Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2013) Volume 2, Issue 2
Keywords: Fish, Fish processing, Fish waste, Protein, Oil, Alcalase, Neutrase, Spray drying, Catalytic triad, Serine proteases, Endoproteases, Jadomycin
Amino acids are used as food additives in the food and animal feed industries, as well as in pharmaceutical applications, such as jadomycin production. Jadomycin is a secondary metabolite produced by the Streptomyces venezuelae ISP5230 under specific nutrient and stress conditions, such as heat, alcohol and phage shocking [1]. The Jadomycin production process takes place in three steps: growth, shocking and production. During the growth phase, a culture of Streptomyces venezuelae ISP5230 is grown in nutrient rich MYM medium at 30°C for 48 h. The culture is later transferred into nutrient deprived amino acids rich production medium. The shocking of the bacteria is carried out by ethanol, temperature or phage. After shocking, Streptomyces venezulae convert amino acids into Jadomycins [2].
The traditional sources used for production of amino acids include hair, keratin, feather, blood meal, soya bean and wheat gluten [3]. These sources cannot keep up with the increasing demand for amino acids. Therefore, microbial and chemical processes are currently used for the production of amino acids [4]. These current techniques include microbial fermentation, enzymatic synthesis, chemical synthesis and protein hydrolysis. Around 11 amino acids are produced by microbial fermentation process, using different microorganisms [4-6]. The cost of microbial production of amino acids depends upon the carbon source, fermentation yield and purification process. Advances in the fermentation technology and improvement in the strain development have allowed industrial scale production of amino acids by microbial fermentation [7]. The main disadvantage of microbial fermentation is that the entire fermentation industry is dependent on glucose or sucrose as carbon sources. A feasible and cost effective alternative, such as using alcohols (ethanol and methanol), instead of sugars (glucose and sucrose) as a substrate for production of amino acids, using bacteria, has not been implemented on a large scale [8].
The enzymatic methods are used in small scale industries to produce synthetic D- and L- amino acids. Enzyme membrane reactors are predominantly used to produce amino acids, such as Alanine, Methionine, Valine, Phenylalanine, Tryptophan, Malic acid, Aspartic acid and Phenyl lactic acid. The advantages of using enzyme membrane reactor include: (a) continuous processes for the production of amino acids, (b) higher productivity and improved rate of reaction and (c) use of multi-enzyme systems and multiphase reactors. The disadvantages include: (a) the enzymes cannot withstand high temperatures, (b) lack of operational stability, (c) specificity of the enzymes, (d) enzyme poisoning, (e) enzyme deactivation, and (f) enzyme leakage in immobilized systems [9,10]. Also, the substrate used in the enzyme membrane reactors was mainly obtained by chemical synthesis [6].
Chemical synthesis is carried out to produce D, L-forms of amino acids, using sugar beets or acrylonitrile as a substrate. The advantage of this process is that it can be carried out in a continuous manner on a very large scale; whereas the main disadvantage is that the amino acids produced will be in a racemic mixture of both D and L-forms, which has to undergo a costly optical resolution step. Glutamic acid is produced using this process on a large scale [5].
Protein hydrolysis is carried out using acid (6 N HCl and 110°C), alkali (6 N NaOH and 110°C), or enzymes. During acids or alkali hydrolysis, all the peptide bonds are broken, and a complete hydrolysis of proteins takes place in a short period of time (24 h). It is one of the preferred processes for hydrolysis of vegetable proteins. The main disadvantages of this process are that aspartic acid and glutamic acid are completely destroyed and racemization of amino acids occurs [11]. The enzymatic hydrolysis is carried out to cleave the peptide bonds using a combination of proteases, which allows the quantification of Aspargine and Glutamic acids. However, the application of this process on a large scale is hampered by: (a) the specificity of the proteases to cleave the peptide bonds, (b) a combination of proteases are required for complete protein hydrolysis process, and (c) longer incubation time of 24-48 his required [11,12].
Fish are usually processed before being used for human consumption. There are two types of waste produced during fish processing: solid waste and liquid waste. The solid waste consists of head, frames, fins, tails, skin and gut, and the liquid waste consists of wastewater used during processing. These fish wastes are a rich source of many value added products such as proteins, amino acids, bioactive peptides, collagen, gelatin, oil, calcium and enzymes. Fish waste typically comprises of 58% proteins and 19% ether extract or fat, in addition to minerals, such as calcium, phosphorous, potassium, sodium, magnesium, iron, zinc, manganese and copper [13]. However, fish waste is underutilized, and is being dumped in the ocean causing environmental problems. In the year 2011, 154 million tonnes of fish were produced in Canada, of which 130.8 million tonnes were used for human consumption and 23.2 million tonnes were used for non-food uses [14]. Approximately, 758,090 tonnes of fish waste were dumped in sea during the period of from April 2000 to March 2011 in Canada [15]. The proteins in the fish waste can be extracted and converted to amino acids that can be used for jadomycin production.
The preceding work investigate the use of fish proteins (obtained from fish waste), as a substrate for producing amino acids for use in jadomycin production. The emphasis of this work is to demonstrate the use of enzymatic hydrolysis process for extracting amino acids from fish proteins, and evaluate the feasibility of jadomycin production from the extracted amino acids. The enzymes used in this study were Alcalase and Neutrase.
