Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2015) Volume 4, Issue 3
Curcumin is the main curcuminoid found in the yellow spice turmeric, a prominent member of the ginger family. Studies have revealed that curcumin exhibits numerous beneficial effects such as the ability to reduce inflammation and oxidative stress. The present study examines the neuroprotective effects of curcumin in vitro by subjecting B35 and SH-SY5Y neuroblastoma cells to hydrogen peroxide (H2 O2 ) followed or preceded by treating them with curcumin. Using curcumin concentrations of 5, 10 and 20 µM before and after damaging the cells with H2 O2 has resulted in an increase in cell viability of B35 neuroblastoma cells. In contrast, SH-SY5Y neuroblastoma cells showed an increase in their viability only upon the post-treatment with curcumin. The inhibitory effect of curcumin on caspase-3 and caspase-9, two of the most important mediators in the process of apoptosis, was also examined. We found that curcumin inhibited caspase-3 in a concentration-dependent manner, but not caspase-9. Using 5, 10, and 20 µM of curcumin resulted in 2.6%, 7.9% and 12.2% caspase-3 inhibition, respectively. These findings suggest that curcumin acts as a neuroprotectant and an anti-apoptotic agent through the inhibition of caspase-3, thereby introducing a potential agent for the treatment or prevention of neurodegenerative diseases.
Keywords: Curcumin; Neuroprotection; Apoptosis; Caspase-3; Caspase-9
Turmeric (Curcuma longa), is a yellow curry spice that contains curcuminoids (diferuloylmethanes). Curcumin, a yellow phenolic pigment, is the main member of the curcuminoids family which comprises of three prominent members found in their tautomeric forms: curcumin I (curcumin), curcumin II (demethoxycurcumin), and curcumin III (bisdemethoxycurcumin; Figure 1). Curcumin I is the most abundant and active form.
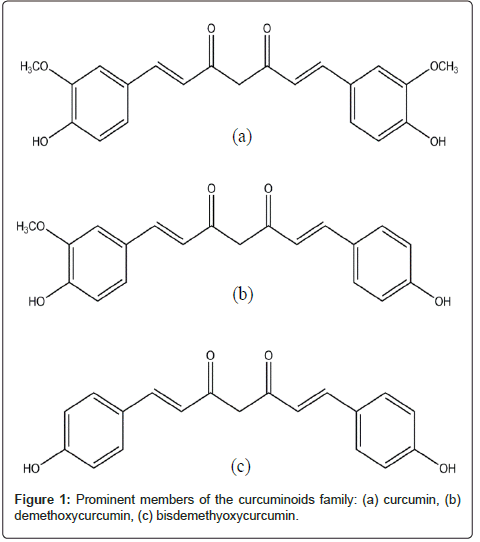
Figure 1: Prominent members of the curcuminoids family: (a) curcumin, (b) demethoxycurcumin, (c) bisdemethyoxycurcumin.
Research has shown that curcumin is an anti-oxidant, free radical scavenger [1-3], anti-inflammatory [4], anti-amyloid [5-7] and antiischemic [8]. Studies have also revealed that curcumin regulates numerous transcription factors, cytokines, protein kinases, adhesion molecules, redox status, and enzymes that have been linked to inflammation. Furthermore, curcumin has showed neuroprotective effects by protecting neurons against oxidative stress and interfering with the process of apoptosis [9-11].
While reduced apoptosis can lead to the development of cancerous tissue due to uncontrolled cell growth, increased apoptosis can lead to a number of neurological and autoimmune disorders as well [12]. This information promoted the development of several therapeutic agents that have been targeted against one or more of the steps involved in this programmed cell death to either activate or inhibit the process depending on the disease state. This study focuses on inhibiting apoptosis. When appropriate cell death stimuli are present, apoptosis is initiated through the activation of apoptotic factors and caspases. Caspases are a family of cysteine-aspartic proteases that play a crucial role in apoptosis induced by various deleterious and physiologic stimuli. Caspase-3 and caspase-9 are two of the main mediators in the apoptotic signaling pathway. The activation of caspase-3 by caspase-9 results in cellular death. Inhibition of caspases can delay or even inhibit apoptosis, indicating a potential application in neurodegenerative disorders.
