Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2015) Volume 2, Issue 2
In this study, the protective role of salicylic acid (SA) and trehalose (Tre) in relation to lipid peroxidation, membrane leakage, membrane stability index, antioxidant defense system as well as non-enzymatic antioxidants were investigated in drought stressed wheat Gemmieza-7 (drought sensitive cultivar) and Sahel-1 (drought tolerant cultivar) plants. Water stress reduced bio-membranes stability through inducing its lipid peroxidation resulting in an increment in membrane leakage with marked decrease in membrane stability index of flag leaf of both wheat cultivars during grain-filling. Moreover, it was obvious that drought significantly increased the activity of ascorbic acid oxidase (AAO), peroxidase (POD) and phenylalanine ammonia lyase (PAL) activities and induced non-significant reduction in polyphenol oxidase (PPO) activity in flag leaves of both wheat cultivars during grain-filling in comparing with well watered plants. Among cultivars, tolerant one showed higher enzymes activity than the sensitive one. Application of SA and/or Tre markedly increased AAO, POD and PAL activities and non-significant decrease in PPO activity in flag leaf of water stressed wheat plants. Generally, SA and Tre treatment appeared to be the most effective treatment in counteracting the negative effects of water stress and Sahel-1 appeared to induce better results than Gemmieza-7 and proved to be more tolerant.
Drought stress caused significant increase in the amount of total phenols and flavonoids in flag leaf of both cultivars during grain-filling where the sensitive plants accumulated more total phenols and flavonoids contents than the tolerant one. Furthermore, water stress increased the non-photosynthetic pigment content of the two wheat cultivars particularly drought sensitive one during grain filling. These results suggest that the exogenous application of SA and/or Tre assisted the plants to become more tolerant to drought stress-induced oxidative damage by upregulating the membrane characteristics and enhancing their antioxidant defense system as well as non- enzymatic antioxidants.
Keywords: Wheat; Drought; Salicylic acid; Trehalose; Membrane characteristics; Antioxidive system
ROS: Reactive Oxygen Species; MDA: Malondialdehyde; AAO: Ascorbic Acid Oxidase; PAL: Phenylalanine Ammonia Lyase; POX: Peroxidase; PPO: Polyphenol Oxidase
Drought is one of the major abiotic stresses which adversely affect crop growth and yield and thus a constraint for plant productivity worldwide [1,2]. Drought stress adversely affects a variety of vital physiological and biochemical processes in plants [2].
The integrity of cellular membrane structure and the preservation of membranes are of primary importance in the tolerance of some plant species to water stress [3]. Maintenance of membrane integrity and function under a given level of dehydration has been used as a measure of drought tolerance [4]. The degree of cell membrane injury induced by water stress may be easily estimated through measurements of membrane stability index (MSI) [5]. In addition, membrane leakage (ML) to cell electrolytes is an indicator of cell membrane integrity, and so it is commonly considered as one of the best physiological components of drought tolerance in plants [6]. Furthermore, malondialdehyde (MDA) content is usually used to measure the extent of lipid peroxidation resulting from oxidative stress under water deficit conditions [1,7]. The three parameters (MSI, ML and lipid peroxidation expressed as MDA content) are greatly interrelated. In this connection, increased MDA content was recorded to be accompanied by an increase of ML in drought-stressed plants, which indicated that lipid peroxidation led to membrane fluidity resulting in enhanced membrane permeability, as proved by lowered MSI values [7].
The toxic effect of water stress is through oxidative stress caused by enhanced production of reactive oxygen species (ROS) [8]. The ROS are extremely reactive in nature because they can interact with a number of cellular molecules and metabolites, thereby leading to irreparable metabolic dysfunction and death [9]. Antioxidant defense system has been touted as beneficial for enhancing plant stand and mitigating the effects of biotic and abiotic stresses [10]. Moreover, antioxidant defense system comprises both enzymatic and nonenzymatic components [11]. Enzymatic antioxidants include superoxide dismutase (SOD), catalase (CAT), ascorbic acid peroxidase (APX) and peroxidase (POD) [12] as well as ascorbic acid oxidase (AAO). Ascorbic acid oxidase (AAO) catalyzes a complex reaction in which four electrons are transferred simultaneously from ascorbic acid to molecular oxygen, yielding two water molecules [13].
The ability of higher plants to overcome the toxic effects of active oxygen seems to be very important for their tolerance to stresses. Antioxidants are the first line of defense against free radical damage [14]. There are several compounds which contribute to the antioxidative properties, these include anthocyanin, polyphenols, vitamin C, flavonoids and carotene [15]. Under salt stress, the level of nonenzymatic antioxidant was increased, due to their capacity to protect itself against oxidative stress [14].
