Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Commentry - (2017) Volume 8, Issue 1
Early fingerlings of Anabas testudineus were nurtured in the laboratory for six weeks under altered nutritional status (different dietary protein level and source) and feeding regimes (ad libitum, ‘limited’, and ‘starvation’ intervals). Their RNA/DNA, Ca/P ratios and protein biosynthesis were determined with highly sensitive fluorescence technique, spectrophotometer and liquid scintillation system respectively. Well-fed early fingerlings (4-6 weeks age group) having higher RNA/DNA (2.11), Ca/P (1.82) ratio and protein biosynthesis rates both in muscle and liver have significantly differs from fingerlings with ‘limited’ food supply and 'starvation' intervals. Rate of protein biosynthesis >244.33 & 603.24 in muscle & liver respectively (count per minute mg protein-1) when compared with well-fed fingerlings and reared under 'limited' food supply and 'starvation' intervals. At ‘starvation’ interval no significant differences were recorded due to different diet treatment. However, fingerlings showed higher mortalities, decreased growth, RNA/DNA, Ca/P ratio and protein biosynthesis rate that may be caused by malnutrition. Fingerlings (early) sampled after 42 days in nearby areas during late monsoon (September) showed RNA/DNA, Ca/P ratio and protein biosynthesis rate were surprisingly low almost as low as ‘starved’ fingerlings, when judged on the basis of mean body weight as compared to well fed fish. It was also observed under microscope that some of these fingerlings were seriously affected by an intestinal disease and this may be due to the ingestion of unsuitable copepods. The low RNA/DNA, Ca/P ratio and protein biosynthesis rate in field samples and ‘starved’ fish are, therefore, interpreted as a sub-lethal result of this disease and malnutrition respectively.
Keywords: Anabas testudineus; Intestinal disease; Nutritional condition; Ca/P; Protein bio-synthesis; RNA/DNA
In intensive and semi-intensive aquaculture practices, production of specific fish can only be economical when its qualitative and quantitative feed requirements are known [1]. Since protein is considered to be one of the most important and expensive nutrients in any feed, so that its inclusion in formulated diets should be approached with care. Although protein requirements of cultivable warm water fishes have been already studied by many workers [2-6] still yet not much information is available on the influence of dietary protein level, isonitrogenous diets with variable protein source and different feeding regimes (ad libitum, ‘limited’, and ‘starvation’ interval) on the growth performance, RNA/DNA ratio, Ca/P ratios and the rates of protein bio-synthesis in the muscle and liver of the Indian climbing perch, Anabas testudineus . Therefore, an effort has been made to determine the correlation with the growth and RNA/DNA, Ca/P ratio and protein bio-synthesis rate.
It is generally accepted that fish stock size are mainly determined during early life history. Survival of young fish can be controlled both by predation and starvation, and by the interaction of these two factors, because starving fish larva/fry are more vulnerable to predation [7]. "The nutritional condition of larval/fry fish, which is the key factor in this context, can be characterized histologically [8] by measuring proteolytic enzymes [9] or the RNA/DNA ratio [10-15]."
The RNA concentration is a sensitive parameter to determine the growth rate of an organism because it is the organizer of protein synthesis [16]. DNA concentration represents an index of cell number since cellular DNA content is sensitive to changes in environmental conditions [17]. The ratio of RNA/DNA is, therefore, a more accurate index of metabolic activity than RNA concentration alone, because this ratio is not affected by the number or size of the cells in tissue sample [18]. Similarly Ca/P ratio or protein biosynthesis rates also may be used as an index of growth [1] or the status of the nutritional condition.
Indian climbing perch (Anabas testudineus ) fingerlings were reared in the laboratory under different feeding conditions. Their RNA/DNA, Ca/P ratio and protein biosynthesis rate were taken as reference values in an attempt to characterize the nutritional status of Anabas fingerlings (early) sampled in the ponds of nearby areas.
The present study reports on the nutritional condition of A. testudineus with the help of RNA/DNA, Ca/P ratio and protein biosynthesis rate in the laboratory cultured fish with different diet treatments and field samples from natural waters of adjoining areas.
