Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2013) Volume 2, Issue 2
Plant parasitic nematodes are major threat for crop plants and cause severe economic losses worldwide annually. Various strategies deployed for the control of these notorious parasites has resulted either in limited success, or having huge negative impact on environment. RNA interference (RNAi) is a gene-silencing phenomenon that is conserved in various eukaryotes. Experimentally induced RNAi is highly specific and potent, leading to its wide utilization in functional studies for exploring gene functions. Crops engineered through RNAi have proven to be successful in protection against pest and parasites, including nematodes. Engineering nematode resistance in crop plants through host-derived RNAi is largely based on the selection of target gene. The expression of nematode specific dsRNA in plants generates siRNAs and taken up by nematodes on feeding. Depending upon the function, level of expression and silencing efficacy of the target gene, resistance was determined. RNAi seems to be promising in many aspects, such as providing durable resistance to crops against plant parasitic nematodes in the near future. In the present article, we have reviewed the published work on the host-derived RNAi for developing nematode resistance in plants.
<Keywords: RNAi, dsRNA, siRNA, Host-derived RNAi, Plantparasitic nematodes, Nematode resistance
Plant parasitic nematodes (PPN) are major agricultural pests, which infest almost all cultivated crop plants causing drastic economic loss estimated as ~ 157 billion US$ [1]. Clear symptoms of infections are often deficient, leading the nematodes unnoticed by farmers, until the infestation is rigorous. Moreover, nematode infection often results in secondary diseases to the host plant, leading to the underestimation of the exact crop losses attributed by PPN [2]. Among plant parasitic nematodes, most of the damage is often contributed by sedentary endoparasitic genera Meloidogyne spp (root knot nematode (RKN)), Globodera and Heterodera spp (cyst nematode). Root knot nematodes are able to attack wide range of crops (~ 2000), while each species of cyst nematodes has a narrow host range. These parasites invade plant roots as mobile infective second juvenile stage (J2) to reach plant vascular cylinder, where they migrate intercellularly (root-knot nematode), or intracellularly (Cyst nematode), to become sedentary and induce the formation of feeding cells that supports the development and reproduction of females over a period of few weeks. Depending on the nematode species, the feeding cells either develop into specialized multinucleate giant cells (root-knot nematode) or syncytia (cyst nematode), by inducing mitosis without cytokinesis, leading to the formation of galls and subsequently, plant dies after several cycles of nematode infection [3-5].
There are several methods widely used for controlling these notorious parasites, such as chemical nematicides, cultural practices, organic means and use of resistant cultivars. Although chemical nematicides are effective means of nematode control in recent decades, issues such as mammalian toxicity, ground water contamination, environmental toxicity and residues in food have caused much stringent rule for their use in agriculture, and in many countries, effective nematicides have been banned for commercial use [2]. Utilization of green manures for nematode control was also carried out in field conditions, but assessing their effectiveness remains challenging. Cultural practices such as crop rotation are mostly used for controlling nematode pests in field conditions, but certain nematodes remain dormant for many years, e.g. cyst nematodes. The cover crops, particularly marigold, have been recommended for nematode suppression. However, marigold as an intercrop has its own limitations, i.e. it is sensitive to short-day photoperiod, and also serves as host for some other pests. For limited range crops, resistant cultivars have been a commercial success, but invariably this approach is unable to control many nematode problems. Few naturally occurring resistant genes have been identified and cloned for developing nematode resistance [6]. However, attempts to transfer resistance from one crop to others have so far been unsuccessful, for reasons remain unclear. Further other approaches, including traditional breeding to develop nematode resistant plants, have resulted in limited success [7]. Therefore, there is an urgent need for developing a novel strategy for successful mitigation of these plant parasitic nematodes. In this regard, RNA interference (RNAi) technology has proven to be novel and successful for the control of various pests and parasites, including nematodes.