Several studies support a major role of SA in modulating the plant response to various abiotic stresses [16,17]. In addition, it has been found that plants treated with SA generally exhibited better resistance to drought stress [17]. Treatment of drought stressed plants with exogenous SA was found to be effective in modulating both enzymatic and non-enzymatic components of antioxidant defense system [17,18]. Application of SA significantly decreased lipid peroxidation in sweet basil under salinity stress condition [19]. In addition, Yazdanpanah et al. [20] reported that SA application declined adverse effect of drought in savory by decreasing malondialdehyde (MDA) and other aldehydes. Moreover, there is evidence that exogenous application of SA can alter antioxidant capacity in plants, thereby providing protection against oxidative damage [21] and inducing stress tolerance. In general, low concentrations of SA may enhance the antioxidant capacity in plants, but high concentrations of SA may cause cell death or susceptibility to abiotic stresses [22].
Trehalose (Tre), is a non-reducing disaccharide, which involves in the acquisition of various stress tolerance [23]. Organic compatible solutes like Tre play important roles under multiple abiotic stresses [24]. Among different compatible solutes, Tre is a non-reducing disaccharide of glucose that stabilizes biological structures and macromolecules such as proteins and membrane lipids during dehydration and other abiotic stresses [25]. Tre as an osmoprotectant maintains cellular osmotic balance. It stabilizes dehydrated enzymes, proteins and lipid membranes efficiently [26]. It protects biological structures from damage at desiccation [27]. Furthermore, Tre has been shown to efficiently stabilize dehydrated enzymes, proteins and lipid membranes, as well as protect biological structures from damage during desiccation by replacing water. Moreover, Tre forms an amorphous glass structure that limits molecular motion, preventing protein aggregation and scavenging free radical under stress conditions [28]. Tre treatment protected Arabidopsis thaliana against drought stress by increased survival rates, increased cell membrane stability and a decreased leakage of electrolytes after the stress treatment [29]. Moreover, Tre acts as a direct and indirect scavenger of ROS [29]. Exogenously applied Tre decreased electrolyte leakage and MDA content with increasing concentration of Tre [23].
The objective of this study was to evaluate the protective role of salicylic acid (SA) and trehalose (Tre) in relation to lipid peroxidation, membrane leakage, membrane stability index and antioxidant defense system as well as non-enzymatic antioxidants in relatively drought tolerant (Sahel-1) and susceptible (Gemmieza-7) wheat genotypes.
Plant material and growth conditions
Pure strains of Triticum aestivum L. Gemmieza-7 (drought sensitive cultivar) and Sahel-1 (drought tolerant cultivar) were kindly supplied by the Agricultural Research Center, Ministry of Agriculture, Giza, Egypt.
For soaking experiment, a homogenous lot of Triticum aestivum L. (i.e. either sensitive or tolerant cultivar) grains were selected. The grains were separately surface sterilized by soaking in 0.01 M HgCl2 solution for three minutes, then washed thoroughly with distilled water. The sterilized grains from each cultivar were divided into two sets (≈ 500 g per set for each cultivar). Grains of the 1st set were soaked in distilled water to serve as control, while those of the 2nd were soaked in salicylic acid (3 mM) for about 6 hours.
After soaking, thoroughly washed grains were drilled in 20 November 2011 and 2012 in plastic pots (20 cm in diameter) filled with 5.5 kg soil (clay/sand 2/1, v/v), where fifteen grains was sown in each pot. The pots were then kept in a greenhouse at Botany Department, Faculty of Science, Mansoura University, Egypt. The plants were subjected to natural day/night conditions (minimum/ maximum air temperature and relative humidity were 15/25°C and 35/45%; respectively) at mid-day during the experimental period. The plants in all sets were irrigated to field capacity by tap water.
After two weeks from sowing, thinning was started so that five uniform seedlings were left in each pot for the subsequent studies. On the day 65 after planting (at the beginning of heading) the pots of the 1st set was allocated to four groups (20 pots per each group) as follows: control (cont.), water stress (WS), trehalose control, trehalose + water stress (trehalose + WS). The 2nd set group was allocated to four groups as follows: salicylic acid control (SA), salicylic acid + water stress (SA +WS), control trehalose + salicylic acid (SA + trehalose) and salicylic acid + trehalose + water stress (SA+ trehalose +WS). For trehalose (1.5 mM) treatment, the plants were sprayed by trehalose 48 hrs before starting the stress period and weekly during the stress period.
Water deficit was imposed by withholding water at the reproductive stage for 30 days within two periods: on the day 65 from planting (heading stage) and at the day 80 from planting (anthesis stage). Each droughted pot received 500 ml water at the end of 1st stress period. At the end of stress periods, rewatering to the field capacity was carried out. The undroughted (control) plants were irrigated to the field capacity during the stress period, and all plants were left to grow until grain maturation under normal irrigation with tap water. After thinning and at heading, the plants received 36 kg N h-1 as urea and 25 kg P ha-1 as super-phosphate.