In order to study the effects of curcumin on apoptosis, cellular stress can be induced and the change in cell viability can then be measured. Differential cell viability upon the addition of curcumin to the cells would be an indication of neuroprotective activity. One way of inducing stress in cells is through the use of the neurodamaging agent hydrogen peroxide (H2O2) [13-15]. Endogenously, it is formed from a superoxide anion through superoxide dismutase producing reactive oxygen species. H2O2 alters mitochondrial membrane permeability resulting in the release of cytochrome c into the cytoplasm, which in turn activates the caspase cascade leading to apoptosis.
The purpose of this study is to evaluate the neuroprotective effects of curcumin on SH-SY5Y human and B35 rat neuroblastoma cells. The inhibitory effect of curcumin on caspase-3 and caspase-9 is explored as the possible mechanism of neuroprotection.
Curcuminoids
Curcumin (>90% pure curcumin according to the manufacturer) was obtained from Cayman, 1180 East Ellsworth Road, Ann Arbor, Michigan 48108, USA. Thin layer chromatography (TLC) was used to assess the purity of curcumin. A mixture of chloroform: ethanol: glacial acetic acid (95:5:1) was used as mobile phase. Curcumin was dissolved in methanol at a final concentration of 0.5 g/mL and sonicated for 10 minutes. The stationary phase was a silica gel 60 HP-TLC plate (Whatman, P.O. Box 643065 Pittsburgh, Pennsylvania 15264, USA). UV light detector was used to analyze the TLC plate using wavelengths of 355 and 254 nm.
Cell culture
SH-SY5Y human and B35 rat neuroblastoma cells were grown at 37º C under a humidified atmosphere of 5% CO2. The cells were cultured in Dubelcco’s Modified Eagle’s Medium (DMEM) with 10% fetal bovine serum (FBS), 50 U/ml penicillin, and 50 μg/ml streptomycin until 80-90% confluent growth was achieved. The cells were harvested by using 0.25% trypsin-EDTA solution. Cell counting was performed with a cellometer by using the trypan blue exclusion technique. The aforementioned reagents were purchased from Mediatech, Inc. a Corning Subsidiary, 9345 Discovery Blvd., Manassas, Virginia 20109, USA. The cells were supplied by ATCC, 10801 University Blvd., Manassas, Virginia 20110, USA.
Cell treatments
Solutions of curcumin were prepared by using 10% dimethyl sulfoxide (DMSO) and polyethylene glycol (PEG) to give concentrations of 1-20 μM [7,13,16,17]. Hydrogen peroxide (H2O2) was dissolved in phosphate buffered saline (PBS) to final concentrations of 100- 1000 μM [16,18]. DMSO, PEG, H2O2 and PBS were purchased from Sigma Aldrich, P.O.Box 14508, St. Louis, Missouri 63178, USA. In the pretreatment assay, different concentrations of curcumin were added to wells and after two hours incubation at 37º C, the cells were treated with H2O2 and incubated for additional two hours. In the post-treatment assays, curcumin was added two hours after the initial addition of H2O2, and the cells were also incubated at 37º C for two more hours.
Cell viability assay
The viability of the cells was measured using a colorimetric method to determine the extent of neurodegeneration or neuroprotection. This method uses (3-(4,5-dimethylthiazol-2-yl)-5-(3- carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt; MTS) along with an electron coupling reagent, phenazine ethosulfate (PES) [19-21]. MTS can be bioreduced by the metabolic activity of cells into a formazan product that is soluble in the cell culture medium. 5.0 x 104 cells/well of SH-SY5Y and B35 cells were seeded in 96-well plates and treated as previously described (see 2.3 cell treatments). 10 μL of MTS was added to the cells followed by three hours of incubation. Absorbance measurement was performed on the well plates using a multi-mode microplate reader at a wavelength of 490 nm (Synergy HT, Biotek, 100 Tigan Street, Winooski, Vermont 05404, USA). Determination of cell viability is possible as the concentration of the formazan product is directly proportional to the number of metabolically active cells. MTS was obtained from Promega, 2800 Woods Hollow Road, Madison, Wisconsin 53711, USA.
