (effective); 100% inhibition (highly effective)].
Fungal pathogenic organisms have been found to be previously associated with tuber rots of yam in different locations in Nigeria. F. moniliforme is one of the fungi pathogenic organisms causing deterioration in yam tubers [12,43]. Result of the finding revealed that A. niger , A. flavus , B. Theobromae , F. oxysporum and F. moniliforme are the most frequently occurring fungi pathogenic organisms in both Pepa and Hembankwase yam tuber cultivars. This result is similar to the findings of Ogunleye and Ayansola, who found these pathogenic organisms to be the most devastating rot causing organisms in yam tubers in different locations in Nigeria [12,15,44]. Result of the pathogenicity test shows that the mycelia of F. moniliforme were able to initiate rot in the healthy looking yam tubers. The pathogen enters and established itself in the tissues of yam and was able to utilize the nutrients of yam for growth and development. The control tubers that were not inoculated with the mycelia of F. moniliforme did not in any way show any symptom of rot indicating absence of infectious propagules in the bored yam tissues.
The findings showed that T. harzianum has inhibitory effects on the mycelia growth of F. moniliforme in all the treatments in the dual culture. The result also revealed that the mechanism of action by T. harzianum was by competition with F. moniliforme for nutrients and space, as well as mycoparasitism over the pathogen and probably secretion of antibiosis. The rapid growth and competition for nutrient and space by the antagonist inhibited the growth of the pathogen. The pathogen presented a defined edge of growth opposite to the antagonist, with lysis occurring in the mycelial fragments in the contact zone. T. harzianum must have acted by possible production of chitinolytic and/or glucanases enzymes in bio-control by Trichoderma [30]. This enzymes function by breaking down the polysaccharides, chitin, and glucans that are responsible for the rigidity of F. moniliforme cell walls, thereby destroying the cell wall integrity and limiting the growth of F. moniliforme . In all the treatments, the antagonist overgrew the pathogen resulting to starvation and subsequently death of the pathogen [30,45].
The use of Trichoderma species have been successfully employed as biological control agents due to their high reproductive capacity, efficient utilization of nutrients, strong aggressiveness against other pathogens as well as rapid and effective colonization of wound sites against the invading pathogens [46-48]. The fast growth rate of this antagonist is similar to the work of Ekefan et al. who suppressed the growth of Colletotrichum capsici with T. harzianum isolates eventually overgrowing it within seven days of incubation [49]. Dual culture combinations of T. harzianum with F. moniliforme showed the inhibition of the pathogen in all the treatments with the highest percentage growth inhibition in the treatment where the antagonist was introduced 2 days before inoculation of the pathogen. According to Campbell, Okigbo, there are no bio control agents that have enough competitive ability to displace an already established pathogen [18,50].
The time lapse between inoculation of T. harzianum and F. moniliforme contributed to the success recorded with the antagonist against the pathogenic fungus. Minimum inhibition concentration showed that the antagonist T. harzianum was more effective when it was introduced 2 days before inoculation of F. moniliforme ; this is due to its high competitiveness for nutrient and space, mycoparasitism as well as production of antibiosis which is responsible for degradation of fungus cell wall. Since T. harzianum has fast growth rate and can easily colonize site, it is recommended that application of the antagonist on yam setts, leaves of growing plant as well as yam tubers in storage can greatly inhibit fungal growth and development and hence reduce preharvest and postharvest losses of the crop.
The finding has revealed that T. harzianum has the capacity of inhibiting mycelia growth of F. moniliforme isolated from rotted yam tubers. The result showed that the best time of introducing this antagonist in order to achieve highest level of effectiveness in controlling post-harvest yam tuber rot fungal pathogens is to introduce it some days before the arrival of the fungus. Biological control agents should therefore be employed as alternative ways of reducing rot in yams compared with synthetic fungicides since they are biodegradable, eco-friendly, less expensive and target specific.
The authors declare that there is no conflict of interest regarding the publication of this paper.