Three samples from flag leaves were taken from each treatment for the biochemical analyses during grain-filling (21 days post-anthesis) (i.e. 106 days after sowing). Data were obtained and the mean values (per plant) were computed for each treatment.
Determination of membrane stability index
The membrane stability index (MSI) was determined according to Sairam et al. [30].
Determination of membrane leakage
Membrane leakage (ML) was determined according to Vahala et al. [31].
Estimation of lipid peroxidation
Estimation of lipid peroxidation was assayed spectrophotometrically using thiobarbituric acid-malondialdehyde assay (TBA-MDA) according to the procedure adopted by Hodges et al. [32].
Antioxidant enzymes
Preparation of enzymatic extract: For the extraction of antioxidant enzymes, the method of Agarwal and Shaheen [33] was adopted.
Estimation of peroxidase activity (POD): Peroxidase activity was assayed as the increase in absorbance at 420 nm due to the formation of purpurogallin [34].
Estimation of polyphenol oxidase activity (PPO): Polyphenol oxidase activity was assayed as the absorbance at 420 nm due to the formation of purpurogallin [34].
Estimation of phenylalanine ammonia lyase activity (PAL): Phenylalanine ammonia lyase activity was determined according to Havir and Hanson [35].
Estimation of ascorbic acid oxidase (AAO) activity: AAO Activity was determined according to the fact that, the rate of oxygen consumption during ascorbic acid oxidation is proportional to the amount of enzyme present. AAO Activity was determined using the method of Oberbacher and Vines [36].
Phenolic constituents
Estimation of flavonoids: The total flavonoid content was determined following the spectrophotometric method of Dewanto et al. [37].
Estimation of total phenols: Total phenols were estimated using the method of Malik and Singh [38].
Non-photosynthetic pigment
Estimation of lycopene and β-carotene: Lycopene and β-carotene were determined according to the method of Nagata and Yamashita [39].
Estimation of anthocyanins: Anthocyanins were extracted from the oven-dried ground tissues by suspending in 10 ml of acidified methanol (methanol: water: HCl, 79: 20: 1, v/ v) and autoextracting at 0°C for 72 hours in dark with continuous shaking. The extracts were then centrifuged for 10 minutes at 5000xg and the absorbance was measured at 530 and 657 nm for each supernatant [40].
Anthocyanins = A530 - 1/3 A657
Statistical analysis
It should be mentioned that the sample numbers which were taken for investigation were as follows: three for all chemical analyses and only the mean values were represented in the respective figures. The data were subjected to one-way analysis of variance (ANOVA), and different letters indicate significant differences between treatments at p ≤ 0.05, according to CoHort/ CoStat software, Version 6.311.
Changes in membrane characteristics
Changes in lipid peroxidation: As compared to the control values, water stress induced a marked increase (p ≤ 0.05) in the values of lipid peroxidation of tolerant wheat cultivar and a non-significant increase in sensitive cultivar (Figure 1).
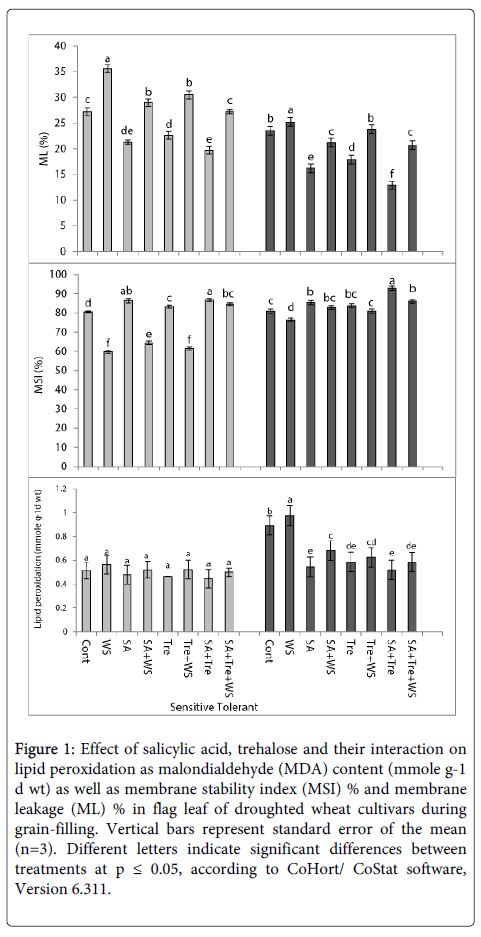
Figure 1: Effect of salicylic acid, trehalose and their interaction on lipid peroxidation as malondialdehyde (MDA) content (mmole g-1 d wt) as well as membrane stability index (MSI) % and membrane leakage (ML) % in flag leaf of droughted wheat cultivars during grain-filling. Vertical bars represent standard error of the mean (n=3). Different letters indicate significant differences between treatments at p ≤ 0.05, according to CoHort/ CoStat software, Version 6.311.