The main aim of this research was to evaluate the enzymatic extraction of amino acids (using the enzymes Alcalase and Neutrase) from fish protein, for use as a substrate for production of jadomycin. The specific objectives to: (a) evaluate the effectiveness of the enzymes Alcalase and Neutrase individually, and in combination on the extraction of amino acids from fish protein, (b) study the effect of hydrolysis time (24 and 48 h) on the enzymatic hydrolysis of protein, (c) to determine the profile of amino acids extracted from fish protein using Thin Layer Chromatograph (TLC), and (d) to characterize the amino acids extracted from fish protein using Gas Chromatography- Flame Ionization Detector (GC-FID).
Fish protein
The fish protein was obtained by spray drying the fish protein hydrolysate extracted from Mackerel fish waste, using the procedure described by Manoj and Vasudevan [16].
Chemicals and enzymes
The enzymes used in the study (Alcalase and Neutrase) were obtained from Sigma-Aldrich, Oakville, Ontario, Canada. The standard amino acids were obtained from Bioshop Canada Inc., Burlington, Ontario, Canada. The reagents included: 1 N hydrochloric acid, 6 N hydrochloric acid, 1 N sodium hydroxide, 20% Trichloroacetic acid, 1 M potassium phosphate monobasic and 1 M potassium phosphate dibasic. 1 M pH 8 phosphate buffer was prepared by adding 94.7 ml of 1 M potassium phosphate mono-basic and 5.3 ml of 1 M potassium phosphate dibasic with 100 ml of distilled water.
Hydrolysis of protein
The protein hydrolysis process was carried out in order to extract amino acids from the protein derived from mackerel fish waste as shown in Figure 1. Four grams of dried protein samples were weighed using a digital scale (Metler AE 200, Mettler-Toledo International Inc., Mississauga, Canada), and added to 100 ml of distilled water making a 4% solution. Several solutions were made and placed in a water bath shaker (Precision 2870 Series water bath shaker, Thermo Scientific, Ohio, USA), which was preheated to 55°C. The samples were kept in the water bath for 30 min. The Alcalase enzyme was added to the samples at the rate of 20 µL/g of protein. The hydrolysis process was continued for either 24 h or 48 h at 140 rpm. The samples were heated in a water bath (Precision 280 Series, Thermo Scientific, Ohio, USA) for 5 min at 90°C to deactivate enzyme. The samples were then cooled and analyzed using Thin Layer Chromatography ((TLC) (Sigma-Aldrich, Oakville, Ontario, Canada)).Samples were spray dried (Mini Pulvis Spray GS- 310, Yamato Scientific America, USA) at an inlet temperature of 130°C, an outlet temperature of 90°C and a flow rate of 2 ml/min. The spray dried amino acids were analyzed using gas chromatography (Hewlett- Packard HP5890 Series II gas chromatograph, coupled with Flame Ionization Detector (Agilent Technologies, Mississauga, Ontario, Canada).
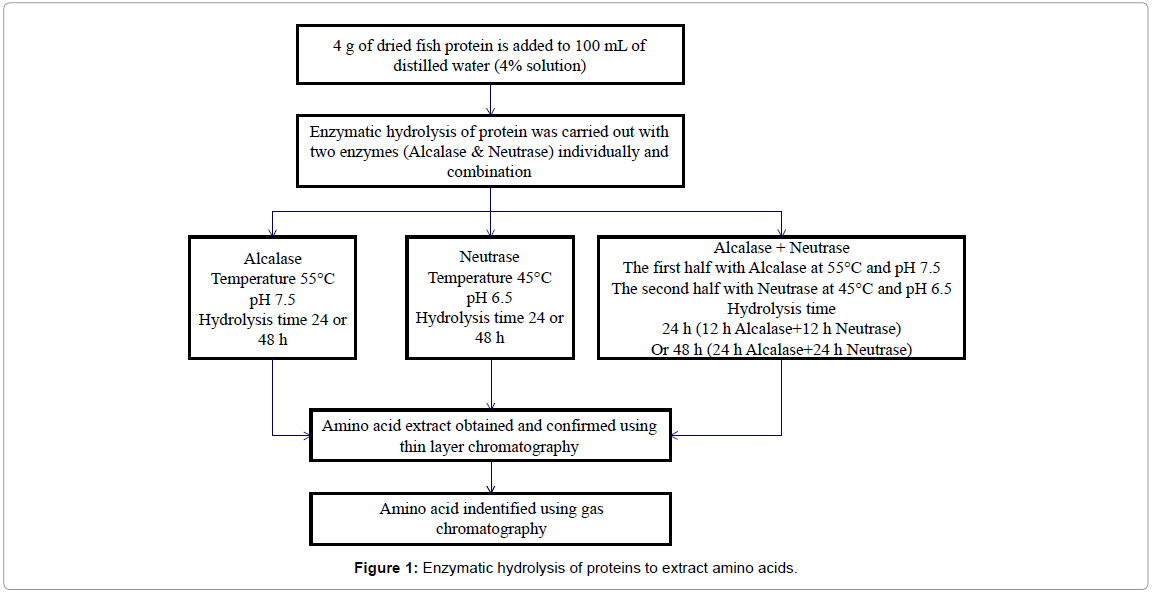
Figure 1: Enzymatic hydrolysis of proteins to extract amino acids.