Aquaria experiments
Ten formulated diets were prepared in pelleted form of which ED01-ED05 at different protein level, containing approximately 20.0%, 25.0%, 30.0%, 35.0%, and 40.0% crude protein level respectively and is nitrogenous diets ED06-ED10 (Crude protein level ≅ 35.0%) containing soybean meal, fish meal, goat blood, silkworm pupae and carcass waste respectively as major protein source (Table 1). The proximate composition of the experimental diets was analysed following the standard methods of AOAC [19].
| Ingredients | Halver’s synthetic diet | Different dietary protein level | Isonitrogenous with different dietary protein source | ||||||||
| ED01 | ED02 | ED03 | ED04 | ED05 | ED06 | ED07 | ED08 | ED09 | ED10 | ||
| Mustard oil cake | - | 2.55 | 17.85 | 35.68 | 56.09 | 49.2 | 30.58 | 7.65 | - | - | - |
| Line seed oil cake | - | - | - | - | - | - | - | - | - | 31.56 | - |
| Ground nut oil cake | - | - | - | - | - | - | - | - | 35 | - | 24.79 |
| Wheat bran | - | 66.65 | 51.35 | 33.52 | 13.11 | - | 17.68 | - | - | 13.71 | - |
| Rice bran | - | - | - | - | - | - | - | 11.55 | 29.2 | 19 | 37.09 |
| Soybean meal | - | - | - | - | - | - | 50.94 | - | - | - | - |
| Goat blood | - | - | - | - | - | - | - | - | 35 | - | - |
| Silkworm pupae | - | - | - | - | - | - | - | - | - | 34.93 | - |
| Fish meal | - | 30 | 30 | 30 | 30 | 50 | - | 80 | - | - | - |
| Carcass waste | - | - | - | - | - | - | - | - | - | - | 37.32 |
| Casein | 38 | - | - | - | - | - | - | - | - | - | - |
| (Vit. Free) | |||||||||||
| Gelatin | 12 | - | - | - | - | - | - | - | - | - | - |
| Corn oil | 6 | - | - | - | - | - | - | - | - | - | - |
| Cod liver oil | 3 | - | - | - | - | - | - | - | - | - | - |
| White dextrin | 28 | - | - | - | - | - | - | - | - | - | - |
| a-cellulose | 10 | - | - | - | - | - | - | - | - | - | - |
| Premix2 | 3 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| 1Each diet contains 1 g% chitin as an internal marker (HROM). 2Vitamin and mineral mixture (vitaminate forte; Roche India Ltd). [Each 0.8 g contains Vit. A.I.P. (as Acetate) 2500 IU; Thiamine mononitrate I.P. (Vit.B1) 2.00 mg; Riboflavine I.P.(Vit.B2) 3.00 mg; Nicotinamide I.P. 25.00 mg; Pyridoxine hydrochloride I.P. (Vit.B6) 1.50 mg; Calcium pantothenate U.S.P. 5.00 mg; Cyanocobalamine I.P. Vit.B12 1.00 mcg; Ascorbic acid I.P. (Vit. C) 50.00 mg; Cholecalciferol U.S.P. (Vit.D3) 200 IU; Vit. ENF (as dl-alpha-tocopheryl acetate) 10.00 mg; Biotin (Vit.H) 0.05 mg; Calcium phosphate I.P. 208.00 mg; Dried ferrous sulphate I.P. 10.62 mg; magnesium phosphate, dibasic 48.00 mg; Manganese hypophosphite 0.60 mg; total phosphorus in the preparation 44.60 mg]. |
|||||||||||
Table 1: Proportion [% dry weight] of different ingredients in the prepared diets1, control [Halver’s synthetic diet] and experimental diets [ED01-ED10].
Two successive series of experiments were run. One batch with the varying levels of protein diet and another with different dietary protein source. Live healthy specimens of the order perciformes A. testudineus (4-6 weeks after hatching; average weight 3.28-3.97g and length 40-45 mm) were kept in separate specially designed continues flow fiber glass aquaria with aeration, temperature, photoperiod and water current control facility (size: 76.0 cm × 40.0 cm × 35.0 cm) of 100 liter water holding capacity in a batch of 20 fishes, after acclimatization of 15 days in the laboratory condition. A concurrent control batch of fish was maintained with Halver's synthetic diet. Each set of experiment was triplicate (Table 2).
