Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Review Article - (2018) Volume 9, Issue 2
The Extracellular matrix (ECM) is a vital trigger of molecular events in development, homeostasis, cell phenotypes, and evolution. Annotated genome databases and their comparative analyses enable the assessment of the most conservative representation of the ECM components among the Metazoa. Bivalves are widely distributed animals present in various ecological niches. They have shells, indirect developmental life cycles, and highly diverse adaptations and habitat reflected by their ECM repertoire. Only recently, researchers began to characterize the conservation among invertebrates and unique species-specific ECM components involved in biomineralization, development, and cell-cell communications. Here, I discuss the most recent data for the dynamic expression of ECM proteins based on transcriptome data during larval oyster development and in adult organs. Using developmental and cell biology approaches along with confocal microscopy, I showed the expression of certain ECM components in the larvae, adult tissues, and cell cultures. A similar methodology is required to understand the molecular mechanisms and dynamic ECM expression in organisms with very complex life cycles, metamorphosis, indirect development via adaptive larvae, and biomineralization during development.
Keywords: Evolution; ECM; Fibronectin; Integrins; Biomineralization; Development; Mollusc
ECM: Extracellular Matrix
Invertebrates constitute ~95% of all animals on Earth. They are highly diverse in terms of development, lifestyle, morphology, and ecology. The genomes of several different bilaterian organisms, including model animals such as mammals, worms, flies, sea urchins, and ascidians, indicate that they all share common extracellular matrix (ECM) components. ECM components play essential roles in supporting and strengthening the tissues and internal organs of adult animals and in developmental events [1]. Therefore the evolution of these components may predate the diversification of this clade [2]. Most ECM studies have focused on vertebrate species with annotated genomes. The study of ECM in invertebrates, then, is important in providing insight into the origins of ECM.
ECM components play essential roles in supporting and strengthening the tissues and internal organs of adult animals and in developmental events [1]. The origin of ECM is closely linked with the evolution of single cells into multicellular organisms. The proteinaceous network of ECM could provide structural support for cellular aggregation such as intercellular bridges, cellular adhesion, and the mechanical properties that are necessary for the proper functioning of multicellular organisms and tissues [3,4]. Environmental changes and morphological transformations in predators might increase the selection pressures for ECM evolution [5]. These effects may result in a wide variety of ECM components. Recent bioinformatic investigations have demonstrated a high degree of conservation of the core ECM component known as the adhesome. Apparently, the adhesome evolved during the major wave of biotic innovation that occurred with the origin of Metazoa. Adhesomes consist of ECM components, ECM proteases, and cell surface receptors, all of which are broadly conserved throughout the Metazoa [6]. Among them, collagens are the most extensively studied of all the adhesome molecules. ECM components conserved from sponges to humans include three clades of fibrillar collagens and basement membrane collagen IV [7].
The Homoscleromorpha sponges have a basement membrane structure consisting of laminin and perlecan as well as collagen IV, and all these molecules are conserved throughout the Metazoa [7,8]. Another ECM glycoprotein and adhesome component, fibrillin, is found in the medusa Podocoryne carnea [9,10]. Other ECM components found in vertebrates such as agrin, thrombospondin, and secreted protein acidic and rich in cysteine (SPARC)/osteonectin are also conserved in sponges [11]. Adhesion receptor families such as integrins, syndecan, glypican membrane proteoglycans, and cluster of differentiation 36 (CD36) are present in sponges and throughout the Metazoa [12-15]. Other ECM components (dystroglycan and discoidin domain receptors) originated later within the Metazoa [16]. All of these examples support the idea that ECM originated in metazoan ancestors. Nevertheless, there is as yet limited evidence as to how the ECM components, each with its own distinctive properties, interact with each other to achieve the appropriate functions.
Many marine invertebrates are characterized by the ability to synthesize ECM components together with exoskeletal structures of different structure and shape, which serves to protect internal soft body from environment and predators. Possibly, the reason of ECM mineralization was increasing in atmospheric oxygen and oceanic calcium levels. The first biocomposites (Complex of mineral and ECM components) were built in association with collagen fibrils and had remarkable mechanical properties. Such biomaterials appeared in many phyla between 535 million and 545 million years ago (dawn of the Phanerozoic period) and, possibly, independently in several group animals. Biomineralized structures were developed from SiO2. They are found in radiolarians, diatoms, sponges, boreal copepods, and brachiopod larvae. Ca3(PO4)2 appeared in brachiopods and vertebrates. CaCO3 was found in brachiopod and molluscan shells, coral skeletons, echinoderm spines and tests, and sponge spicules [17-19]. Beginning more than 500 million years ago, molluscs developed a wide variety of shells. Organic and non-organic compositions and biomineralization in bivalve shells have been relatively well studied [18-20]. Mollusc shells consist of 95% to 99% CaCO3 and the rest is organic matrix including proteins and polysaccharides secreted from outer epithelial cells in different regions of the mantle [21,22]. Moreover, granulocytic hemocytes are able to participate in shell regeneration, which supports the cellular hypothesis stating that CaCO3 and matrix proteins are formed in the hemocytes and transported to the mineralization area [23].