Hydrolysis of protein with Neutrase was carried out using the same procedure, but the temperature and pH were adjusted to 45°C and 6.5, respectively. For hydrolysis of protein using the two enzymes in combination, the pH was adjusted to 7.5, and the sample was placed in a preheated (55°C) water bath and kept in the water bath for 30 min before adding the Alcalase enzyme (20 µL/g of protein). The hydrolysis was continued for 12 h and the pH were reduced from 7.5 to 6.5 using 1 N HCl, and the temperature of the water bath was reduced to 45°C, before adding the Neutrase enzyme. The hydrolysis was continued for another 12 h. The same procedure was repeated, but allowing 24 h for each enzyme.
Thin layer chromatography
Thin layer chromatography is the process of separating compounds based upon their differential affinity, in which the stationary phase is the thin layer of silica gel on a solid plate, and the mobile phase is the phase in which a small amount of sample spotted on the plate is carried through the stationary phase via capillary action. The procedures described by Srivastava et al. [17] were undertaken for the proper movement of the samples. A silica gel TLC plate (Sigma-Aldrich, Oakville, Ontario, Canada) was obtained, and a thin line was drawn 3 cm from the bottom and also from the top of the plate. On the bottom line, 21 evenly spaced markings were made for spotting the amino acid standards. All the amino acid standards were prepared in 10 ml test tubes in distilled water (concentration of 1mg/ml). The prepared amino acids standards were spotted on the markings using capillary tubes. The spotting of amino acids was carefully done, in such a way that the spots were small (bigger spots might lead to an imprecise result). After spotting, the plate was allowed to air dry. In the TLC tank, the solvent system consisting of butanol, glacial acetic acid and water in the ratio of 4:1:1 (the mobile phase) was prepared. The solvent system in the tank was exactly 1 cm from bottom of the tank. The TLC plate was then placed in the tank and was closed tightly. The solvent was allowed to run up to the top marked line on the plate. After that the TLC plate was carefully taken out, the line on the top of the sheet was the solvent front. The plate was allowed to completely dry, and then taken to the fume hood to spray 2% Ninhydrin in ethanol solution on it. It was then kept in a hot air oven (Isothermal Temperature 655 F oven, Fisher Scientific, Ottawa, Ontario) to dry at 100°C for 5 min. After drying, the spots were visible, and they were immediately circled and spotted in the center with a pencil. The Retardation factor (Rf) value of each amino acid was calculated using the following formula:
Rf=(Distance travelled by the spot)/(Distance travelled by the solvent) (1)
In a test tube, 100 µl from each amino acid standard solution was mixed together. To identify the amino acids present in a hydrolysed sample, the solution with the combined amino acids, and hydrolysed sample solution were spotted together on a same chromatogram plate and compared.
Gas chromatograph identification of amino acids
A Gas Chromatograph (Hewlett-Packard HP5890 Series II gas chromatograph, Agilent Technologies, Mississauga, Ontario, Canada) was used to identify and quantify each amino acid in the samples. Non-packed silica capillary column (Alltech Associates, Bannockburn, Illinois, USA), with intermediate polarity, with a length of 25 m and diameter of 0.54 mm was used. Helium was used as the carrier gas, and the stationary phase in the column was amino acids. The initial pressure was 34.47 kPa, the flow rate was 4 m/min and the velocity was 1cm/sec. A Flame Ionization Detector was used for the detection of amino acids. The rate of temperature for heating was set at 10°C/ min, to reach a final temperature of 210°C. The total time for each run was 60 min. The sample size injected into the chromatogram was 10 µl. An amino acid derivatization kit (Grace Discovery Sciences, Deerfield, Illinois, USA), in which 40 mg of the amino acids were placed in a small reaction vial, was used. 3 ml of 0.2 M HCl was added, and the solution was heated to 110°C for 5 min. Then, the vials were dried under a stream of dry nitrogen. Acetyl chloride was slowly added to isopropanol, and this mixture was added to the dried sample and heated to 100°C for 45 min. The vial was later uncapped and heated to approximately 115°C, under a stream of dry nitrogen to remove excess reagent. The vial was later cooled in an ice bath (-2°C) for 2 min. After cooling, 3 ml of methylene chloride and one ampoule of Penta Fluoro Propionic Anhydride (PFPA) were added to the vial, and the vial was reheated to 100°C for 15 min. The vial was again cooled to ambient temperature and dried under a stream of dry nitrogen, and dissolved in suitable solvent for analysis using the gas chromatograph.
Determination of amino acid yield
The yield of individual amino acid is the ratio of peak area of the corresponding amino acid to the total area count of the chromatograph. It was calculated as follows:
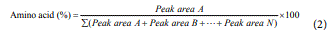
The hydrolysis of fish protein was performed to extract amino acids by the enzymatic breakdown of the peptide bonds in the proteins. The amino acids were identified by the Thin Layer Chromatography and quantified by Gas Chromatography.
