Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2012) Volume 1, Issue 7
Keywords: Aframomum danielli; Heligmosomodes bakeri; Mus musculus; Plant extracts
Parasitic infections are today one of the world’s most devastating causes of death in livestock and humans. This occurs especially in the developing countries where climatic factors (temperature, rainfall and humidity), hygiene, poverty and demographic conditions favour the development of parasites, their infections and the transmission of their diseases [1]. Intestinal helminthiasis is an important public health problem in the tropical and subtropical zones, the study which has recently been neglected in favor of HIV/AIDS, Malaria and Tuberculosis. Small ruminants’ (sheep and goats) improve life conditions of populations in the rural farming environment where poverty is of great concern. They actively contribute in the improvement of social cohesion (weddings and traditional customs) of the farming household procure some revenue and allow the education of children. In traditional livestock systems, these animals are affected by disease among which gastrointestinal nematode parasitism is highly prevalent and creates a huge economic loss. This loss results concretely from the mortality of young animals and the decrease in production [2-4]. In developed countries, gastrointestinal parasites are controlled by the use of commercial anthelminthics whereas in the developing countries, traditional methods of helminth control remain largely dependent on medicinal plants. The misuse and poor formulation of synthetic drugs for many decades have led to the development of anthelminthic resistant (AR) worm strains or populations [5,6]. Also some side effects are noted on the animals. The use of disinfectants to control free living stages of the parasites is harmful to the environment [7]. Therefore, new approaches to nematode parasite control are needed to counteract problems of the cost, availability of synthetic drugs and AR [5,8]. This study aims at evaluating ovicidal and larvicidal properties of extracts of Aframomum danielli commonly calls “Nsoak” in the Kenyang language and used traditionally by the people of Sabis in the Upper Banyang Subdivision, Manyu Division Southwest Region, Cameroon as a worm medicine. The traditional treatment is based on eating the seeds and swallowing the juice of the ripened fruit each time you have stomach ache. Heligmosomoides bakeri used in this experimental study is a useful organism and has been used for testing anthelminthics.
Aframomum danielli, Dalziel 1984 belongs to the family Zingiberaceae. It is a perennial succulent plant and has tuberous rhizomes, with tuber bearing roots, a short stem replaced by a pseudostem form from leaf sheaths. Leaves are simple, those towards the base of the plant are usually bladeless and reduced to sheath, flowers are highly coloured with yellow stamens. Species identification was carried out at the National Herbarium Cameroon (NHC) Yaounde. Seeds of A. danielli collected along Taiyor–Sabes secondary forest (in Upper Banyang Subdivision, Manyu Division Southwest Region Cameroon,) were spread on polyethene bags for four hours every day for a week, and then ground using a grinding machine. The powder was sieved and preserved in air tight plastic bags at a temperature of 24°C and relative humidity (RH) of about 67% for subsequent use in the laboratory.
Preparation of extracts
Three types of extracts were prepared to compare their effects and to increase the chances of detecting their ovicidal and larvicidal properties. These extracts were cold water extract (CWE), hot water extract (HWE) and ethanolic extract (EtE).
Ethanolic extracts
One hundred grams of ground seed powder of A. danielli were macerated in 1.5 litre of ethanol (95%), which removed more polar compounds. The mixture was stirred daily at the same time to permit a better extraction of the active ingredients of the plant, and 72hrs (3 days) later, the ethanolic extract was obtained [9]. This was followed by the dilution of 0.2g of the concentrated extract (using a rotary evaporator) first with 3% tween, a diluent which helped to dissolve the organic extracts. After 3-8 minutes, distilled water (DW) was added to obtain 20 millilitres. This became known as the standard solution from which series of dilutions were made to obtain solutions with different concentrations (625, 1250, 2500, 3750 and 5000 μg/ml)
Cold and hot water extracts
One hundred grams of seed powder of A. danielli were macerated in 1.5 litre of DW. The mixture was stirred daily for 48 hrs (to avoid growth of fungi) and after cold water extract was obtained. The solution was first filtered using a tea sieve and then a filter paper (pore diameter 2.5 μm) placed inside a glass funnel. 200 ml of filtrate was distributed inside four beakers, which were then placed inside an oven heated at 50°C for seven days allowing the extract to get dry. 0.2g of the dried extract obtained was diluted with 20ml of distilled water to get a standard solution from where after a series of dilutions, five different concentrations like those of the organic extract were obtained.