Caspase inhibition assays
Caspase-3 and -9 (Abcam, 1 Kendall Square, Cambridge, Massachusetts 02139, USA) and their respective assay substrates: aspartic acid, glutamic acid, valine, aspartic acid, 7-amino-4- trifluoromethyl coumarin (DEVD-AFC) and leucine, glutamic acid, histidine, aspartic acid, 7-amino-4-trifluoromethyl coumarin (LEHDAFC) were used to study the effect of curcumin on both caspases. The enzymatic activity was measured by detecting the fluorescence emitted from free 7-amino-4-trifluoromethyl coumarin (AFC) upon its release in the media by the action of caspase-3 or -9. Using a multi-mode microplate reader, (Synergy HT, Biotek, 100 Tigan Street, Winooski, Vermont 05404, USA), fluorescence was detected at an excitation wavelength of 360/40 nm and an emission wavelength of 528/20 nm. Standard caspase inhibitors were used as positive controls: Z-DEVDfluoromethyl ketone (FMK) for caspase-3 and Z-LEHD-FMK for caspase-9 (Biovision, 155 South Milpitas Blvd., Milpitas, California 95035, USA) [22,23]. Fluorescence was measured after pure caspases were mixed with their substrates followed by the addition of curcumin (5−20 μM) or standard inhibitors (10−20 μM). The lower fluorescence value corresponds to the better caspase inhibition.
Statistics
Data is presented as mean ± SEM. Statistical significance is set at a level of p<0.05 using one way ANOVA, followed by Dunnett’s test for all groups compared to the control. At least three replicates were used per treatment in a 96-well plate, and each experiment was repeated three times (n=3).
Purity analysis of curcuminoids
Three bands were detected on the TLC plate under 355 nm UV light. The retention factor (Rf) values of 0.342, 0.516 and 0.645 were observed, which matched standard Rf values found in the literature (Plant Drug Analysis: A thin layer Chromatography Atlas by Wagner and Bladt) [24]. The compounds separated by TLC were bisdemethoxycurcumin (Rf 0.342), demethoxycurcumin (Rf 0.516) and curcumin (Rf 0.645) (Figure 2). When using 254 nm UV light, only two bands were detected; curcumin and demethoxycurcumin.
Figure 2: Inhibitory effect of the Morinda citrifolia 70% aqueous-ethanolic fruits (Mc.Cr.F) (A) and leaves extract (Mc.Cr.L) (B) on spontaneous and high K+- induced contractions and concentration-response curves of Ca++ in isolated rabbit jejunum. The symbols represent mean ± S.E.M, n= 4-6.
Although the curcumin sample used in the study was a mixture of curcuminoids (90% curcumin as reported by the producer, Cayman), all three compounds share similar chemical structures and pharmacological activities. Curcumin is the least polar among the three compounds as indicated by its high Rf value (due to least interaction with silica gel). On the other hand, the most polar compound, bisdemethoxycurcumin, had the highest interaction with the stationary phase which resulted in the lowest Rf value. Curcumin’s band on the TLC plate was the biggest and most prominent of the three, which qualitatively indicated its abundance in the mixture (consistent with the producer).
Caspase inhibition assays
A concentration-dependent effect of curcumin on the inhibition of caspase-3 was observed with 5, 10, and 20 μM of curcumin resulting in 2.6%, 7.9% and 12.2% caspase-3 inhibition, respectively. The standard inhibitor Z-DEVD-FMK (positive control) inhibited caspase-3 by 38% (Figure 3). %inhibition values were obtained by comparison with the pure enzyme’s activity on its substrate as baseline (not shown on graph).