| Proximate composition (%) | Diets | ||||||||||
| ED10 | |||||||||||
| Halver's diet | ED01 | ED02 | ED03 | ED04 | ED05 | ED06 | ED07 | ED08 | ED09 | ||
| Moisture | 66.65 | 6.72 | 6.38 | 5.85 | 5.26 | 4.12 | 5.27 | 3.34 | 5.41 | 5.17 | 3.72 |
| Dry matter | 33.35 | 93.28 | 93.62 | 94.15 | 94.74 | 95.88 | 94.73 | 96.66 | 94.59 | 94.83 | 96.28 |
| Ash | 5.05 | 9.99 | 10.56 | 11.24 | 12 | 15.2 | 5.95 | 18.78 | 10.8 | 9.8 | 16.09 |
| Organic matter | 94.95 | 90.01 | 89.44 | 86.76 | 88 | 84.8 | 94.05 | 81.22 | 89.2 | 90.2 | 83.91 |
| Crude protein | 45.62 | 21.45 | 25.42 | 30.04 | 35.34 | 38.63 | 34.68 | 35.46 | 34.81 | 34.3 | 34.83 |
| Crude lipid | 9.1 | 3.28 | 5.84 | 7.46 | 9.3 | 9.38 | 13.6 | 6.68 | 5.65 | 13.09 | 12.8 |
| Crude fibre | 10 | 5.73 | 5.67 | 5.61 | 5.53 | 4.51 | 6.59 | 3.09 | 10.38 | 10.37 | 13.13 |
| Nitrogen free extract | 30.23 | 50.73 | 45.3 | 38.98 | 31.76 | 27.36 | 32.91 | 32.65 | 32.94 | 27.26 | 19.43 |
| Calcium | 1.95 | 4.05 | 4.27 | 4.51 | 4.8 | 6.33 | 1.62 | 8.17 | 2.29 | 1.7 | 3.27 |
| Phosphorus | 0.99 | 0.3 | 0.45 | 0.62 | 0.81 | 0.86 | 0.96 | 0.82 | 0.8 | 0.51 | 1.17 |
| Gross energy (K cal g-1) | 5.087 | 3.835 | 4.077 | 4.23 | 4.401 | 4.374 | 4.905 | 4.099 | 4.276 | 4.718 | 4.51 |
| P/E ratio (mg protein/K Cal) | 89.68 | 55.93 | 62.35 | 71.02 | 80.29 | 58.32 | 70.7 | 86.51 | 81.41 | 72.7 | 77.22 |
| 1Each data is a mean of 10 separate determinations. | |||||||||||
Table 2: Estimated proximate composition percent1 of the prepared diets [Halver's synthetic diet and experimental diets ED01-ED10] on dry matter basis.
At each series of experiment three feeding regimes were established: (I) food was given three or four times a day in excess (ad libitum ); (II) food was given only once a day and two hours later the uneaten feed was drained out (‘limited’); (III) feeding was similarly to the second group, but after every 8-10 days of feeding fingerlings were exposed to a starvation interval of 4-5 days (‘starved’) [20]. Aquaria were cleaned daily and extra feed or faecal matter were removed up to an age of 6 weeks (after collection and acclimatization), samples of living fingerlings were taken at every 14 days for further analyses. In the case of ‘starved’ larvae samples were taken following the starvation interval. Larvae were killed with TCMP (Tri-chloromethyl-propanole, 1.5 gl-1), examined under a microscope for fullness of the gut and checked for possible damage. Weights were measured to the nearest 0.01 g.
Field sampling
During July, August, September, fingerlings of A. testudineus were sampled in the ponds of adjoining areas. They were caught with a conical net of 1.0 m diameter and 2.0 m in length, with mess sizes of 1500 μm near the aperture and 900 μm in the rear part. Surface trawls were made near the bank of the pond (shore area) at about 1.0 ms-1. After several trawls of 10-12 minute each, sampled fry were killed with TCMP, measured for weight, and stored in ice bucket until arrival at the laboratory. Some of the fry were taken into the laboratory in live condition through bucket for protein biosynthesis study.