For the hot water extract (HWE), a similar procedure to that of CWE was applied except for the fact that the DW was heated at 100°C and 100g of the seed powder poured into hot distilled water. The mixture was allowed to stand at a temperature of 24°C for 3 hrs and the rest of the protocol is the same as that of CWE.
Recovery of eggs and larvae
Freshly passed out feces of artificially infected mice (Mus musculus) was collected using a tea spoon.1g of feces was homogenized in a mortar using a pestle with 60 ml of saturated NaCl solution. This solution was successively filtered first using a tea sieve and later a 150 μm sieve. The filtrates were transferred into two test tubes and filled until the formation of an upper miniscus. A cover slip was used to cover each of the tubes and they were allowed to stand for 3 minutes. This was to enable the eggs of the parasite to move upwards and attach onto the cover slip. The latter was removed and placed on a slide and observed under the microscope using the 4X objective to confirm the presence of Heligmosomoides bakeri (Behnke and Harris), [10] eggs. The cover slips and the slide were rinsed into a beaker using distilled water and the solution allowed to stand for two hours to enable sedimentation of the eggs. Furthermore, the supernatant was siphoned using a syringe and distilled water added and allowed again to stand for another two hours. After a series of rinsing to remove the salt solution, eggs considered free from salt were collected and stored in the refrigerator at 4°C for further use.
The eggs obtained from above were cultured according to the technique of Smyth. 3 ml of eggs suspension was poured into 2 petridishes with their bottom covered with filter paper. These petridishes were covered to prevent excessive evaporation of water and maintain a high relative humidity (65–67%). The petridishes were allowed to stand at room temperature of 25°C for 3, 4 and 5 days respectively with the aim of obtaining L1 and L2 larvae. These larvae were differentiated based on their morphological features and mobility. They were concentrated using the Baermann’s apparatus as described by [11,12].
Evaluation of the ovicidal and larvicidal properties
The ovicidal and larvicidal efficacy tests of the extracts on the free living stages of H. bakeri were carried out at three different levels according to [13-17].
All tests were repeated four times, each including the control.
Data analysis
The 50% inhibitory concentration (IC50) and the 50% larvicidal concentration (LC50) were determined from the linear regression curve obtained between larval mortality expressed in probit and decimal logarithm of extract concentrations (μg/ml). Comparisons of different proportions were made using the chi-square test and the results regarded as significant at P≤ 0.05.
Egg hatch assay
The effects of cold water, hot water and ethanolic extracts on the fresh eggs of H. bakeri are given in Figure 1. The mean inhibition rates were 0% for both distilled water (placebo) and 3% tween (negative control). Following our study, we observed that the cold and hot water extracts (CWE and HWE) remained the most effective, irrespective of the concentration compared to the controls which exhibited no inhibitory effect on the fresh eggs, on the other hand, the ethanolic extract (EtE) remained the least effective. Both CWE and HWE showed concentration dependent effects, in other words, their effects increased as concentration increased from 3750 to 5000 μg/ml. Both CWE and HWE induced a significant increase (P<0.05), while the effect of EtE was not significant (P>0.05). Mean embryonic inhibition rates stood at 59.2 and 74.8% for HWE and 74.2 and 75.6% for CWE respectively at 3750 and 5000 μg/ml (Figure 1).