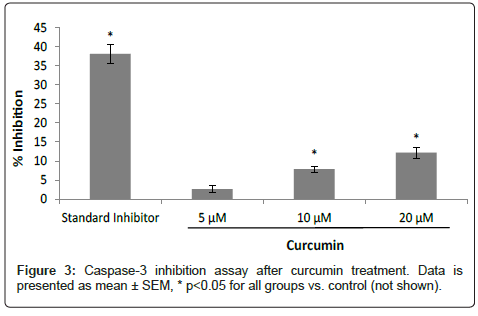
Figure 3: Caspase-3 inhibition assay after curcumin treatment. Data is presented as mean ± SEM, * p<0.05 for all groups vs. control (not shown).
Curcumin did not exhibit inhibitory effect on caspase-9, as none of the different concentrations of curcumin resulted in any statistically significant inhibition compared to the control. More research is needed to understand the full effect of curcumin on apoptosis and this enzyme.
Cell viability assays
The viability of B35 and SH-SY5Y neuroblastoma cells was assessed using the MTS assay. Different concentrations of H2O2 were used to stress the cells and evaluate their viability upon treating them with curcumin. Both cell types started undergoing apoptosis in about two hours after H2O2 exposure as indicated by their shrinkage and granulation (Figure 4).
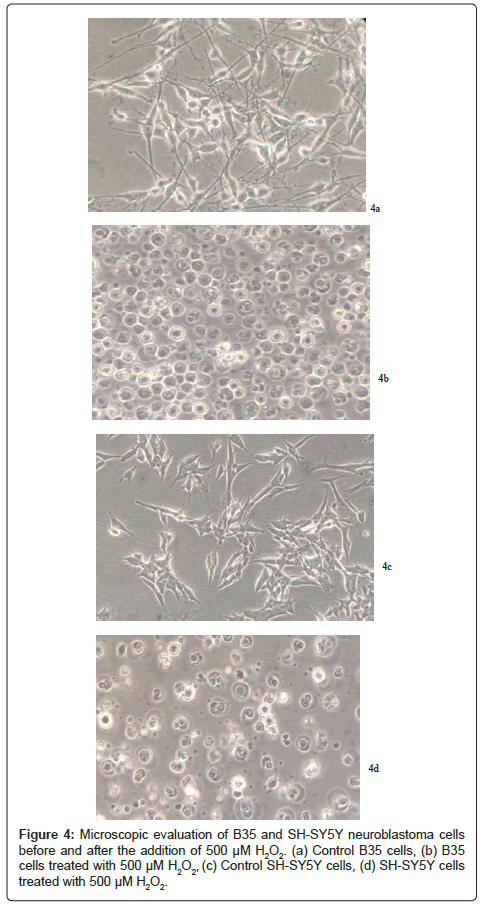
Figure 4: Microscopic evaluation of B35 and SH-SY5Y neuroblastoma cells before and after the addition of 500 μM H2O2. (a) Control B35 cells, (b) B35 cells treated with 500 μM H2O2, (c) Control SH-SY5Y cells, (d) SH-SY5Y cells treated with 500 μM H2O2
SH-SY5Y human neuroblastoma cells: Treatment with curcumin alone did not affect the viability of SH-SY5Y cells significantly (Figure 5). When curcumin was added to the cells two hours after the addition of H2O2, the percentage of cell viability increased compared to the cells treated with H2O2 alone. Post-treatment with 20 μM of curcumin in cells treated with 500 μM and 1000 μM of H2O2 increased cell viability from 57.4% and 52.3% to 90.3% and 102.3%, respectively. The cell viability increases observed with the 5 and 10 μM curcumin posttreatment were not statistically significant (Figure 6). Pretreatment with curcumin two hours before adding H2O2 to the cells did not prevent the decrease in their viability (Figure 7).