RNAi is an apparently ancient defense mechanism against invading viruses, prevents deleterious effect of transposon movement, regulation of gene expression and chromatin modification. It is a powerful tool to suppress gene expression and analyze gene function. RNAi operates at transcriptional (called as transcriptional gene silencing-TGS), as well as post-transcriptional (called as post-transcriptional gene silencing- PTGS) for gene silencing, which has been previously reported as cosuppression in plants [8], and quelling in fungus [9]. In fact, these three techniques appeared to be remarkably well conserved in several eukaryotes, which is initiated by the action of dicer enzyme on dsRNA, leading to the production of small interfering RNAs (siRNAs), and these siRNAs along with the RNA induced silencing complex (RISC) are involved in sequence specific silencing of the target mRNA (Figure 1) [10-12]. Stoichiometric considerations revealed that a few dsRNA molecules are enough to cause RNAi phenotypes, leading to the discovery that the silencing response can be amplified through the action of RNA-dependent RNA polymerases (RdRPs) on the targeted mRNA, which results in the synthesis of secondary siRNAs [11,13]. In fact, complementary RNA (cRNA) synthesis allows siRNA formation and homology-dependent silencing to spread outside of the initial inducer sequence, a process called transitivity. However, the precise mechanism involved in RdRP-mediated synthesis of secondary siRNAs appears to vary in different organisms. Secondary siRNAs have 5’-triphosphate, are antisense to the target transcript, and are associated with specific Argonaute proteins [14]. In case of C. elegans, transmembrane proteins SID-1 and SID-2 are independently for dsRNA ingestion mediated RNAi. SID-1 is required for the dsRNA uptake into all cells, while SID-2 is required for silencing initiated by dsRNA [15].
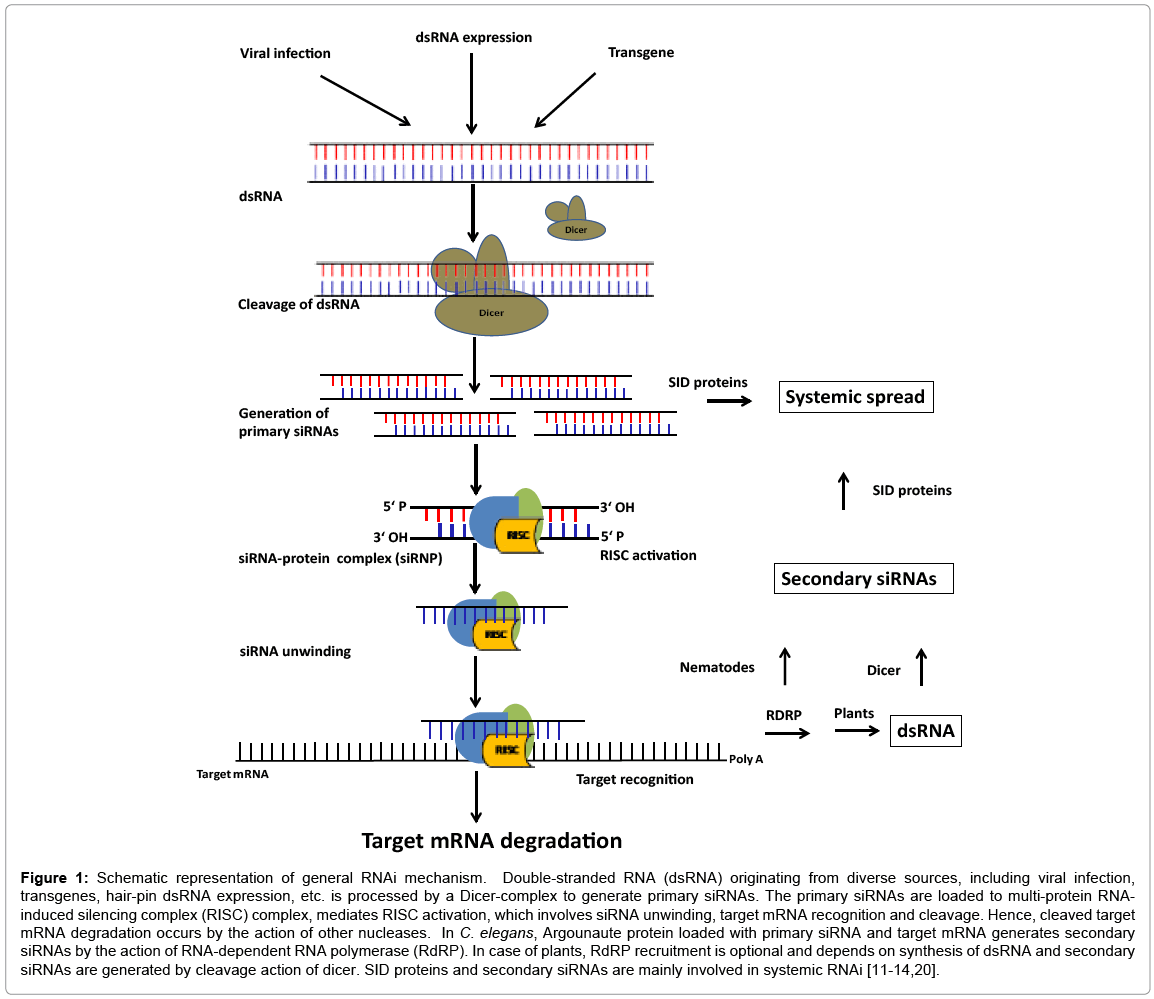
Figure 1: Schematic representation of general RNAi mechanism. Double-stranded RNA (dsRNA) originating from diverse sources, including viral infection, transgenes, hair-pin dsRNA expression, etc. is processed by a Dicer-complex to generate primary siRNAs. The primary siRNAs are loaded to multi-protein RNAinduced silencing complex (RISC) complex, mediates RISC activation, which involves siRNA unwinding, target mRNA recognition and cleavage. Hence, cleaved target mRNA degradation occurs by the action of other nucleases. In C. elegans, Argounaute protein loaded with primary siRNA and target mRNA generates secondary siRNAs by the action of RNA-dependent RNA polymerase (RdRP). In case of plants, RdRP recruitment is optional and depends on synthesis of dsRNA and secondary siRNAs are generated by cleavage action of dicer. SID proteins and secondary siRNAs are mainly involved in systemic RNAi [11-14,20].