Identification of amino acids by thin layer chromatography
Thin layer chromatography (TLC) was performed on Silica G TLC plates. The standard amino acids were first subjected to TLC, and the distance travelled by each amino acid was measured to calculate the retardation factor of each amino acid using Equation 1. The results are shown in Table 1. The standard amino acid chromatogram is shown in Figure 2.
| Amino acid | Distance travelled | Retardation factor | Retardation factor |
|---|---|---|---|
| By spot (cm) | By solvent (cm) | ||
| L-Alanine | 3.9 | 12.5 | 0.312 |
| L-Arginine-Hcl | 1.5 | 12.5 | 0.120 |
| L-Aspargine | 2.5 | 12.5 | 0.200 |
| L-Aspartic acid | 2.8 | 12.5 | 0.224 |
| L-Cysteine-Hcl | 2.9 | 12.5 | 0.232 |
| L-Cystine | 1.1 | 12.5 | 0.088 |
| L-Glutamic acid | 3.9 | 12.5 | 0.312 |
| L-Glutamine | 2.6 | 12.5 | 0.208 |
| L-Glycine | 3.1 | 12.5 | 0.248 |
| L-Histidine | 1.7 | 12.5 | 0.136 |
| L-Hydroxyproline | 2.9 | 12.5 | 0.232 |
| L-Isoleucine | 6.9 | 12.5 | 0.552 |
| L-Leucine | 7.2 | 12.5 | 0.576 |
| L-Lysine | 1.0 | 12.5 | 0.080 |
| L-Methionine | 5.9 | 12.5 | 0.472 |
| L-Phenylalanine | 7.0 | 12.5 | 0.560 |
| L-Proline | 2.5 | 12.5 | 0.200 |
| L-Serine | 3.2 | 12.5 | 0.256 |
| L-Threonine | 3.6 | 12.5 | 0.288 |
| L-Tryptophan | 7.0 | 12.5 | 0.560 |
| L-Tyrosine | 6.7 | 12.5 | 0.536 |
| L-Valine | 5.5 | 12.5 | 0.440 |
Table 1: Thin layer chromatograph of standard amino acids.

Figure 2: Thin layer chromatograph of standard amino acids (1: L-Valine; 2: L-Proline; 3: L-Glutamine; 4: L-Threonine; 5: L-Tryptophan; 6: L-Hydroxyproline; 7: L-Aspargine; 8: L-Histidine; 9: L-Cysteine-Hcl; 10: L-Cystine; 11: L-Alanine; 12: L-Arginine-Hcl; 13: L-Aspartic acid; 14: L-Tyrosine; 15: L-Glycine; 16: L-Isoleucine; 17: L-Phenylalanine; 18: L- Lysine; 19: L-Leucine; 20: L-Methionine; 21: L-Serine; 22: L-Glutamic acid).
The enzymatic hydrolysis of fish protein was carried out using Alcalase, Neutrase and combination of Alcalase and Neutrase for 24 and 48 h. After the hydrolysis process, the samples were first heated at 90°C for 5 min to deactivate the enzymes, and then allowed to cool down. The TLC plates were spotted with standard amino acids for identification. After completing the run, the plates were dried and sprayed with Ninhydrin to identify the presence of amino acids after hydrolysis. Figure 3 represents a chromatogram containing standard amino acids in the first and second spots. The third spot is the sample of hydrolyzed protein with Alcalase for 24 h, and the fourth spot is the sample hydrolyzed with Alcalase for 48 h. Figures 4 and 5 represent the chromatogram for identification of amino acids after hydrolysis for 24 or 48 h with Neutrase and combination of Alcalase and Neutrase, respectively. The differentiation in the spots was not clearly seen due to the streaking and spreading of spots, and single dimensional chromatography was not helpful in separation of amino acids.
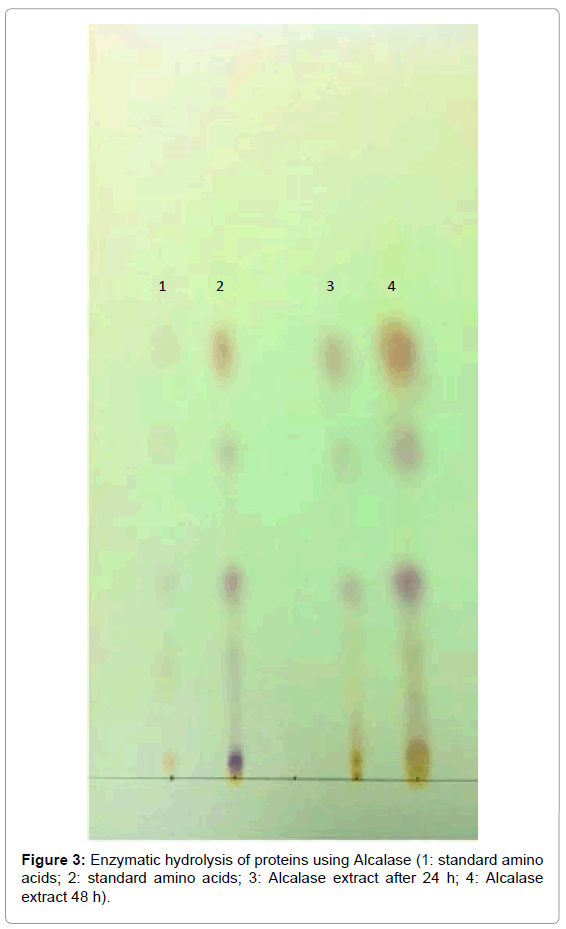
Figure 3: Enzymatic hydrolysis of proteins using Alcalase (1: standard amino acids; 2: standard amino acids; 3: Alcalase extract after 24 h; 4: Alcalase extract 48 h).