Digestibility estimation
The digestibility of the nutrient in the feed was estimated by using the following equation:
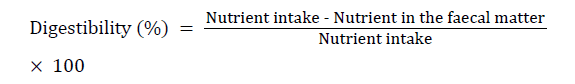
1.0 g percent chitin [hydrolysis resistant organic material, (HROM)] was used in 10 formulated diet (ED 01-ED10) and Halver’s synthetic diet as an internal marker that are usually being used in the digestibility estimation [21-27]. The different steps involved for the estimation are as follows:
Nutritive value (Feed utilization)
Food conversion ratio (FCR) and protein efficiency ratio (PER) were calculated following the formula as described by
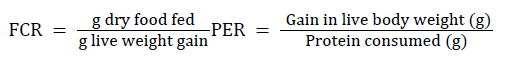
Extraction and estimation of nucleic acids
The extraction procedure developed by Clemmesen for marine fish larvae was adapted to the higher dry weight of coregonid larvae (0.6-10 mg) [12]. By means of teflon pistle individual fingerlings were homogenized for 1 minute on ice in 1.0 ml STE buffer (0.05M Tris-HCl buffer containing 0.1M NaCl and 0.01M EDTA) at pH 7.5, contained 0.2 mg ml-1 proteinase K. All further steps were carries out at room temperature. After adding 100 μl of 20% Sodium dodecyl sulphate (SDS) the sample shaken for 15 minute and centrifuged at 8000 g for 15 minutes. From the supernatant, 0.5 ml was transferred to a new 1.5 ml vial and 0.5 ml of a 1:1 mixture of phenol-8-hydroxyquinoline (1:0.001) and chloroform-isoamyl alcohol (24:1) was added. After centrifuging for 10 minutes each, the aqueous phase was transferred to a new vial and phenol-chloroform phase was discarded. The extraction was carried out twice. To remove phenol residues from the aqueous solution, the sample was washed twice with chloroform-isoamyl alcohol by shaking for 5 minutes, followed by 5 minutes of centrifugation. Finally, two 0.1 ml aliquots of the purified nucleic acid solution were transferred to sterilized test tube containing 2.5 ml of STE buffer at pH 7.5. Two specific fluorophors which react with nucleic acids were used. Ethidium bromide (EB) was used to determine DNA and RNA simultaneously, Bisbenzimidazole (BB) was used to determine DNA alone (Clemmesen 1988). The EB-fluorescence was determine at 365 nm excitation and 590 nm emission wave length, the BB-fluorescence at 352 nm excitation and 448 nm emission wave length [28].
Estimation of Calcium and Phosphorus
The amount of calcium (Ca) present either in the feed or in the muscle was estimated titrimetrically following the Clark-Collip modification of Kramer-Tisdall method as described by Oser. 2.0 g sample (feed/muscle) was made into ash in a Muffle furnace at 550°C ± 50°C, dissolved in 20.0 ml of 6N HCl, boiled and filtered through Whatmann filter paper No. 42. 2.0 ml filtrate was diluted with 98.0 ml of distilled water and added 2 drops of methyl red indicator. Then diluted ammonium hydroxide (NH4OH) was added drop wise till the colour changes to brownish orange. Subsequently, 25% hydrochloric acid (HCl) was added to have a pink colour, entire mixture diluted to 150 ml and after boiling, 10.0 ml of hot saturated ammonium oxalate solution was added. Next day after filtered through ash less filter paper, the precipitate washed with distilled water (150 ml) and subsequently 5.0 ml of concentrated H2SO4 was added. After heating, titrated with N/10 Potassium permanganate (KMnO4) to the first appearance of pink colour. The calcium content was calculated as:
Ca (%) = Amount of titrant 0.02 × 10 × 50
The phosphorus content was estimated following the methods of Fiske and Subba Row as described by Oser (1960). 2.0 g sample was made into ash, 70.0 ml of distilled water and 10.0 ml of ammonium molybdate solution was then added. After mixing, 4.0 ml of 1,2,4- aminonapathosulphonic acid was added to it and made upto 100 ml with distilled water. The optical density (OD) of the standard and unknown was determined in a spectrophotometer (Systronics-106) at 660-720 nm. The amount of phosphorus present in the feed or muscle was calculated as:

Protein biosynthesis study
The rate of protein synthesis in the liver and muscle of the experimental fish were measured as incorporation of amino acid (14C L-leucine) in the liver and muscle [29]. The fish from each of the experimental set were injected intramuscularly with the isotope 14C Lleucine (specific activity 282 Mci MMOL-1 ; solution in 0.01N HCl; obtained from Bhaba Atomic Research Centre, Trombay, Mumbai-400 085) both at the initial stage and at the termination of six weeks experimental period at the rate of 0.05 μCi 100g-1 body weight of the fish. The injected fish were sacrificed to measure the level of incorporation. The liver and muscle tissues were weighed and processed with trichloroacetic acid (TCA) extraction for protein. The final precipitate was dissolved in 0.5 ml 1.0% NaOH solution, boiled and mixed well. 7.5 ml of Scintillation fluid was added in the solution and radioactivity was counted in an Automatic Liquid Scintillation System (EC LSS 20). Protein estimation was made following the standard method of Lowry [30].