RNAi has been successfully utilized for exploring gene functions in many eukaryotes, including nematodes. Induction of RNAi by delivering dsRNA has been successful in non-parasitic nematode, Caenorhabditis elegans, and majority of its genes has been characterized using this technique [16]. Delivering dsRNA to plant parasitic nematodes has been technically challenging, because these obligatory parasites requires living host for feeding and the pre-parasitic nematodes are of non feeding and do not ingest fluid at this stage. Several house keeping and parasitism genes were silenced in vitro for functional studies using this approach are well documented, has been reviewed extensively earlier [7,17].
Initial reports highlighted the utility of RNAi for functional studies in plant parasitic nematodes was achieved by treating dsRNA/siRNAs to the infective stage juveniles, and flurescine isothionate (FITC) was commonly utilized to monitor the up take of dsRNA/siRNAs. In most of the cases, octopomaine, resorcinol, spermidine and serotonin were used to stimulate the uptake of dsRNA from solution [17]. Alternative route for dsRNA up take may also exist in plant parasitic nematodes. Silencing of Meloidogyne artiellia chitin synthase gene was achieved by soaking the intact eggs in dsRNA solution, resulted in depletion of transcripts and delayed hatching of juveniles from treated eggs [18]. Profound silencing in terms of reduced transcript level and abnormal phenotypic behavior could be induced by targeting neuronally expressed genes using various sizes of dsRNA (88 bp, 222 bp, 227 bp, 316 bp) in water, without any stimulated ingestion. This has been suggested that alternative dsRNA up take route were also possible through sensory organs (amphids), secretory/excretory pore, or even the cuticle [19].
Due to grand success in functional genomics, RNAi has emerged as versatile tool with wide range of applications in plant biology. Recent developments in RNAi led to the identification of core RNAi components in many eukaryotes, and have envisioned researchers to adopt this tool for wider applications in crop improvement, including engineering crop plants against plant parasitic nematode menace. In the past decade, various studies indicated key developments in the RNAi machinery, and also whole genome sequence data available for few of the plant-parasitic nematodes [3,20], provides a new horizon for the successful exploitation of RNAi for nematode control.