Application of SA and/or Tre caused significant decrease (p ≤ 0.05) in lipid peroxidation of tolerant wheat cultivar and a non- significant decrease in sensitive cultivar as compared with water-stressed plants. The interaction of SA and Tre was the most effective in decreasing the values of lipid peroxidation in flag leaf of stressed wheat plants.
Changes in membrane stability index (MSI) and membrane leakage (ML): Data cleared that the pattern of change in MSI% is opposite to that in ML% (Figure 1). As compared to control plants, the significant reduction (p ≤ 0.05) recorded in MSI% in response to water stress was accompanied with an increase in ML% of both wheat cultivars during grain-filling. The effect was more pronounced with the sensitive one. SA and/or Tre induced marked increase (p ≤ 0.05) in MSI of waterstressed wheat plants. In addition, application of SA and/or Tre reduced (p ≤ 0.05) the values of ML in stressed wheat plants. The interaction of SA and Tre was the most effective treatment. Generally, Sahel-1 induced better results than Gemmieza-7 and proved to be more tolerant.
Changes in antioxidant enzymes system
The data presented in Figures 2 and 3 showed that, water stress induced marked increase (p ≤ 0.05) in ascorbic acid oxidase (AAO), peroxidase (POD) and phenyl ammonia lyase (PAL) activities and induced a non-significant reduction in polyphenol oxidase (PPO) activity in flag leaves of both wheat cultivars during grain-filling comparing with the control plants. Among cultivars, tolerant one showed higher enzymes activity than sensitive one. Application of SA and/or Tre markedly increased (p ≤ 0.05) AAO, POD and PAL activities and a non- significant decrease in PPO activity in flag leaf of water stressed wheat plants. Generally, SA and Tre treatment appeared to be the most effective treatment in counteracting the negative effects of water stress on AAO, POD, PAL and PPO activities.
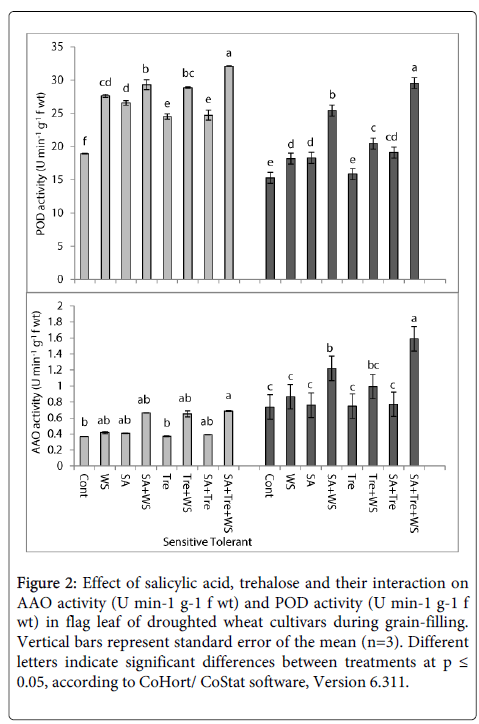
Figure 2: Effect of salicylic acid, trehalose and their interaction on AAO activity (U min-1 g-1 f wt) and POD activity (U min-1 g-1 f wt) in flag leaf of droughted wheat cultivars during grain-filling. Vertical bars represent standard error of the mean (n=3). Different letters indicate significant differences between treatments at p ≤ 0.05, according to CoHort/ CoStat software, Version 6.311.
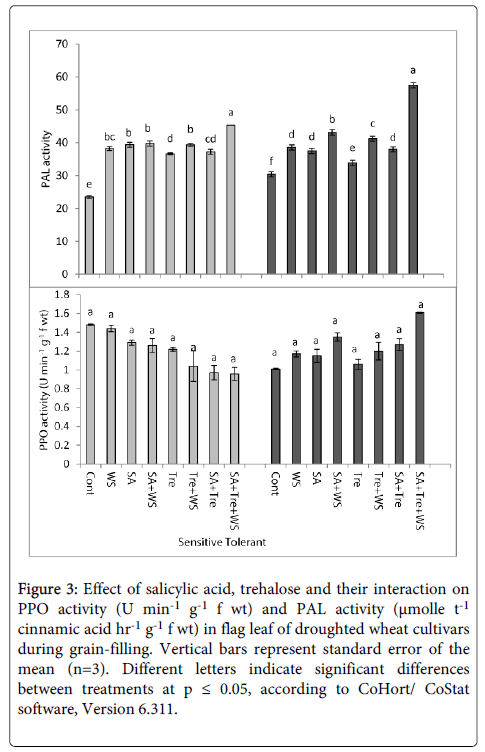
Figure 3: Effect of salicylic acid, trehalose and their interaction on PPO activity (U min-1 g-1 f wt) and PAL activity (μmolle t-1 cinnamic acid hr-1 g-1 f wt) in flag leaf of droughted wheat cultivars during grain-filling. Vertical bars represent standard error of the mean (n=3). Different letters indicate significant differences between treatments at p ≤ 0.05, according to CoHort/ CoStat software, Version 6.311.