Monitoring of water quality
The water quality of the experimental aquaria was monitored periodically (once in a week following the methods of APHA [31]. The water temperature during the experimental period ranged from 29.0°C to 30.5°C. The pH, dissolved oxygen content and alkalinity of the water ranged between 6.9 and 7.1, 2.9 and 6.1 mg liter-1 and 93.95 and 115.05 mg liter-1, respectively.
Statistical analysis
All calculations and statistical analysis for comparing the data (level of significance) were done on Intel P-III PC 650 using statistical packages SPSS, STATISTICA, EXCEL etc.
The growth of the fish was positively correlated with the increase in the dietary protein level. The gain in body weight and feed intake were significantly (P<0.001) higher with ED01-ED05 when compared with the well fed fish (ad libitum ) and the fish with ‘limited’ food and ‘starvation’ interval. Similar observations have been reported by Austreng and Refstie in rainbow trout, Viola and Zohar in market size hybrids of Tilapia (Oreochromis sp .), Machiels and Henken in Clarias gariepinus , Patra in Clarias batrachus and in Anabas testudineus [5,15,32,33]. Results also indicate that the weight gain with 40% protein diet (ED05) in not significantly higher than the fish fed with the diet having 35% crude protein level (ED04). Although it is significantly higher (P<0.01) in case of ED05 when the fish provided with limited food and starvation interval (Figure 1a and 1b). It is possible therefore, that comparatively higher lipid (9.30% in ED04 and 9.38% in ED05) and nitrogen free-extract (31.76% in ED04 and 37.36% in ED05) might have influenced the growth of the fish fed on these diets as the sparing action of dietary lipid and carbohydrate on dietary protein level is well documented [34,35].

Figure 1: Different steps involved for the estimation.
The gain in body weight was positively affected by the different dietary protein source (ED06-ED10) and were maximum with the carcass waste (ED10) followed by silk worm pupae (ED09), goat blood (ED08), fish meal (ED07) and soybean meal (ED06) although there is no significant differences were found with the diet ED08 and ED09. But the growth increment was significantly lower (P<0.001) in all the diets (ED06-ED10) when the fish were maintained with ‘limited’ food and ‘starvation’ interval. The soybean meal diet shows lowest growth may be due to the total plant protein diet. The same phenomenon (showed in Figure 2a and 2b) has been reported with Plaice and Channel catfish [36].
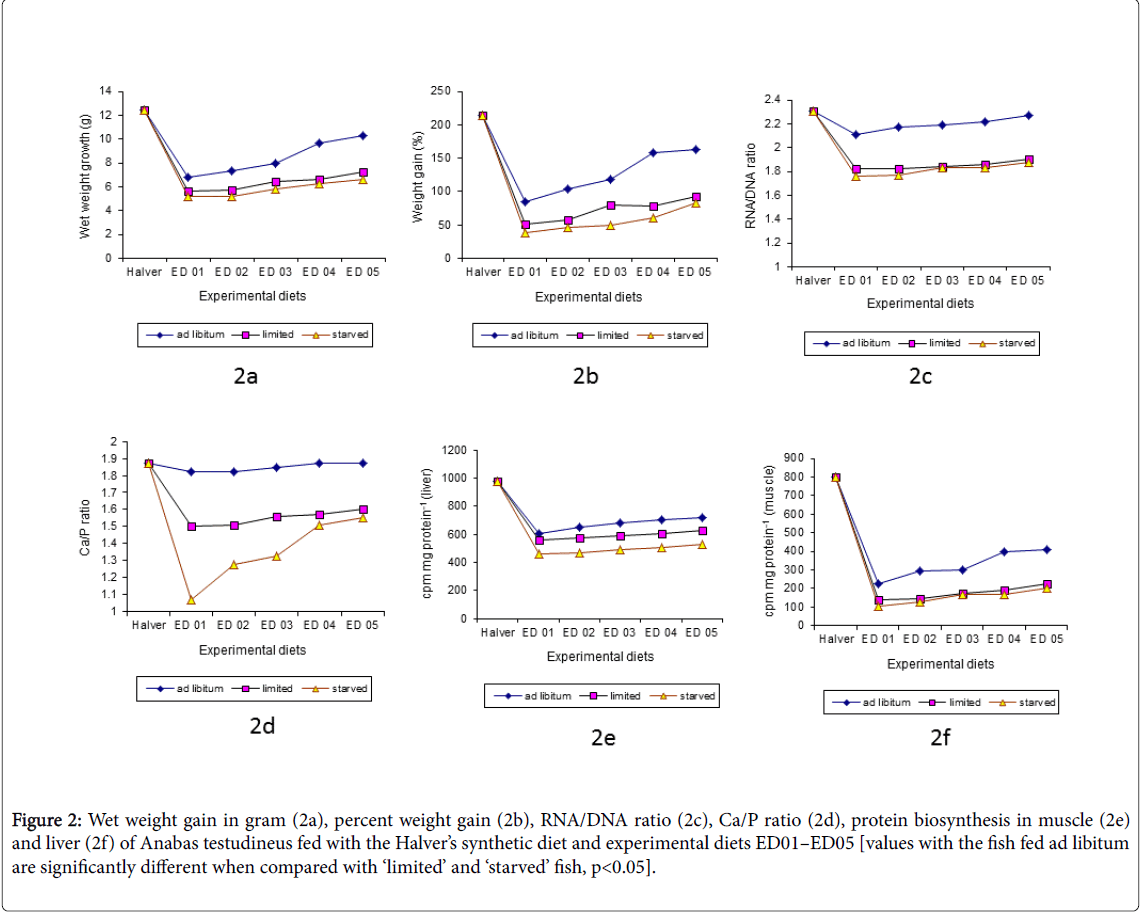
Figure 2: Wet weight gain in gram (2a), percent weight gain (2b), RNA/DNA ratio (2c), Ca/P ratio (2d), protein biosynthesis in muscle (2e) and liver (2f) of Anabas testudineus fed with the Halver’s synthetic diet and experimental diets ED01–ED05 [values with the fish fed ad libitum are significantly different when compared with ‘limited’ and ‘starved’ fish, p<0.05].
In general the digestibility of protein, lipid and energy were found to be positively correlated with the crude protein level of the diets (ED01- ED05), similar to the findings of Austreng and Refstie (Austreng and Refstie and Patra. With the diet ED06-ED10 the digestibility of protein, lipid and energy were varied with the different dietary protein source and maximum with the soybean meal (ED06) followed by silk worm pupae (ED09), carcass waste (ED10), goat blood (ED08) and fish meal (ED07). Although the digestibility of protein, lipid and energy were slightly higher in all the treatments with the diet ED01-ED10 when food is limited and there are starvation intervals. Digestibility of CP and CL were significantly (P<0.05) greater when equated between well fed fish and starved fish [37].