Expression of dsRNA, corresponding to the nematode target gene through host plant, mediates the production of siRNAs. Based on the sequence similarity between the siRNAs and the corresponding target mRNA, silencing is achieved upon nematode feeding. Although most of the reports successfully utilized RNAi for studying the gene function in vitro, however, they were not technically feasible to use as nematode control under field conditions. And major hurdle for potential application of plant-mediated RNAi was whether nematodes ingest dsRNAm when feeding on plant cells expressing dsRNA constructs. These plant parasitic nematodes produces specialized feeding structures inside the plant cell on which they feed through out the development called feeding tube, act as a molecular sieves, and may helps in restricting nematodes from ingesting plant defense related macromolecules [21,22]. And the size exclusion limit is accurately unknown, but it has been reported that up take of GFP (28 kDa) and different other dyes indicated that a size exclusion for these plant parasitic nematodes are between 20 and 40 kDa [17,23]. One recent report indicated that RKN species (M. hapla) infective stage juveniles could ingest a range of Bacillus thuringiensis proteins sizes from 45 kDa to 140 kDa, directly through the stylet under in vitro condition [24]. Moreover, it has been proven that linear dsRNA/siRNA was possibly taken up by plant parasitic nematodes [25-37]. And also, feeding tube would be possibly excludes both siRNA incorporated into the RISC, as well as non-linear dsRNA molecule [17-19,21]. The delivery of parasites gene-specific dsRNA through the host may be more effective and appropriate method for nematode control.
Host-delivered RNAi appears to be most promising strategy for nematode control. Based on the choice of the target gene selected, it can be broadly categorized as dsRNA against the targets, which are constitutively expressed throughout nematode life cycle, genes involved in nematode development, neurotransmission, nematode effectors/ parasitism genes (Table 1). Some of these nematode target genes have been validated experimentally through in vitro RNAi experiments in C. elegans and other plant parasitic nematodes [15,17,25-38]. On the other hand, soon after the nematode infection, there are number of genes differentially regulated in host cells, which are responsible for feeding site establishment, and also helps in successful parasitism [39]. Genes involved in the feeding site formation and other plant genes including those involved in cell cycle, miRNAs which are differentially regulated during nematode infections are of primary interest. There is no direct evidence reported till now for the utilization of candidate microRNAs for nematode control.
| Species | Target Gene | Promoter-the control of | Host Plant | Target mRNA depletion | Observed phenotype | Time point | Reference |
|---|---|---|---|---|---|---|---|
| M. incognita | Integrase Splicing factor | 35S | Tobacco | Yes | Decreased number of established nematode | 6-7 wpi | [40] |
| M. incognita M. javanica M. hapla M. arneria | Secreted peptide 16 D 10 | 35S | Arabidopsis | Not detected | Decreased number of galls & gall size | 4 wpi | [30] |
| H. glycines | Major sperm protein | ACT2 | Soybean | Not detected | Decreased number of eggs | 8 wpi | [47] |
| M. javanica | Putative transcription factor | 35S, TobRB7 | Tobacco | Yes | - | 6 wpi | [43] |
| H. schacbii | 4G06, ubiquitin-like | 35S | Arabidopsis | Yes | Decreased number of females | 14 dpi | [44] |
| H. schacbii | 3B05, cellulose binding protein | 35S | Arabidopsis | Yes | - | 14 dpi | [44] |
| H. schacbii | 10A06, Zinc finger protein | 35S | Arabidopsis | Arabidopsis | Low number of developing females | 14 dpi | [44] |
| H. schacbii | 8H07, SKP1-like | 35S | Arabidopsis | Yes | Decreased number of developing females | 14 dpi | [44] |
| H. glycines | Ribosomal protein 3a,4. Spliceosomal SR protein | FMV-sgt | Soybean roots | Not detected | Reduction in number of females | 8 dpi | [41] |
| M. incognita | Tyrosine Phosphate, itochondrial stress -70 protein precursors MSP | Soybean roots | Yes | Decreased number of galls, 5-6 fold reductions in mature female diameter | 14 dpi | [44] | |
| H. glycines | Fib 1,Y 25 | 35S | Chimeric soybean root system | Yes | Suppression of nematode reproduction & development | 5 wpi | [42] |
| H. glycines | Cpn 1 Y25,Prp 17 | 35S | Composite soybean plants | Yes | Reduction in reproduction & development | 5 wpi | [48] |
| M. incognita | Rpn7 | 35S | Tomato hairy roots | Yes | Reduction in reproduction & motility | 40 dpi | [37] |
| M. incognita | Parasitism gene 8D05 | 35S | Arabidopsis | Not detected | Reduction in gall number | 8 wpi | [45] |
| M. incognita | Calreticulin - MiCRT | 35S | Arabidopsis | Yes | Reduction in gall number | 8 wpi | [46] |
*dpi-days post infection; wpi-weeks post infection
Table 1: Host-derived RNAi for the control of plant parasitic nematodes.