Previous studies have focused on changes in the genes involved in biomineralization. They concentrated on separate stages of mollusc development and provided only a fragmentary view of how gene expression changes during ontogenesis. Annotated genomes permitted the development of new tools that enable a more global view of the changes in gene expression that occur over the course of larval developmental [24,25]. Visualization of multi-omics data and transcriptome analysis may be the best approaches for obtaining solid information about non-model animal development and the role of matrix proteins in shell formation. The genetic sequences and gene expression profiles involved in biomineralization during development have been investigated in the adult pearl oyster P. fucata [26,27]. The expression of several matrix proteins (nacrein, N16, prismalin-14, aspein, MSI60, and MSI31) was analyzed by real-time PCR. It was confirmed that they participate in larval shell formation [26]. Miyazaki et al. [26] found that the expression of these six matrix genes started 18 h post-fertilization (hpf) (Trochophore stage), coinciding with the initiation of shell formation and growth. They followed the ongoing expression of the aforementioned genes up to 49 d post-fertilization (dpf). Later, Liu et al. [27] applied microarray analysis to the global gene expression profiles during P. fucata development. They observed that the genes involved in biomineralization, including those encoding nacrein, pearlin, Pif, amorphous calcium carbonate-binding protein (ACCBP), prisilkin-39, and the shematrin family, are highly upregulated in juveniles [27]. They also established that the genes encoding for chitin synthase and PFMG2 were highly expressed at the D-veliger stage, whereas PFMG6, PFMG8, and PfN23 were significantly upregulated at the umbo stage. Therefore, they play different roles in regulating shell formation. Moreover, they identified a new gene expressed in the mantle that encoded secreted proteins with tandemly arranged repeat units characteristic of most matrix proteins. RNAi knockdown disrupted the nacreous and prismatic shell layers, indicating the potential roles of these proteins in shell formation [27]. Feng et al. [18] performed a BLASTp bioinformatics analysis using the shell matrix proteome from the oyster C. gigas . The reference database consisted of 443 shell matrix proteins from nine other species. They identified and characterized shell protein homologs, chitin related proteins, enzymes (tyrosinase, peroxidase), and ECM-related proteins (Pif, BMSP, EGF-ZP, and SPARC) [18]. Nevertheless, the onset and transformation of ECM and bivalve adhesome components during mollusc development and expression in adult tissues remain unclear.
Adult tissues
The knowledge of molecular ECM composition in mollusc taxa is limited. ECM genes are highly expressed in the mantle (outer and inner parts), digestive gland, female gonad, gills, adductor muscle, hemocytes, and labial palps [25]. Additionally, the annotated oyster genome and the predicted proteins of conservative adhesome components suggested high expression of syndecan, collagen IV, and fibrillar collagen, and SPARC in the oyster mantle [25]. In general, the expression of the ECM components in the inner mantle was slightly lower than that in the outer mantle and very low in the digestive glands. Syndecan and SPARC were highly expressed in the female gonads. Fibrillar collagen, perlecan, and SPARC were all highly expressed in the gills, whereas the expression of collagen IV and MMP was elevated in the adductor muscles. Hemocytes highly expressed SPARC, integrins, and syndecan. In the labial palp organ, SPARC and syndecan were highly expressed [25]. These data for the RNA expression of ECM components are important for understanding ECM composition in molluscs and the development of ecological markers for testing healthy tissues of molluscs. An immunohistochemical study of the subepidermal connective tissue in the adult molluscan integument showed that the ECM is similar in gastropods, bivalves, and cephalopods, but differs from that of vertebrate dermis. In Pecten and Mytilus species, immunostaining for I-like collagen was particularly conspicuous in the areas surrounding muscles and glands [28]. Collagen V was intensely positive in subepidermal connective tissues of bivalve Pecten, whereas in Mytilus the staining particularly involved the most superficial part of the subepidermal tissues. Positive immunostaining for collagen VI was detected in the integument of the bivalve Pecten and Mytilus , whereas staining for collagen IV revealed an intensely positive laminar structure under the epithelium. Laminin positivity was observed immediately beneath the epithelium of all bivalve sections examined. In all bivalves, anti-nidogen immunoreactivity was always localized in the immediate sub-epithelial zone and around other structures within the subepidermal connective tissues. The sub-epithelial zone of all bivalves studied was positive for heparan sulfate proteoglycan. The molluscan integumental ECM contains collagens resembling types I, V, and VI, but has no collagen similar to type III. Vertebrate laminin, nidogen, and heparan sulfate were present in many molluscan basement membranes. Therefore, basement membrane structure and composition must have remained constant throughout the evolution of all animal phyla [28].