Changes in non-enzymatic antioxidant
Data in Figure 4 revealed that drought stress caused significant increase (p ≤ 0.05) in the amount of total phenols and flavonoids in flag leaf of both cultivars during grain-filling where the sensitive plants accumulated more total phenols and flavonoids contents than the tolerant one. Application of SA and/or Tre markedly increased (p ≤ 0.05) total phenols and flavonoids contents. Generally, SA and Tre treatment appeared to be the most effective treatment in increasing total phenols and flavonoids contents in stressed wheat plants.
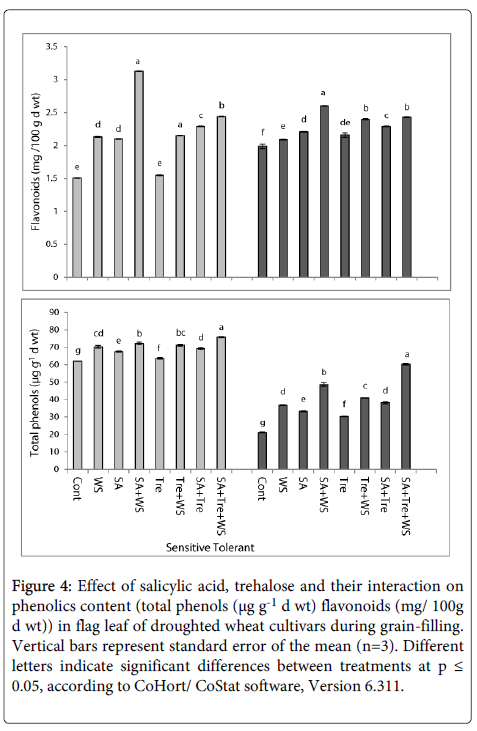
Figure 4: Effect of salicylic acid, trehalose and their interaction on phenolics content (total phenols (μg g-1 d wt) flavonoids (mg/ 100g d wt)) in flag leaf of droughted wheat cultivars during grain-filling. Vertical bars represent standard error of the mean (n=3). Different letters indicate significant differences between treatments at p ≤ 0.05, according to CoHort/ CoStat software, Version 6.311.
Changes in non-photosynthetic pigment
In relation to wheat cultivar, the flag leaves of the control tolerant plants had higher non-photosynthetic pigment (anthocyanin and β- carotene) contents than the sensitive one (Figure 5). Water stress resulted in a massive increase (p ≤ 0.05) in the non-photosynthetic pigment content of the two wheat cultivars particularly drought sensitive one during grain filling. Application SA and/or Tre induced additional increase (p ≤ 0.05) in anthocyanin, lycopene and β-carotene contents. The effect was more pronounced with SA treatment in case of anthocyanins pigment and with SA and Tre treatment in case of lycopene and β-carotene pigment.
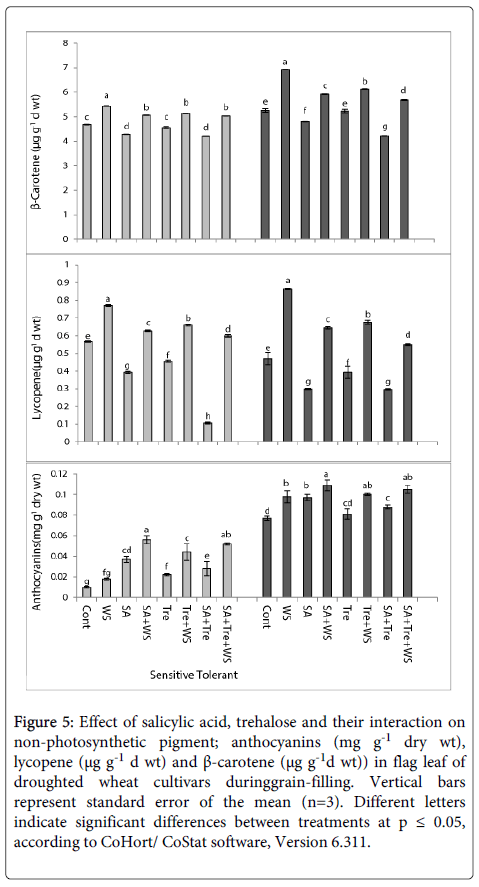
Figure 5: Effect of salicylic acid, trehalose and their interaction on non-photosynthetic pigment; anthocyanins (mg g-1 dry wt), lycopene (μg g-1 d wt) and β-carotene (μg g-1d wt)) in flag leaf of droughted wheat cultivars duringgrain-filling. Vertical bars represent standard error of the mean (n=3). Different letters indicate significant differences between treatments at p ≤ 0.05, according to CoHort/ CoStat software, Version 6.311.