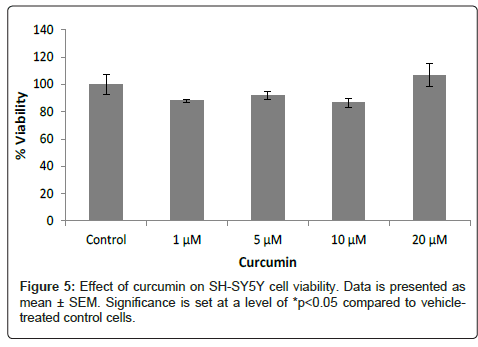
Figure 5: Effect of curcumin on SH-SY5Y cell viability. Data is presented as mean ± SEM. Significance is set at a level of *p<0.05 compared to vehicletreated control cells.
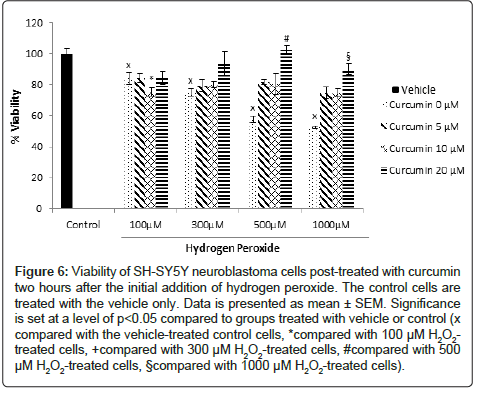
Figure 6: Viability of SH-SY5Y neuroblastoma cells post-treated with curcumin two hours after the initial addition of hydrogen peroxide. The control cells are treated with the vehicle only. Data is presented as mean ± SEM. Significance is set at a level of p<0.05 compared to groups treated with vehicle or control (x compared with the vehicle-treated control cells, *compared with 100 μM H2O2- treated cells, +compared with 300 μM H2O2-treated cells, #compared with 500 μM H2O2-treated cells, §compared with 1000 μM H2O2-treated cells).
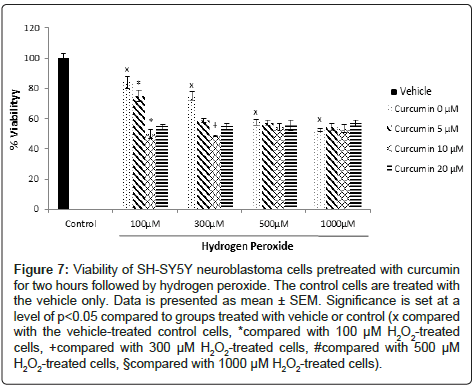
Figure 7: Viability of SH-SY5Y neuroblastoma cells pretreated with curcumin for two hours followed by hydrogen peroxide. The control cells are treated with the vehicle only. Data is presented as mean ± SEM. Significance is set at a level of p<0.05 compared to groups treated with vehicle or control (x compared with the vehicle-treated control cells, *compared with 100 μM H2O2-treated cells, +compared with 300 μM H2O2-treated cells, #compared with 500 μM H2O2-treated cells, §compared with 1000 μM H2O2-treated cells).
B35 rat neuroblastoma cells: Curcumin had no direct effect on the viability of B35 cells when used alone (Figure 8). When used as postand pretreatment, curcumin increased the viability of the cells damaged by H2O2. In the cells treated with H2O2 followed by the post-treatment with curcumin, the viability of the 300 μM H2O2-treated cells increased from 59.8% to around 71% after the addition of 5, 10 and 20 μM of curcumin. The cells treated with 500 μM H2O2 had a similar increase in their viability from 50.6% to 78.2%, 72.3% and 74.2% after adding 5, 10 and 20 μM of curcumin, respectively. As for the cells treated with the highest H2O2 concentration, 1000 μM, the viability increased from 22.4% to 67.5%, 69.5% and 70.2% following the addition of 5, 10 and 20 μM of curcumin, respectively (Figure 9).