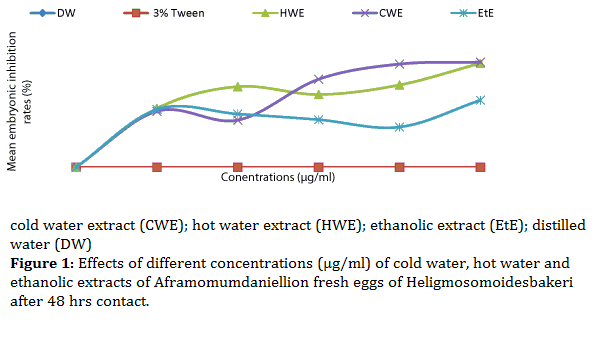
Figure 1: Effects of different concentrations (μg/ml) of cold water, hot water and ethanolic extracts of Aframomumdaniellion fresh eggs of Heligmosomoidesbakeri after 48 hrs contact.
At the end of this manipulation, unhatched eggs were considered dead. Different mortality rates were calculated with respect to the different concentrations. The transformation of mortality rates into probits and concentrations into log decimals gave a linear relation where from the equation of the straight line, we calculated the 50 % inhibitory concentrations(IC50). The values of IC50 were as follows: 1112.2, 1440.4 and 197.8 μg/ml for HWE, CWE and Et E respectively. As for the embryonated eggs, all the extracts showed a significant increase (P<0.05) in inhibiting hatching rates. The mean inhibition rates of embryonated eggs increased from 62.7 to 67.8% for HWE; 61.2 to 66.7 % for CWE and from 28.6 to 34.0 % for EtE respectively as the concentrations increased from 3750 to 5000 μg/ml (Figure 2) and the IC50 values were 63252.2μg/ml, 1079.6μg/ml and 17577.3μg/ml for HWE, CWE and Et E.
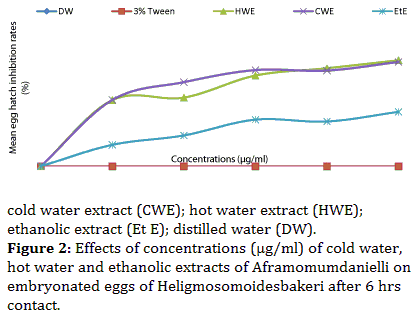
Figure 2: Effects of concentrations (μg/ml) of cold water, hot water and ethanolic extracts of Aframomumdanielli on embryonated eggs of Heligmosomoidesbakeri after 6 hrs contact.
Larval development
The EtE remained the most effective showing a general increase in trend. It induced a significant increase (P<0.05) in the L1 larval mortality rate. Its values increased from 86.7 to 97.3% as concentration increased from 3750 to 5000 μg/ml. The HWE equally induced a significant increase (P<0.05), its values equally increased from 38.2 to 73.9% as concentration increased from 3750 to 5000 μg/ml. On the other hand, CWE induced a non significant increase (P>0.05) in mortality. Meanwhile, the values of CWE decreased from 52.5 to 40.5 % as concentration increased from 3750 to 5000 μg/ml (Figure 3).
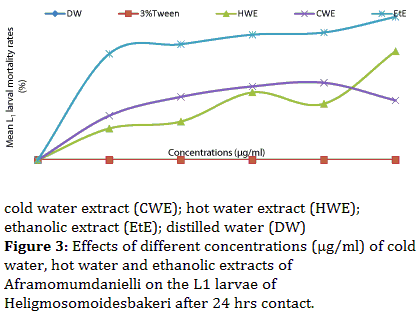
Figure 3: Effects of different concentrations (μg/ml) of cold water, hot water and ethanolic extracts of Aframomumdanielli on the L1 larvae of Heligmosomoidesbakeri after 24 hrs contact.