The results on nitrogen balance indicate an increase in N intake and faecal N excretion with corresponding increase in the dietary protein levels as has also been reported by Rychly in rainbow trout and Patra in Indian climbing perch and it is variable with the dietary protein source [5,38]. Like nitrogen, the apparent absorbed feed energies also increased with the protein level and varied with the protein source. Nitrogen and energy intake and absorption were significantly higher (p<0.05) when compared to the ad libitum fish with ‘limited’ and ‘starved’ fish, although it was not significant when compared between ‘limited’ and ‘starved’ [34].
It also observed an increase in FCR value with the increasing dietary protein levels (Beamish and Medland 1986). But in the present study , FCR value was found to increase in 30% protein level (ED03) as compared with the higher protein level diet (ED04, ED05) and variable in performance and highest with carcass waste (ED10) followed by goat blood (ED08), fish meal (ED07), silk worm pupae (ED09) and soybean meal (ED06). It is lowest in the Halver's synthetic diet (control) and higher when the fish provided with the ‘limited’ food. The results indicate negative correlation between the dietary protein levels and PER values and variable with the different dietary protein source as has been noted by Kim in the fish studied by them. Significantly higher (p<0.05) PER value has been recorded in this study when ‘starved’ fish compared with ad libitum and ‘limited’ fish [39].
Cumulative mortalities and sample means of live weight gain, percent weight gain, RNA/DNA, Ca/P ratio and protein biosynthesis rate in the liver and muscle are shown in Figure 1a-1f and 2a-2f. In the first series of experiment, fish fed ad libitum , a significant increase (‘t’ value, p<0.05) in the levels of RNA/DNA ratio in the muscle were noticed in the fish fed the diets ED01-ED05 as compared to the fish fed ‘limited’ and with ‘starvation’ intervals and also the fish sampled from nearby areas of same size groups (Figure 1c). The second series of experiment with the diet ED06-ED10 RNA/DNA ratios (‘t’ value, p<0.05) showed similar trend as observed in the first series (Figure 2c). The sampled fish showed even lower RNA/DNA value than ‘limited’ and ‘starved’ fish of the first and the second series of experiment. In laboratory reared healthy fingerlings, the differences between well fed specimens and those which were reared under restricted food supply were evident at the different dietary protein level and source: fingerlings with RNA/DNA ratios around 2.3 and lower must be regarded as under nourished. Although differences were detected either in the different protein level or variable dietary protein source, ‘starved’ fish showed lowest RNA/DNA ratio when compared with ad libitum and ‘limited’. Furthermore, it was not possible to distinguish significantly between fingerlings which were given a ‘limited’ ration and those which were exposed to intermittent ‘starvation’ intervals. This may be caused by the short length of the starvation intervals. Longer starvation intervals might lead to even lower RNA/DNA ratio. Nevertheless, either in different protein level or variable protein source, mortality was clearly higher in ‘starved’ fingerlings than in those fingerlings on ‘limited’ ration.