House-keeping genes
These are constitutively expressed throughout the nematode development and implicated in major cellular processes. Targeting these genes through host-generated dsRNA ultimately hampers the nematode development, and hence, impedes parasitic success. The first demonstrated success of host-delivered RNAi for the nematode control was achieved by targeting splicing factor and integrase. Tobacco plants expressing dsRNA of these two genes under the control of constitutive promoter resulted in target gene depletion and also mitigate the nematode (Meloidogyne incognita) establishment [40]. Also, soybean roots expressing RNAi constructs targeting three essential genes involved in mRNA metabolism Heterodera glycines RPS-3a, RPS- 4 and SPK-1 displayed variations in nematode infection, i.e. 80-88% reduction in nematode numbers [41]. In other report, transgenic soybean expressing PRP 17 gene resulted in significant reduction in nematode (H. glycines) infection (53%) and (79%) reproduction. In the same study, two of the other genes Cpn 1 were found to be embryonic lethal, in case of C. elegans was selected, and dsRNA corresponding to these genes expressed through composite soybean plants displayed up to 95% reduction in number of egg masses developed to H. glycines infection was observed [42]. Based on these reports, it may be enticing to choose such genes; some precautions should be administered before selecting such kind of targets. Most of the houses keeping genes are highly conserved across different species, which may have off-target effects on host plant/other beneficial organisms/humans. Transgenic tobacco plants expressing different dsRNA hairpin structures targeting RKN (Meloidogyne javanica) putative transcription factor MjTIS-11, leads to the down-regulation of transcripts in nematodes feeding on the transgenic RNAi lines. Even though the presence of siRNAs in transgenic plants was detected, but it does not lead to any lethal phenotypes [43]. This result highlighting the most important fact, that not all genes are amenable for developing nematode resistance through host-derived RNAi. Hence it is imperative to look for novel and potent targets for successful exploitation of RNAi-mediated nematode control.
Nematode-specific genes
These are most attractive targets for engineering crop plants for nematode resistance through host-derived RNAi. The parasitism/ effector genes which are unique to nematodes have been already identified, available in public databases, and targeting such genes could provide durable resistance with less safety related issues. In fact, utility of such nematode specific targets for developing nematode resistance through plant-mediated RNAi has already been explored. Transgenic Arabidopsis plants expressing dsRNA of sub-ventral esophageal gland cell secreted protein 16D10 conserved region of RKN displayed 65-90% reduction in number of galls, with reduced gall size and corresponding reduction total egg production [30]. Based on their observation, either full length dsRNA or processed siRNAs were sufficient to induce an RNA silencing phenotype. Same strategy has been proven to be successful in Arabidopsis; although no complete nematode resistance was observed when unique and conserved regions of parasitism genes were targeted through in planta RNAi. Reduction of developing nematodes ranged from 23-64% in different RNAi lines. In fact, in planta dsRNA/siRNA up take in cyst nematodes appears to be functional because of the specific down-regulation of target mRNA, and nematodes feeding on RNAi plants were less successful in further development, and also led to a reduction in number of mature females [44]. Silencing of parasitism gene 8D05 of M. incognita through host derived RNAi in Arabidopsis resulted in drastic reduction in gall number [45]. Also, targeting M. incognita calreticulin (MiCRT) through transgenic expression of dsRNA in Arabidopsis resulted in target gene reduction and decreased gall number [46].
Genes involved in development and reproduction
There is increasing evidence based on the sequence data, that it is now possible to utilize genes which are involved in nematode development and reproduction. However, there are few examples available, as they found to be involved in reproduction, and are also specific to nematodes. One such target is major sperm protein (MSP) and silencing soybean cyst nematode (H. glycines) MSP through transgenic soybean derived dsRNA displayed 68% reduction in egg production. Moreover, the progeny of the hatched from the eggs of those nematodes feeding on transgenic RNAi lines apparently displayed 75% reduction of egg production, when allowed to infect susceptible wildtype soybean plants. These results strengthen the knowledge about the in planta RNAi can be successfully used to control cyst nematodes as well [47]. Moreover, transgenic soybean expressing dsRNA fragments specific to genes related to H. glycines reproduction or fitness (CPN- 1, Y25, PRP-17), significantly suppressed the nematode reproduction and development [48]. RNAi constructs targeting four different genes (lactate dehydrogenase LD, tyrosine phosphatase TP, mitochondrial stress 70 protein precursor MSP and ATP synthase beta chain mitochondrial protein), of M. incognita, which has high homology with cyst nematodes, and C. elegans genes were transformed with soybean roots and investigated for nematode infection. Out of the four RNAi constructs tested, constructs targeting MSP and TP were showed up to 94% reduction in gall formation [49]. In another report, four RNAi lethal homologues of C. elegans were selected, encoding ribosomal protein 3a and 4, a spliceosomal SR protein and synaptobrevin of H. glycines, and expressed as tandem inverted repeats in transgenic soybean plants inhibited the female development [41]. Similar strategy was deployed by another group, to silence the genes encoding a beta subunit of coatomer (COPI) complex, pre mRNA splicing factor and protein with unknown function, which has led to hampering nematode infection and female development of H. glycines [38]. Although, all the reported experiments on plant-mediated nematode resistance has utilized transgenic plant expressing empty RNAi vector as control in evaluating the resistant phenotype, but most care should be taken in terms of considering many more factors to reduce the unintended noise of off-target effects.