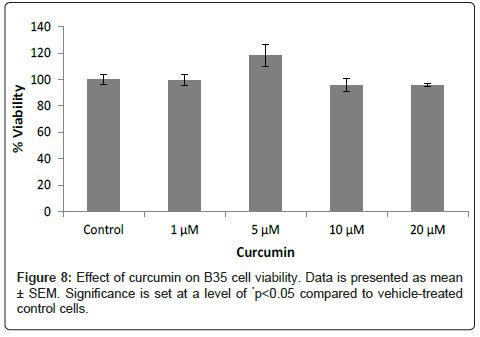
Figure 8: Effect of curcumin on B35 cell viability. Data is presented as mean ± SEM. Significance is set at a level of *p<0.05 compared to vehicle-treated control cells.
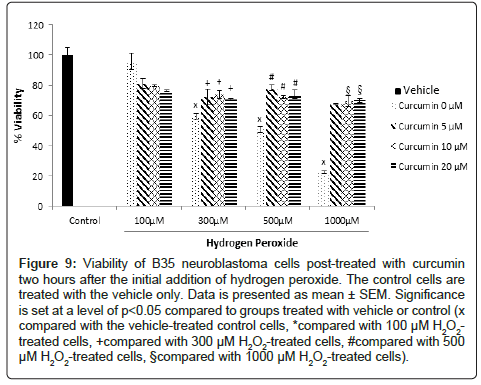
Figure 9: Viability of B35 neuroblastoma cells post-treated with curcumin two hours after the initial addition of hydrogen peroxide. The control cells are treated with the vehicle only. Data is presented as mean ± SEM. Significance is set at a level of p<0.05 compared to groups treated with vehicle or control (x compared with the vehicle-treated control cells, *compared with 100 μM H2O2- treated cells, +compared with 300 μM H2O2-treated cells, #compared with 500 μM H2O2-treated cells, §compared with 1000 μM H2O2-treated cells).
In the cells pretreated with curcumin and followed by H2O2 treatment, the viability of the 1000 μM H2O2-treated cells increased from 22.4% to 63.8%, 56.7% and 63.2% upon adding 5, 10 and 20 μM curcumin, respectively. As for the cells treated with 500 μM H2O2, their viability increased from 50.6% to 75%, 60.5% and 61.1% following the addition of 5, 10 and 20 μM curcumin, respectively. The viability of the 300 μM H2O2-treated cells increased from 59.8% to 102.6%, 63.9% and 60.6% upon using the same respective concentrations of curcumin (Figure 10).
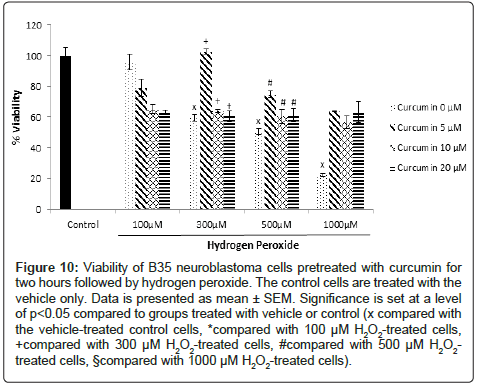
Figure 10: Viability of B35 neuroblastoma cells pretreated with curcumin for two hours followed by hydrogen peroxide. The control cells are treated with the vehicle only. Data is presented as mean ± SEM. Significance is set at a level of p<0.05 compared to groups treated with vehicle or control (x compared with the vehicle-treated control cells, *compared with 100 μM H2O2-treated cells, +compared with 300 μM H2O2-treated cells, #compared with 500 μM H2O2- treated cells, §compared with 1000 μM H2O2-treated cells).
The results obtained from our study support preliminary data showing the neuroprotective effects of curcumin on cells [25-27]. Although the neurodamaging effect of 300-1000 μM of H2O2 was more prominent in B35 cells compared to SH-SY5Y cells (as manifested by the greater reduction in cell viability), both cell lines showed the neuroprotective effect from the treatment with curcumin. Cell viability was enhanced in B35 cells treated with H2O2 upon the postand pretreatment with curcumin suggesting that curcumin exhibits protective effects in cells exposed to oxidative stress. However, the viability of SH-SY5Y cells increased only when curcumin was used as post-treatment. With the higher concentrations of H2O2 (500 and 1,000 μM), 20 μM of curcumin showed the best neuroprotection; whereas with the lower concentrations of H2O2 (100 and 300 μM), 5 μM of curcumin was sufficient to improve cell viability. Protecting SH-SY5Y cells from oxidative stress and beta amyloid induced neurotoxicity by treating them with curcumin have also been shown in other studies [6,7,17,27-29]. Further research is needed to explain the mechanism behind the differences in cell viability when curcumin is used as pretreatment versus post-treatment in SH-SY5Y cells.