A critical RNA/DNA ratio of 2.5 was also found for herring larvae (Clupea harengus L .) older than 10 days. In yolk sac herring larvae, however, no clear separation was possible between starved and fed specimens [28]. Study also found that increasing RNA/DNA ratio even in starved herring larvae until the stage of yolk sac stage. The subsequent rise of the RNA/DNA ratio with age in well-fed Anabas fingerling has also been observed in striped bass (Morone saxalitis , Walbaum) larvae and in herring larvae (Clemmesen, 1989), while in winter flounder (Pseudopleunectes americanus , Walbaum) did not find any dependence on larval age [10,11]. The application of RNA/DNA method to field samples is only possible with certain precautions. Knocking the age of individual fingerlings would aid considerably in the correct interpretation of the data. Nevertheless, when mean body weight is determined it is possible to make some cautious statements about the nutritional conditions of fish sampled in the nearby areas. From the mean fingerlings weight of 11.24 g-12.42 g and their ambient temperature which ranged from 29.00-30.50oC, it can be assumed that they are in the same age group. Mean RNA/DNA ratio of the field samples taken in the late monsoon months were below to the ratio which was observed in ‘starved’ fish reared in the laboratory, and in fact, in almost all the samples diseased fingerlings were present.
Unfortunately, these fingerlings which were easily identified by the symptoms described by Eckmann it is assumed that all sampled fingerlings were affected by intestinal disease in varying degrees, and this led to the unexpectedly low RNA/DNA ratio [40]. Barron examined the influence of toxicant stress on fathead minnows (Pimphales promelas Rafinesque) and found RNA/DNA and protein content per larvae, like RNA/DNA ratio, to be sensitive to toxicant stress [41]. A similar reaction might explain the low RNA/DNA ratios observed in diseased Anabas fingerlings in this study.
The Ca/P ratio in the muscle of the fish fed the diet ED01-ED10 increased significantly (P<0.01) with increasing dietary protein level and good quality protein source, when well fed fish compared to the fish fed ‘limited’ food and ‘starvation’ interval and sampled fish (same size group) of adjoining areas (Figures 1d and 2d). A definite correlation between serum calcium and phosphorus ratio was recorded in fish fed with different diets [42,43]. The result of the present investigation also indicated a positive correlation between the Ca/P ratio and dietary protein level and quality source. Again a positive correlation was observed between dietary Ca/P concentration and growth of the fish. Although, no definite relation was found between the levels of calcium and phosphorus in the diet and muscle could be established in the present study with the diet ED06-ED10.
Relatively little attention has been focused on protein biosynthesis in fish, although studied [44,45] suggest that, significant findings could be derived from research on mechanism of protein biosynthesis on aquatic species. The incorporation of 14C L-leucine (cpm mg protein -1) in the liver and muscle indicated a direct relationship with the protein levels and source in the diets. On the other hand, protein synthesis rates in the fish fed with the diets ED01-ED10 followed the same trend exhibited by the growth rate of the fish. And it is significantly (P<0.001) higher in well fed fish (ED01-ED10) as compared with the field samples and the ‘limited’ and ‘starved’ fish. It is also found that, the fingerlings were affected by an intestinal diseases which was caused either by malnutrition (limited food with starvation interval) or by the ingestion of unsuitable diets (copepods and others) in the field samples collected from nearby areas during late monsoon (Figures 2e,2f and 3e,3f) when judge under microscope.