Nematode responsive host plant genes
Based on the existing literature, it is now clear that plant-nematodes have evolved different strategies to facilitate compatible interaction with the host plant and mediates their infection successfully [4,21]. Nematodes secretes a variety of effectors into host cell performing numerous functions, including cell wall degrading, suppression of host defenses and interact with host signaling pathways to facilitate feeding site formation [21]. Soon after plant parasitic nematode infect the roots, many number of host genes becomes up-regulated. This, in turn, facilitates the feeding site establishment and also helps in successful parasitism [50]. Down tuning the expression of those nematode responsive plant genes may be a vital strategy for the control of plant parasitic nematodes. Recently, Cappelle et al. [51] have reported that silencing of nematode induced Arabidopsis thaliana cyclin dependant kinase (AtCDKA; 1) gene under the control of nematode responsive promoter WRKY23 resulted in reduced transcript abundance, and also led to retardation in reproduction of cyst and RKN. Although silencing was observed, significantly fewer galls developed relative to wild-type control plants, but it has not provided complete resistance. Limited success for providing complete resistance could be because of the weakness of the promoter or multigene nature. Targeting multiple host genes involved in the feeding site establishment, under the control of highly specific nematode responsive promoters for stringent control of dsRNA expression, could be appropriate for complete resistance. Because those genes involved in feeding site establishment were detrimental for plant growth, silencing multiple genes may result in deleterious effect on plant development [17]. Another possible strategy for successful nematode control in near future may be through the microRNAs and other small RNAs [39]. A recent report indicated the cyst nematode induced small RNAs and their targets in model plant Arabidopsis [39]. Interestingly, Dicer (DCR) like and RNA-dependent RNA polymerase (RDR) mutants of Arabidopsis exhibited decreased susceptibility to H. schachtii infection, suggesting the possible role of small RNAs in plant nematode interaction. Also, nematode induced small RNAs, including microRNAs and their corresponding targets have been identified. The authors showed that there was a negative correlation between the smallRNA accumulation and their corresponding target transcript abundance. Moreover, it has been suggested that nematode induced microRNAs and small RNAs likely to be involved in feeding site establishment and parasitism, respectively [39]. Over-expression of those nematode induced candidate microRNAs, and/or silencing of their corresponding targets, may provide clear insight about plant-nematode parasitism, and also may provide nematode resistance to crop plants.
High specificity degradation of target mRNA results in effective silencing, which eventually leads to lethal for the pests. Even single mismatch between the target mRNA and corresponding siRNA inhibits gene silencing [52]. In general, terminal bases are tolerant to mismatches, whereas the central bases are not. 9-11 bases are least tolerant to mismatches [53]. Silencing can be induced by the 30-50 nt similarity, with the off-target gene results in silencing in C. elegans [54]. In order to paramount such off-target effects, choice of target gene should be specific to the pathogen/pest, and could have minimal/no sequence homology with the sequences of plants, animals, beneficial insects, as well as free living nematodes [38].
• It is possible to silence gene families with single sequence, in case the selected target sequence in conserved among gene families [55,56]. By targeting several genes with the single gene construct should correlate with greater silencing effect.
• In general, genetic resistance is based on multigenic complex, difficult to transfer it to susceptible cultivars. RNAi strategy could possibly create a single resistant locus that will facilitate the transfer of nematode/pest resistance to other cultivars [38].