According to Figure 4, EtE remained the most effective extract showing an increasing trend as the concentration also increased. It induced a significant increase L2 larval mortality rate (P<0.05) from 92.0 to 100% as concentration increased from 3750 to 5000 μg/ml. The CWE also showed a significant larvicidal activity (P<0.05). Meanwhile, the HWE showed lower and non significant activity (P>0.05). The means larval mortality rate values increased from 58.8 to 66.5 for HWE and from 52.4 to 66.5 % for CWE respectively at concentrations 3750 and 5000 μg/ml.
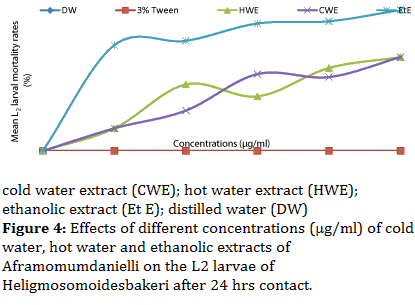
Figure 4: Effects of different concentrations (μg/ml) of cold water, hot water and ethanolic extracts of Aframomumdanielli on the L2 larvae of Heligmosomoidesbakeri after 24 hrs contact.
The mean embryonic inhibition rates on fresh eggs of H. bakeri obtained in the controls dishes (DW and 3% tween) are 0.0% each. These values indicated that they both had no inhibitory effect on the fresh eggs of H. bakeri. These results were similar to those obtained by Wabo Pone et al., [18] on the fresh eggs of Ancylostoma caninum (1.2%) and on the fresh eggs of H. bakeri (3.0%) respectively. The variation of mean embryonic inhibition rates observed inside the petridishes treated with the different plant extracts justified the fact that this plant possesses ovicidal properties. These values obtained from embryonation tests with the various concentrations of different extracts differed significantly from the negative controls. Generally, extracts of this plant presented an important inhibitory effect on the embryonic development of fresh eggs of H. bakeri. Similar findings were reported by Wabo Pone et al., [13] with extracts of Canthium mannii on fresh eggs of H. bakeri. The extracts of this plant had concentration dependent effects with concentrations greater than or equal to 3750 μg/ml. This observation was similar to the findings of Assisa et al., [19] on the ovicidal effects of Spigellia anthelmia on Haemonchus contortus eggs. In this study, the three plant extracts inhibited embryonation of fresh eggs. This activity was more effective with the ethanolic extract of A. danielli (IC50 = 197.8 μg/ml). These activities may be due to the fact that the active ingredients such as saponin, tannin, glycosides, polyphenols, flavonoides, terpenoides present in the different extracts penetrate the egg shell to stop the segmentation of blastomeres [20,14].The mean egg hatch inhibition rates of eggs of H. bakeri obtained in the controls were equal to 0.0 % each. This means that controls exhibited no effect on the embryonated eggs. The three extracts of this plant were concentration dependent with concentrations greater than or equal to 3750μg/ml. A similar finding was obtained by Wabo Pon e et al., [15] with extracts of Ageratum conyzoides on embryonated eggs of H. bakeri. The same finding was shown by Eguale et al. with extracts of A. schimperiana on Haemonchus contortus eggs. The CWE, HWE and the EtE extracts of this plant inhibited egg hatch mechanism. The highest effect was exhibited by the hot water extract (67.8 μg/ml). This value was a bit similar to the findings obtained by Jabbar et al., [21] on one hand, during their in vitro study with the aqueous and methanolic extracts of Chenopodium album and Caesalpinia crista and on the other hand by Adama et al., [22] during their in vitro study with the aqueous extracts of Anogeissus leiocarpus and Daniellia oliveri on the eggs of Haemonchus contortus. The fact that hot water extract was more effective was similar to the findings observed by Marie-Magdeleine et al., [23] with the aqueous extracts of the seeds of Curcuibita moschata on embryonated eggs of Haemonchus contortus. The fact that the larvae remained inside the embryonated eggs treated with various extracts may be justified by the fact that either the active ingredients of the extract penetrated the egg shell through osmosis or passed through the cuticular walls of the larvae into its circulatory system or the larvae on trying to hatch from the eggs into the external environment, absorbed water containing product prefers to remain inside the egg and die. The difference in the activity of the different extracts may be due to the different solvents used. In effect, both water and alcohol are polar solvents which allow the extraction of different polar compounds found in plants (Ciulei), [9]. Aqueous extracts contain heterosides and iridoids which are water hydro-soluble compounds affecting egg hatching, where as ethanolic extracts in addition to hydro-soluble compounds, also have lipid substances, alkaloids and polyphenols, terpenoids and tannin (Ciulei), [9] which also exhibit ovicidal and larvicidal properties. The controls (distilled water and tween 3%) did not kill the rhabditoide and non sheathed L2 larvae. This confirms the fact that the dead larvae found inside the treated Petridishes were due to active ingredients of the plant extracts, thus indicating their larvicidal properties.