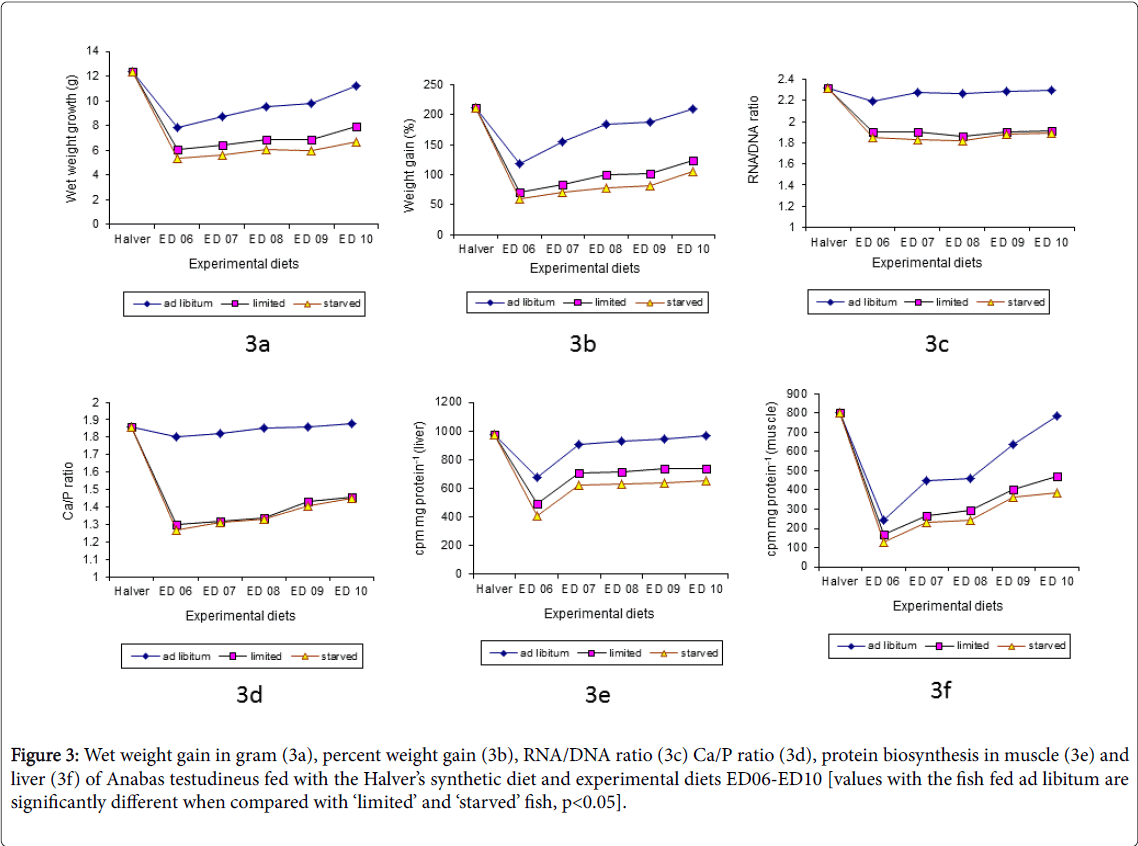
Figure 3: Wet weight gain in gram (3a), percent weight gain (3b), RNA/DNA ratio (3c) Ca/P ratio (3d), protein biosynthesis in muscle (3e) and liver (3f) of Anabas testudineus fed with the Halver’s synthetic diet and experimental diets ED06-ED10 [values with the fish fed ad libitum are significantly different when compared with ‘limited’ and ‘starved’ fish, p<0.05].
In the present study high rate of protein biosynthesis in the liver and muscle is consistent with the findings of when fish fed ad libitum with the diet ED01-ED05 and ED06-ED10 [45]. The diet prepared with animal protein components (ED07-ED10) showed higher rates of protein synthesis as compared to the plant protein component (ED06). Similar observation has been reported in the Clarias batrachus and in A. testudinus [46].
The growth of the fish A. testudineus is depend on the quantity and quality of the ingested food.
The fish fed with the nutritionally balanced artificial diet are rich in nutritional condition when judged with the help of growth, RNA/DNA and Ca/P ratio and the rate of protein biosynthesis in liver and muscle in comparison to the field samples. It will also help to characterize the nutritional condition of healthy Anabas fingerlings as well as to identify pathological situations taking in to account the fingerlings age and weight or length.
The authors would like to thank the Science for Equity Empowerment and Society Program, Department of Science and Technology, Govt. of India for financial assistance to carried out the research work (Project file No.: SEED/SCSP/008/ 2014 [G]). We also extend our gratitude to the fishermen community who has shared their valuable opinion.
Authors declare there have no conflict of interest.