Cell membrane is one of the main cellular targets common to different stress conditions [41]. The degree of membrane damage reflects the measure of plant tolerance to stresses such as drought [1]. In this regard, lipid peroxidation, membrane stability index (MSI) and membrane leakage (ML) could be considered as widely used stress indicators of plant membranes. In the present study, stress conditions caused significant increase in lipid peroxidation and ML with marked decrease in MSI of wheat plants. These results are in agreement with those obtained by Fazeli et al. [42] who demonstrated that drought could increase malondialdehyde (MDA) content in roots of two sesame cultivars causing significant increment in lipid peroxidation. Also, Jamil et al. [43] cleared that drought significantly decreased MSI in different wheat cultivars. Moreover, the amount of electrolyte leakage from the leaves of poplar plants was reported to increase under water stress conditions [44].
It is well known that water stress enhances free radical production, which induces the lipid peroxidation of bio membranes, reflecting the stress-induced damage in tissues [45]. Evidence suggests that membranes are the primary sites of injury to cells and organelles [46] because ROS can react with unsaturated fatty acids to cause peroxidation of essential membrane lipids in plasma lemma or intracellular organelles leading to leakage of cellular contents, rapid desiccation and cell death [47]. In the present study, lipid peroxidation was estimated as malondialdehyde (MAD) content which is one of the decomposition products of polyunsaturated fatty acids (PUFA) of biomembranes [48] where, PUFA are the main membrane lipid components susceptible to peroxidation and degradation [49].
Thus, the increase in MDA content might result from stomatal closure causing a decrease in leaf CO2 concentration. This, in turn, might cause a decrease in the concentration of NADP+ available to accept electrons from PSI and/or PSII and thus initiate O2 reduction with the concomitant generation of ROS [50].
Under water stress conditions, electrolyte leakage could be attributed to the damage of cell membranes which become more permeable due to less water availability [51]. In a study on maize plants under water deficit condition, Valentovic et al. [52] found that electrolyte leakage increased by drought treatment in two maize cultivars. In a similar trend, the electrolyte leakage increased as water deficit stress was intensified in dragonhead plants [53]. Quan et al. [54] also found higher electrolyte leakage in drought-stressed maize plants than in plants grown under normal conditions. The data obtained by Masoumi et al. [55] also indicated water stress-induced membrane injury, indicated by higher membrane leakage in Borujerd Kochia plants.
A major impact of environmental stress on plants is cellular membrane modification due to water stress, expressed in increased permeability and leakage of ions [56]. The degree of cellular membrane damage reflects the measure of plant tolerance to stresses [57]. In this respect, cell membrane stability or cell membrane injury is a physiological index widely used for the evaluation of plant tolerance to various forms of abiotic stress [58]. It was reported that stress could reduce bio-membranes stability resulting in an increment in membrane leakage [59]. Lower membrane stability, as indicated by higher leakage, under water deficit conditions could be ascribed to the fact that stress could denaturate proteins of bio-membranes and reduce plasma membrane H+-ATPase activity. In this context, stress caused great reduction in plasma membrane H+-ATPase activity and total plasma membrane protein concentration in buffalo grass [60].
Application of SA and/or Tre caused significant decrease in lipid peroxidation and ML with marked increase in MSI of wheat tolerant wheat cultivar and a non- significant decrease in sensitive cultivar as compared with water-stressed plants. Generally, Sahel-1 reflect better results than Gemmieza-7 and proved to be more tolerant. The interaction of SA and Tre was the most effective treatment. Hence, Sayyari et al. [61] showed that, SA application significantly decreased MDA accumulation and electrolyte leakage.
Moreover, application of SA significantly decreased lipid peroxidation in sweet basil under salinity stress condition [19]. In addition, Yazdanpanah et al. [20] reported that SA application declined adverse effect of drought in savory by decreasing Malondialdehyde (MDA) and other aldehydes.
Exogenously applied trehalose could decrease MDA and electrolyte leakage. The same results were reported by Li et al. [23] who noticed electrolyte leakage and MDA content decreased with increasing concentration of trehalose.
Thus may be due to multiple functions of trehalose, similar to proline, such as stabilizing enzymes, proteins and lipid membranes, preventing protein aggregation, protecting biological structures and scavenging free radical, numerous studies showed that trehalose involves in the acquisition of multiple stress tolerance [28].