• Long range stability of pest resistance may also be possible by deploying RNAi. Based on the choice of target gene, as well as sequences used for the RNAi construct, stability can be maintained, i.e. especially mutation in the selected target gene for developing RNAi construct would considered to be lethal. Also, it is possible to maximize the effectiveness for durable resistance by selecting target gene sequences that show high homology among the nematode population. However, longterm field and greenhouse studies will be needed to verify stability. In order to tackle the RNAi tolerant nematode population caused by random mutation, RNAi locus can be engineered to target multiple genes [38].
Although RNAi has emerged as a potent and successful technology for crop protection, there are few limitations which need to be properly addressed before adopting this technology. The important factors contributing to the effectiveness and possible solutions to overcome those major limitations were reviewed extensively earlier [57]. Major concern regarding the wider employment of RNAi-based crop protection is off-target effect; host-derived dsRNAs/siRNAs might interfere with the expression of the other non-intended target genes of the host itself, beneficial organisms and humans, etc. based on sequence homology. In order to circumvent the off-target effects, in-depth in silico homology searches were employed, and the region which is having no/minimal homology with the available sequences of the host, as well as other organisms were widely utilized for developing hairpin RNAi constructs [43,55,56]. Some online tools are also available for such analysis, including siRNA scan, pssRNAit, etc. However, the experimental approaches using qPCR and microarray can be deployed to evaluate the off-target effects.
Although RNAi-mediated nematode control in crop plants appears to be the most promising strategy, most of the above discussed methodologies did not provide complete resistance against plant parasitic nematodes. One of the possible ways to achieve complete resistance is through the use of chimeric RNAi constructs targeting multiple genes involved in single or multiple pathways of plant parasitic nematodes/nematode responsible plant genes, under the control of tissue-specific or wound-inducible or nematode-inducible promoter. In fact, the simultaneous silencing of multiple genes has been successfully demonstrated in Drosophila [58], C. elegans [59], and having strong therapeutic potential against viral infections and cancers [60]. Further, the enhanced resistance to plant parasitic nematodes by combining the expression of dsRNAs by crossing appropriate Arabidopsis thaliana lines resulted in an additive effect that further reduced nematode numbers and developmental capacity [61]. It is also notable that silencing miRNA pathway genes led to nematode resistance under in vitro conditions [62]. Targeting microRNA pathway, and/or miRNA genes through plants may provide durable and complete resistance against these notorious plant parasitic pests. And also complete nematode resistance may also be achieved through plant expressing artificial miRNA (amiRNA), containing known miRNA genes with the replaced seed region of the vital gene of plant parasitic nematodes. The primary advantage of amiRNA technology over dsRNA mediated silencing is that specificity, wherein long hairpin constructs were cleaved into multiple fragments, which can cause unintended silencing of non-target genes [38]. Interestingly, in a recent report, amiRNA-mediated silencing of endogenous soybean gene (Rhg1 locus of LRR-K) caused detectable reduction in target mRNA abundance, but as expected, there was no significant impact on soybean cyst nematode infectivity were detected [63]. Recent development in molecular techniques utilized for studying gene expression analysis, including next generation analysis, including microarray, lacer capture microdissection and next generation sequencing opened new era in studying plant-nematode interactions, which generated enormous data. Proper mining of these available resources may provide new insights in developing RNAi-mediated nematode resistance in crop plants. On the other hand, genomic and EST sequences are available for few of the important plant-parasitic nematodes, which provide valuable resource for researchers to explore possible potent and specific RNAi targets for successful control of plant-parasitic nematodes. RNAi-mediated crop protection against nematode pests appears to be most promising than other available methods, in terms of efficiency, stability and its ability to suppress gene expression in a regulated manner. Moreover, it appears to be most promising strategy with lesser environmental impact. Further, substantial research on feasibility studies are necessary for predicting ecological risk assessments before transforming this technology to field conditions.
Manchikatla Venkat Rajam is grateful to the Department of Biotechnology and Department of Science and Technology (DST-Govt. of India), New Delhi, University of Delhi, Delhi, Bejo Sheetal Seeds Pvt. Ltd., Jalna, Maharashtra, India and Sri Biotech Laboratories India Ltd., Hyderabad, India, for generously funding his research programmes on RNAi. The authors thank University Grants Commission for Special Assistance Programme and DST for FIST programme.