The therapeutic implications of curcumin in the treatment of neurodegenerative diseases such as Alzheimer’s and Parkinson’s have been demonstrated in animal models as well [30-32], where disease biomarkers have subsided and memory and learning improved in mice. Moreover, curcumin inhibited neuroinflammation and halted intracerebral hemorrhage in mice brain [33]. It was also shown to attenuate fluoride and aluminum-induced neurotoxicity [34,35], ameliorate the toxicity of TAR DNA-binding protein-43 (TDP-43) which plays a role in the pathophysiology of amyotrophic lateral sclerosis [36] and protect from oxidative stress caused by 1-methyl- 4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) which is a potent neurotoxin that is responsible for Parkinsonian symptoms [37]. Furthermore, curcumin’s antioxidant effect was found to reduce intraocular pressure in a glaucoma animal model [38] and mitigate the damage produced by cerebral ischemia in rats [39]. Our results add to the body of evidence available for the neuroprotective effects of curcumin by showing the differences in neuroblastoma cell viability after the exposure to the natural compound. The results herein match the data from in vivo studies and hold promising implications for curcumin.
When curcumin was added to active caspase-3 and its substrate, the fluorescence emitted from the substrate upon its cleavage by caspase-3 decreased, indicating the reduction or inhibition of caspase activity. This decrease was inversely related to the concentrations of curcumin. 20 μM curcumin exhibited the best inhibition of caspase-3 (12.2%) compared to the inhibition observed with 5 and 10 μM (2.6% and 7.9%, respectively). The standard irreversible inhibitor Z-DEVDFMK showed a greater inhibition than curcumin. Since caspases are important mediators of cell death, inhibition of caspase-3 can lead to uncontrolled cell growth which may result in disorders such as cancer. Therefore, the best extent of caspase-3 inhibition needed to produce an anti-apoptotic effect without creating major side effects would be low to moderate. Caspase-3 is the most notable apoptotic mediator involved in neurodegeneration. It is an effector caspase that works downstream in the apoptotic signaling pathway leading to protein cleavage and cell death, as compared to caspase-9 which is an initiator caspase that acts in the early phases of the proteolytic cascade [40]. Our results show that curcumin had a more prominent effect on caspase-3 than caspase-9. This might be due to better binding of the natural compound to caspase-3 thereby impeding the interaction between the enzyme and its substrate. Subsequently, less fluorescence was emitted by the reduced proteolytic activity of caspase-3. Future research can focus on finding curcumin’s antagonistic effect on other markers of apoptosis in order to understand the mechanism of neuroprotection.
It is worthy to mention that curcumin displays opposite effects on the process of apoptosis depending on the concentration being used. Numerous studies have shown that curcumin exhibits anticancer properties by promoting apoptosis when used at high concentrations (30−100 μM) [41-47]. In contrast, low concentrations of curcumin (1−20 μM) inhibit the activation of apoptosis leading to neuroprotection and cell preservation [6,14,48]. Therefore, the choice of the right concentration will be crucial for future studies [49].
These findings may introduce new hypotheses in the study of the natural product curcumin as a potential agent to produce neuroprotection through the modulation of apoptosis. Future research will focus on measuring activation of caspases and other apoptotic markers upon cellular stress in order to locate new targets for therapeutic intervention.
We thank the Summer Undergraduate Research Fellowship (SURF) at MCPHS University for supporting this research.
The authors have no conflicts of interests to declare.