The beneficial roles of Tre in drought affected wheat plants were observed as those treatments reduced oxidative stress (decreased H2O2 and MDA levels), compared to Tre untreated drought affected plants. Exogenous Tre application was also effective in reducing oxidative stress of other plants in different abiotic stresses [62,63].
Fortunately, plants have developed various protective mechanisms such as enzymatic antioxidant system to eliminate or reduce ROS which are effective at different levels of stress-induced deterioration. Antioxidants can prevent the oxidation of metabolic compounds by assisting in the elimination of free radicals generated by metabolism [64]. The antioxidant defense machinery that protects plants against oxidative stress damages includes both enzymatic and non-enzymatic antioxidant defense systems that work in concert to control the cascades of uncontrolled oxidation and protect the cells from oxidative damage by scavenging the ROS [65]. The results revealed that water stress showed that, water stress induced marked increase in AAO, POD and PAL activities and induced a non-significant reduction in PPO activity in flag leaves of both wheat cultivars during grain-filling comparing with the control plants. Among cultivars, tolerant one showed higher enzymes activity than sensitive one. Similar responses to stress conditions were reported in marigold [66]. In addition, POD is one of the major systems for the enzymatic removal of H2O2 in plants [67]. Since the H2O2 scavenging enzymes, peroxidase, located in both cytosol and chloroplasts can remove H2O2 efficiency. The same results were obtained by Pompelli et al. [68] who reported increased peroxidase activity in response to drought stress. A correlation between the antioxidant enzymes activity and drought tolerance was demonstrated by comparison of tolerant cultivar (Sahel-1) with sensitive one (Gemmieza-7) where higher activity of ROS-scavenging enzymes have been reported in Sahel-1 when compared to Gemmieza-7, suggesting that the antioxidant system plays an important role in plant tolerance against water stress.
The used wheat cultivars in this study showed differential responses in the activity of the three measured enzymes. It seems that the scavenging system depended often on ascorbic acid oxidase (AAO), peroxidase (POD), polyphenol oxidase (PPO) and phenylalanine ammonia lyase (PAL) activities. Hence the activity of these enzymes might offer considerable protection against the water-stress and the level of activity could be used effectively as an index of the severity of the stress. On the basis of our data, the antioxidative response is well correlated with growth sensitivity and tolerance of cultivars to water stress and we could conclude that, Sahel-1 could induce antioxidative system more efficiently than Gemmieza-7, resulting in slow growth suppression and lipid peroxidation under drought stress.
Application of SA and/or Tre markedly increased AAO, POD and PAL activities and a non-significant decrease in PPO activity in flag leaf of water stressed wheat plants. Generally, SA and Tre treatment appeared to be the most effective treatment in counteracting the negative effects of water stress on AAO, POD, PAL and PPO activities. These results are similar to those obtained by Kumar et al. [69] who stated increased of enzyme activity with foliar application of SA. Moreover, treatment of drought stressed plants with exogenous SA was found to be effective in modulating both enzymatic and nonenzymatic components of antioxidant defense system [17]. Moreover, Noreen et al. [70] reported that SA stimulates the antioxidant capacity, so that the leaf peroxidase activity has increased. Positive correlation between leaf peroxidase and super oxide dismutase activities were observed in sunflower lines with root fresh weight and CO2 exchange system. SA increases abscissic acid (ABA) content under stress and maintains the reduction of harmful effects of stress on the plant [71] and causes plants to re-grow.
Application of Tre markedly increased POD and PAL activities and a non- significant decrease in PPO activity. Hence, Tre has been proposed to be a signaling molecule under abiotic stresses [28] which induce plants to speed up their rate of ROS production that sends signal to activate enzymatic antioxidants for ROS scavenging in order to counteract stress-associated oxidative stress.
Collectively, the enhanced antioxidant activities of wheat plants under the studied circumstances were in close parallelism with the enhanced accumulation of various antioxidant metabolites. Among the secondary metabolites recognized in wheat plants, polyphenolics may largely contribute to the great antioxidant capacity of these plants in either control or stressed habitats. The effect of water stress on increasing the phenolic content (flavonoids and total phenols) in wheat plants grown under water stress was proved in the present study. It was suggested that the existence of several hydroxyl groups bonded to an aromatic ring provides the molecule with the ability of donating proton to a radical, so acting as a possible chain-breaking molecule or antioxidant upon secondary oxidation [72].