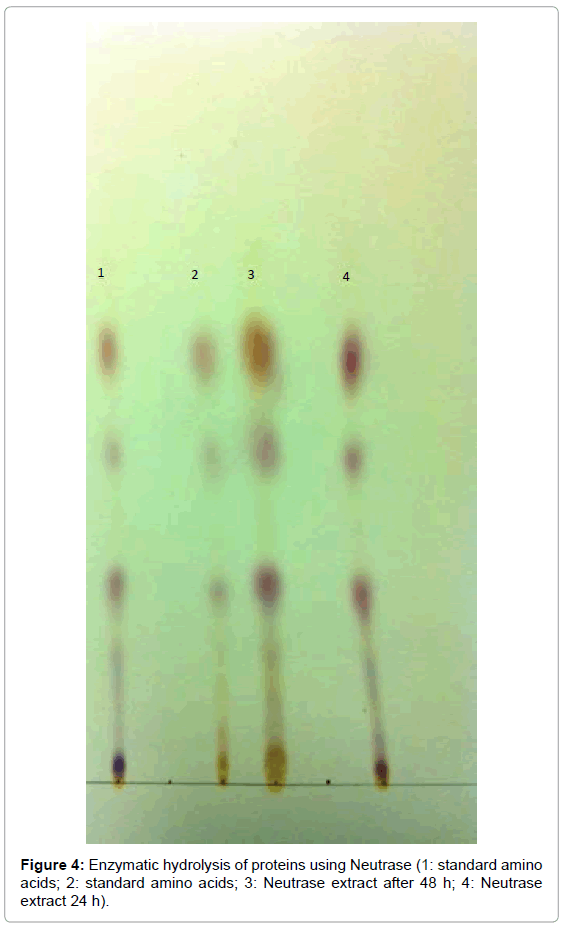
Figure 4: Enzymatic hydrolysis of proteins using Neutrase (1: standard aminoacids; 2: standard amino acids; 3: Neutrase extract after 48 h; 4: Neutrase extract 24 h).
Figure 5: Enzymatic hydrolysis of proteins using Combination of Neutrase and Alcalase (1: standard amino acids; 2: Alcalase+Neutrase-24 h; 3: Alcalase+Neutrase-48 h).
Quantification of amino acids by gas chromatography
The amino acids were quantified using gas chromatography. The proteolytic digestion of fish protein was conducted using Alcalase, Neutrase and combination of Alcalase and Neutrase for 24 and 48 h. At the end of hydrolysis, the samples were spray dried to obtain amino acid powder, and the amino acids were subjected to derivatization for gas chromatograph analysis. The results are shown in Table 2.
| Name | Weight Percent | |||||
|---|---|---|---|---|---|---|
| Alcalase | Neutrase | Combination | ||||
| 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | |
| Alanine | 1.63 ± 0.311 | 5.06 ± 0.28 | 1.29 ± 0.07 | 4.73 ± 0.24 | 4.62 ± 0.28 | 7.59 ± 0.24 |
| Glycine | 0.00 ± 0.00 | 2.99 ± 0.48 | 0.00 ± 0.00 | 2.35 ± 0.29 | 0.00 ± 0.00 | 5.82 ± 0.37 |
| Valine | 1.74 ± 0.24 | 4.04 ± 0.33 | 1.43 ± 0.29 | 3.67 ± 0.38 | 3.10 ± 0.30 | 7.20 ± 0.30 |
| Threonine | 0.00 ± 0.00 | 1.09 ± 0.21 | 0.00 ± 0.00 | 1.03 ± 0.32 | 1.08 ± 0.14 | 5.40 ± 0.51 |
| Serine | 0.89 ± 0.47 | 2.71 ± 0.07 | 0.80 ± 0.25 | 1.99 ± 0.22 | 0.86 ± 0.08 | 4.30 ± 0.48 |
| Leucine | 0.00 ± 0.00 | 4.49 ± 0.24 | 0.00 ± 0.00 | 3.32 ± 0.33 | 2.49 ± 0.28 | 9.00 ± 0.04 |
| Isoleucine | 0.00 ± 0.00 | 1.74 ± 0.41 | 0.00 ± 0.00 | 1.30 ± 0.48 | 3.12 ± 0.20 | 5.30 ± 0.16 |
| Proline | 0.00 ± 0.00 | 0.56 ± 0.11 | 0.00 ± 0.00 | 0.51 ± 0.24 | 0.00 ± 0.00 | 0.98 ± 0.17 |
| Phenylalanine | 0.00 ± 0.00 | 2.49 ± 0.39 | 0.00 ± 0.00 | 2.23 ± 0.31 | 2.12 ± 0.23 | 4.20 ± 0.31 |
| Methionine | 0.00 ± 0.00 | 1.79 ± 0.22 | 0.00 ± 0.00 | 0.00 ± 0.00 | 1.92 ± 0.31 | 2.20 ± 0.31 |
| Aspartic acid | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Glutamic acid | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.34 ± .0.13 | 9.85 ± 0.10 |
| Arginine | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Histidine | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 3.59 ± 0.38 |
| Lysine | 0.00 ± 0.00 | 4.36 ± 0.28 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.32 ± 1.41 | 7.34 ± 0.23 |
| Tyrosine | 0.00 ± 0.00 | 1.54 ± 0.29 | 0.00 ± 0.00 | 0.74 ± 0.28 | 0.00 ± 0.00 | 3.17 ± 0.11 |
Table 2: Amino acid composition from enzymatic hydrolysis using gas chromatography.