The results obtained showed high mortality with concentrations greater than or equal to 3750 μg/ml. Also there was an increase in the mean larval mortality rates with increase in concentration. These observations are similar to the findings of Soetan et al., [24] when they worked on the anthelmintic effects of aqueous extracts of the seeds and leaves of African locust bean (Parkia biglobosa) on the bovine nematode eggs. Throughout this study, we observed that the L1 larvae were more vulnerable to the different plant extracts than the L2 larvae. This finding was contrary to a report by Soulsby [33]. The larvicidal properties of the extracts may be due to the penetration of active compounds across the cuticular linings of the larvae, due to the soft nature of their cuticle on one hand or the absorption of the substance by the larvae on the other hand. Enriquez [25] mentioned that the active compounds penetrate the cuticle of the nematode and prevent the absorption of glucose or block the post-synaptic receptors, thus, paralysing the larvae. These active compounds can also stimulate the secretion of glutamate and gamma amino-butyric acid (GABA) which may block the transmission of nervous impulses or decoupling the phosphorylation oxydative reactions, which led to energy exhortation of the larvae thus leading death Wabo Pone et al., Kabore et al., [16,26], mentioned that the active compounds found with food can cross the intestinal lining of the larvae and gain access to the circulatory system of the organism. Also the active compound like tannin may bind to the cuticle of the nematode, destabilize the membrane and increase cell permeability by combining with membrane- associated sterols Price et al., [27] which leads to death. The three plant extracts had larvicidal properties with the most effective LC50 values (247.7 and 426.9 μg/ml) obtained with the ethanolic extracts for L1 and L2 respectively.
Ovicidal and larvicidal properties of our plant extracts may be due to the presence of secondary metabolites like alkaloids, saponin, polyphenols, carotenoids, tannin, coumarine, cardenolides, triterpenes, saponoside, embeline, sesquiterpen, lactones. Others researchers such as Hounzangbe-Adote et al., Katiki et al., Eguale et al., Wabo Pone et al., [28,29,13-17] have shown that carotenoids, triterpenes, saponin steroids coumarines, tannins and other chemical compounds obtained from plants like glycosides, enzymes, anthraquinone, essential oils, lipid, protein, fibers if present on a plant implies that plant possesses anthelminthic properties.
From the above results, we concluded that, the aqueous and ethanolic extracts of A. danielli proved to have ovicidal and larvicidal properties. They were able to inhibit the embryonic development and egg hatching as well as larval survival of H. bakeri. Further experiments incorporating in vivo studies are required to find out and establish the effectiveness and pharmacological rationale for the use of A. danielli extracts as anthelminthic drugs.
The authors express their profound gratitude to the Ekpor and Sabes communities and particularly to Mr Tacham Walters and late Mr Tiku Daniel Kereh for helping in the collection of the plant specimen and also to the Laboratory of Biology and Applied Ecology (LABEA) of the University of Dschang for providing the parasite strains used in this study.
We declare no conflict of interest.