In the present study, data revealed that drought stress increased the amount of total phenols and flavonoids. Flavonoids are one of the largest classes of plant phenolics performing different functions in plant system, including pigmentation and defense [73]. Intriguingly, the conditions leading to inactivation of antioxidant enzymes can also upregulate flavonoids biosynthesis, suggesting that flavonoids constitute a secondary ROS-scavenging system in plants exposed to prolonged stress [74]. Coinciding with the results obtained in the present study, Haghighi et al. [75] recorded that water stress enhanced the accumulation of flavonoids in Plantago ovata plants. Moreover, they postulated that this increment in flavonoid content might be due to the induction in enzymatic activity occurring under stress, thereby favoring the production of different flavonoid compounds. These nonstructural carbohydrates then tend to accumulate and thus trigger the synthesis of carbon-based defensive substances.
In accordance with these results, drought brought about marked increase in the total amount of phenolic compounds in pea and wheat plants as indicated by Alexieva et al. [76]. Similarly, drought increased the concentration of total phenols in the leaves of five tomato cultivars as recorded by Rodríguez et al. [77]. It is well known that many phenolics are stress-induced metabolites that accumulate in plant tissues after different abiotic and biotic stress stimuli. These metabolites may participate in reactive oxygen species (ROS) scavenging mainly through the antioxidative enzymes utilizing polyphenols as co-substrates [78]. In addition, an increase in the emission of red fluorescence correlated with high content of phenolic compounds under stress conditions revealed the ability of these substances to participate in the adaptation mechanisms of photosynthetic apparatus to water deficit stress [79].
Application of SA and/or Tre markedly increased total phenols and flavonoids contents. Generally, SA and Tre treatment appeared to be the most effective treatment in counteracting the negative effects of water stress on total phenols and flavonoids contents. Hence, SA and Tre has been proposed to be a signaling molecule which induce plants to speed up their rate of ROS production that sends signal to activate non-enzymatic antioxidants for ROS scavenging in order to counteract stress-associated oxidative stress.
Beside, phenolic the non-photosynthetic pigments investigated in the present study may contribute to the antioxidant activity of wheat plants. The increased antioxidant activity due to stress coincided with higher amount of anthocyanin, lycopene and β-carotene. Several antioxidant mechanisms may explain anthocyanins effect, including hydrogen donation, metal chelation and protein binding. Lycopene and β-carotene could react with free radicals, notably peroxyl radicals and singlet oxygen, which is the basis for their function as antioxidants [80].
Perusal of the results showed that water stressed wheat plants increased the amount of anthocyanin and lycopene. However, the same effect was noticed for β-carotene. Anthocyanin is involved in attraction of pollinators with a pronounced effect under defenses against environmental stresses like drought [81]. With respect to drought, plant tissues containing anthocyanin is usually rather tolerant to drought [82]. For example, a purple cultivar of pepper resists water stress better than the green cultivar [83]. The highly drought-tolerant resurrection plant contains anthocyanin at a level that is several-fold more during dehydration than at a hydrated stage. The proposed mechanism behind anthocyanin-enhanced drought resistance is that anthocyanin is able to stabilize water potential [82].
Phenylalanine ammonia-lyase (PAL) is a key enzyme in phenylpropanoid metabolism as it catalyzes the conversion of phenylalanine to trans-cinnamic acid, the first step in the biosynthesis of anthocyannin. Of particular importance, it was reported that PAL is significantly induced by abiotic stresses such as drought [84].
For instance, the results obtained by Sedghi et al. [66] showed that the concentration of carotenoids; namely lycopene and β-carotene, increased under water deficit in Calendula officinalis plants compared with the control plants. Moreover, drought stress caused an increment in the amount of both lycopene and β-carotene in tomato plants [85]. Increasing evidence suggests that carotenoids are required for plant response to dehydration stress [86].
Application SA and/or Tre induced additional increase in anthocyanin, lycopene and β-carotene contents. The effect was more pronounced with SA treatment in case of anthocyanin pigment and with SA and Tre treatment in case of lycopene and β-carotene pigment. These results signify the role of SA in regulating the drought response of plants and suggest that SA could be used as a potential growth regulator to improve plant growth, under water stress. Also, previous studies proved Tre as efficient protectant against drought stress [87].
In conclusion, our results indicate that the drought exposure imposed evident on membrane characteristics and enzymatic antioxidant as well as non-enzymatic antioxidant. Furthermore, one of the most apparent influences of water shortage on the studied wheat plants was the ill impact of stress on membrane characteristics, as revealed by higher lipid peroxidation, lower MSI and higher ML when compared with the unstressed synonyms. On the other hand, the exogenous application of salicylic acid (SA), trehalose (Tre) or their interaction appeared to mitigate this damage effect of drought with different magnitude through stimulation of the enzymatic and nonantioxidants. Moreover, key players involved in SA and Tre-induced drought tolerance needs to be clarified using proteomic and genomic approaches. Although some researchers have successfully manipulated trehalose accumulation in plants for crop improvement purposes; its roles in tolerances to diversified of stresses in relation to plant architecture, growth and metabolism still demand huge researches.