The results indicated that fourteen amino acids were quantified for the samples hydrolysed for 48 h, and only ten amino acids could be quantified for the samples hydrolysed using combination of enzymes for 24 h. Twelve amino acids were quantified individually using Alcalase for 48 h, whereas only three amino acids were quantified for the samples hydrolysed for 24 h. When the amino acids were hydrolysed using Neutrase for 48 h, ten amino acids were quantified, and only three amino acids were quantified when the samples were hydrolysed for 24 h. The yield of amino acids was the highest for the samples hydrolysed using a combination of enzymes for 48 h.
Amino acid extraction
Enzymatic hydrolysis of protein was carried out to extract amino acids for the production of jadomycin. Jadomycin is an antibiotic produced from Streptomyces venezuelae in three steps: growth, shocking and production. In the growth step, the Streptomyces venezuelae is grown in nutrient enriched Maltose-Yeast Extract Medium (MYM). In the shocking step, the grown inoculum is transferred to a nutrient deprived amino acid rich production media, and shocked with 3% (v/v) ethanol. In the production step, the production of jadomycin is induced after shocking, and amino acids are incorporated within the oxazolone ring structure. The presence of amino acids is an important ingredient for jadomycin production. Twelve types of jadomycin can be produced in the presence of different amino acids, including alanine, glycine, histidine, isoleucine, leucine, methionine, phenylalanine, serine, threonine, tryptophan, tyrosine and valine.
In this study, fourteen amino acids were identified and quantified after hydrolyzing the protein samples, using a combination of Alcalase and Neutrase enzymes for 48 h. High values of all the amino acids were obtained. Alcalase (endopeptidase and serine protease) and Neutrase (metallo endoprotease) was able to attack the protein backbone, and cleave the peptide bonds to release amino acids. However, these enzymes were not able to cleave some amino acids, such as arginine and aspartic acid. The yield of proline was also low, which is similar to those reported by Church et al. [12] and Hill and Schmidt [18].
Hill and Schmidt [18] hydrolyzed protein using several enzymes, including papain, carboxypeptidase, enolase, oxidized ribonuclease, leucine aminopeptidase, prolidase and a-corticotropin. Their results indicated that proteolytic hydrolysis of protein depended upon the properties and type of enzyme used in the process. Papain, being as endopeptidase, degraded most proteins to yield amino acids. Leucine aminopeptidase is one enzyme which can degrade proteins that do not contain proline and prolidase, and is capable of completely hydrolyzing proline. However, when carboxypeptidase was used together with aminopeptidase and prolidase, they did not have any significant effect on protein hydrolysis. The report suggested that the yield obtained using a combination of these enzymes was closely comparable to the yields obtained with acid hydrolysis.
D’Aniello et al. [19] hydrolysed proteins using acid (6 M HCl) at 37°C for 24 h, or at 90°C for 15 min, followed by enzymatic hydrolysis using pronase at 50°C for 24 h. The results suggested that during the initial chemical acid hydrolysis step, only 4-8% of proteins were hydrolysed into peptones, peptides and amino acids. The main advantage of carrying out the procedure at 37°C was that no racemization of amino acids occurs, and when the temperature was ramped up to 50°C for enzymatic hydrolysis, using pronase the rate of enzymatic hydrolysis was increased, and the enzyme was not deactivated as 90% hydrolysis of proteins was achieved.
Church et al. [12] hydrolysed proteins using immobilized pronase, proteinase K, carboxypeptidase A and B, aminopeptidase M, intestinal mucosa exopeptidase and prolidase for 18-24 h. The results suggested that use of enzymatic hydrolysis provided an alternative method to hydrolyze proteins, without addition of acid or alkali, and it did not cause racemization of amino acids. The results indicated that the rate of hydrolysis of pronase individually from 0-4 h were similar to the rate of combined exopeptidases, endopeptidase and neutral enzymes at the same time interval, but when time was increased from 4 to 24 h, the rate of hydrolysis drastically increased for the combination of enzymes, with 90% hydrolysis of proteins. The report also indicated that cysteine, cystine, arginine, proline, glutamine and aspargine bonds were most resistant towards proteolytic cleavage. Some disadvantages of enzymatic hydrolysis included multiple pH and temperature changes, if hydrolysis is extended more than 24 h.
In this study, the amino acids extracted from fish protein using two enzymes, Alcalase and Neutrase (individually and together in combination), were identified using single dimensional thin layer chromatography. The results suggested that changing the solvent system, and carrying out two dimensional thin layer chromatography, can help in defining the individual amino acids present in the sample. Bujard and Mauron [20], Bieleski and Turner [21], Jones and Heathcote [22], Ali et al. [23], and Matysik and Soczewinski [24] stated that thin layer chromatography is a simple and rapid analytical method for the identification of amino acids. It is, however, a qualitative analysis of amino acids, in which individual amino acids cannot be well differentiated due to streaking and spreading of spots, and the Rf values are not reproducible.
Stroud [25] reported that the bond cut by the serine proteases is the one that joins amino acids together to form proteins. Each amino acid is composed of an amino group (NH2) and a carboxyl group (COOH) attached to a single alpha carbon atom. The carbon atom is also attached to a hydrogen atom, and one of the twenty side chains by which the amino acids are identified. During synthesis of proteins, the carboxyl group of one amino acid is connected to the next amino group by the extraction of water molecule to form a linkage -CO-NH-. (Peptide bond). During the hydrolysis of proteins, a water molecule consisting of a hydrogen atom and a hydroxyl group is added for each peptide bond broken, thereby restoring the amino and carboxyl group at the site of cleavage to their free amino acid form.
In this study, Alcalase being a serine protease belonging to subtilisin family and Neutrase being a metallo endoprotease, contains three residues critical for catalysis: a serine, a histidine, and an aspartic acid[26,27]. These three residues break the peptide bond in the protein backbone and release amino acid in the system. The specific mechanism for Alcalase and Neutrase breaking the peptide bond has not been reported in the literature. Bachovchin and Roberts [28] and Brand [29] reported a hydrolysis mechanism for chymotrypsin (Figure 6), which is also a serine protease containing three residues, including: a serine, a histidine, and an aspartic acid for the catalysis of peptide bond. Therefore, both Alcalase and Neutrase used in this study are suggested to follow the same mechanism.
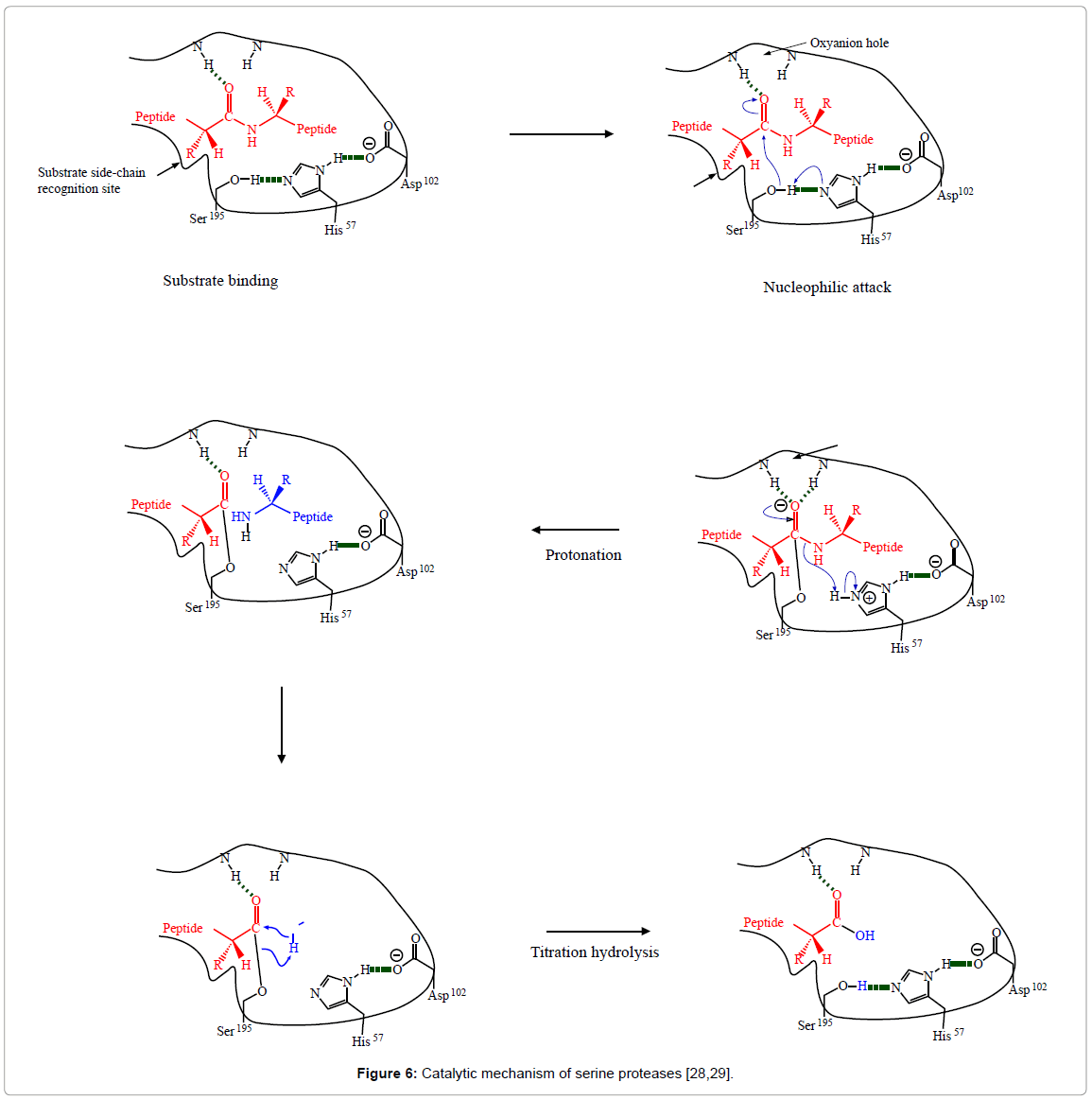
Figure 6: Catalytic mechanism of serine proteases [28,29].
Marquez and Vazquez [30] and Sousa et al. [31] reported that four steps are involved during the general proteolytic cleavage of proteins, which include substrate binding, opening of the peptide bond, proton exchange and titration of amino group. During the hydrolysis of the proteins, the serine 195 pivots approach and attack the carbonyl carbon (-CO-) of the substrate group, at the same time, the hydrogen ion or proton of the hydroxyl group is transferred to the nearby histidine 57, and the serine hydroxyl oxygen forms a covalent bond with the carbonyl carbon. As a result, the double bond connecting the carbon and oxygen in the substrate carbonyl group is transformed into a single bond. During protonation, the proton is delivered to histidine 57, and remains there for a short time and then gets transferred to the -NH- group in the substrate on the other side of the peptide bond that is being broken. In the final step, water attacks the ester bond between the peptide and the serine 195 oxygen, and forms the second product peptide with normal carboxyl group, and the regenerated the serine hydroxyl group. The second peptide is then dissociated from the enzyme to allow another catalytic cycle to begin.
Jadomycin production
The aim of this study was to extract amino acids for use in jadomycin production. Burdock et al. [2] reported that a concentration of 4.5 g/L (0.45%) of individual amino acid is required for the production of jadomycin. Out of fourteen amino acids produced in this study (Table 3), twelve amino acids were present in relatively reasonable concentrations suitable of producing jadomycin. These were alanine (7.59%), glycine (5.82%), histidine (3.59%), isoleucine (5.30%), leucine (9%), lysine (7.34%), methionine (2.2%), phenylalanine (4.2%), serine (4.3%), threonine (5.40%), tyrosine (3.17%) and valine (7.2%).
| Suitable amino acids for jadomycin production | Jadomycin | Extracted amino acids | ||
|---|---|---|---|---|
| (wt %) | Suitability | |||
| Alanine | Jadomycin Ala | 7.59 | ✓ | |
| Glycine | Jadomycin G | 5.82 | ✓ | |
| Histidine | Jadomycin H | 3.59 | ✓ | |
| Isoleucine | Jadomycin B | 5.30 | ✓ | |
| Leucine | Jadomycin L | 9.00 | ✓ | |
| Lysine | Jadomycin K | 7.34 | ✓ | |
| Methionine | Jadomycin M | 2.20 | ✓ | |
| Phenylalanine | Jadomycin F | 4.20 | ✓ | |
| Serine | Jadomycin S | 4.30 | ✓ | |
| Threonine | Jadomycin T | 5.40 | ✓ | |
| Tryptophan | Jadomycin W | 0.00 | x | |
| Tyrosine | Jadomycin Y | 3.17 | ✓ | |
| Valine | Jadomycin V | 7.20 | ✓ | |
* A minimum concentration of 0.45% was considered for economic reasons [2].
Table 3: Jadomycin produced by S. venezuelae ISP5230 by ethanol shock [34,35].
However, tryptophan, which is suitable for producing the jadomycin W was not present in the fish protein. Leu et al. [32] extracted sixteen amino acids from the Atlantic Mackerel (Scomber scombrus), and reported that the amino acid tryptophan was not present in the fish. Doull et al. [33] reported that when glutamic acid was used in the production medium, the color of the medium was orange and lysine, indicating the possibility of jadomycin production. No other reports were found in the literature on jadomycin production from glutamic acid. Therefore, the possibility of producing jadomycins from glutamic acid (9.85%), and proline (0.98%) which were present in the fish protein should be investigated.
Doull et al. [33], Jakeman et al. [1], Borissow et al. [34] and Burdock et al. [2] produced several jadomycins using Streptomyces venezuelae ISP5230, with individual amino acids in the production medium. No reports were found in the literature to suggest the use of more than one amino acid in the same production medium. Therefore, the amino acids extracted from fish protein in this study have to be separated and purified before use for the production of individual Jadomycins [35]. However, the possibility of producing multiple jadomycins using several amino acids in the production medium should also be investigated.
The effect of two types of enzymes (Alcalase and Neutrase), individually and together in combination, and hydrolysis time (24 and 48 h) on the extraction of amino acids from fish protein were studied. During protein hydrolysis, the enzyme Alcalase (being a serine endopeptidase) and the enzyme Neutrase (being a neutral metallo endoprotease) act upon the peptide bonds to release amino acids into the system. The highest amount of amino acids were obtained from the samples hydrolyzed, using a combination of enzymes (Alcalase+Neutrase) for 48 h. Fourteen amino acids were identified. These were: alanine (7.59%), glycine (5.82%), histidine (3.59%), Isoleucine (5.30%), leucine (9%), lysine (7.34%), methionine (2.2%), phenylalanine (4.2%), serine (4.3%), threonine (5.40%), tyrosine (3.17%), valine (7.2%), glutamic acid (9.85%) and proline (0.98%). Two amino acids (arginine and aspartic acid) could not be quantified, as the enzymes were not able to cleave them. All the amino acids, suitable for the jadomycin production, can be obtained from enzymatic hydrolysis of fish proteins. All the amino acids were above the minimum concentration of 0.45% for the jadomycin production. Tryptophan which is suitable of producing jadomycin was not present in the fish protein. The feasibility of jadomycin production from glutamic acid and proline should be investigated. Currently, jadomycins are produced individually, using only one amino acid in the medium, and as such the amino acids extracted from fish protein have to be separated and purified before use for production of individual jadomyicns. The possibility of multiple production of jadomycins in the same medium containing the mixture of amino acids has to be investigated.
The research was supported by the Natural Sciences and Engineering Research Council (NSERC) of Canada.