Among ECM components, integrins are one of the families of cell adhesion receptors, and are multifunctional regulators of development in vertebrates and invertebrates [29,30]. Integrins are heterodimers consisting of associated α and β subunits, and the last subunit is the most conserved in vertebrate and invertebrate animals. Despite the fact that integrins (α - and β chains) are well known to exist in all metazoan, its presumptive ligand, fibronectin (FN, as fibronectin-like protein, FN-LP) evolutionary originate as novel ECM components in Urochordate species C. intestinalis , C. savignyi [2,31]. In contrast to these data, recent genomic and transcriptomic data of bivalves indicate that FN-like and FN III-containing domains exist in genome of bivalves [25,27]. Moreover, the immuno detection of a β integrin-like protein (β integrin-LP) and FN-LP was determined in the bivalve mollusc M. trossulus [32]. Double (integrin/fibronectin) immunofluorescence labelling of adult tissues showed β integrin-LP in the hepatopancreas/stomach, adductor muscle, and male gonad. In the case of the male Mytilus , the integrin immunoreactivity was strongly observed in the gonad and gill integument. Integrin-like-positive cells (probably hemocytes) were found in the adult mussel mantle (Figure 1). Fibronectin immunoreactivity was not detected in the male gonad [32], but was spatially distributed in other adult mussel tissues from the intracellular (cytoplasmic) localization of the hepatopancreas/ stomach and adductor muscle to an extracellular location, with the formation of FN-like positive fibers in the gill and mantle (Figure 1) [32]. Possibly, in addition to the immunopositive basal membranes, the antibodies against integrin recognized different subpopulations of M. trossulus hemocytes in tissues. These results indicate that antibodies against β1 integrin specifically recognize the mussel target proteins and can be used for their detection in adult mussel tissues and could be useful for cell biological studies [32]. Despite that FN antibodies recognize the basal membranes and hemocytes in adult bivalves, the confirmatory foradditional proof of cross-reaction properties of FN are needed in future.
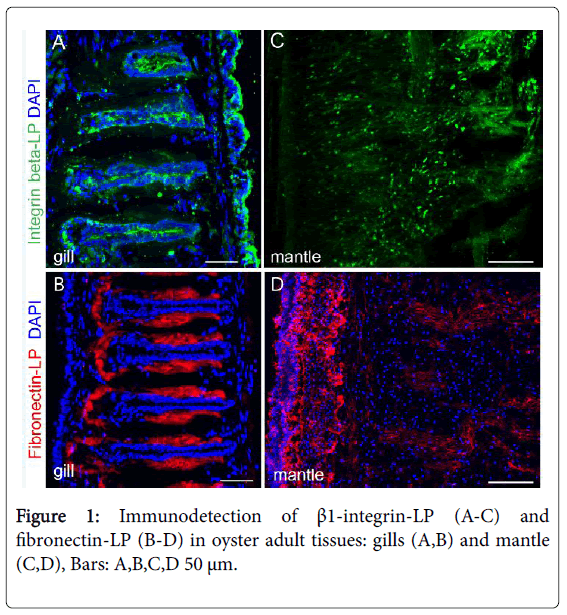
Figure 1: Immunodetection of β1-integrin-LP (A-C) and fibronectin-LP (B-D) in oyster adult tissues: gills (A,B) and mantle (C,D), Bars: A,B,C,D 50 μm.
Most bivalves have a biphasic life cycle that consists of actively swimming planktonic larval stages (from a ciliated trochophore to a pediveliger larva) and sedentary benthic adult animals (Figure 2). At the late trochophore or early veliger stage, the shell gland cells begin secretion [33]. The bivalve mollusc shell is ectodermal in origin. The cellular mechanisms are well described and are roughly divided into the following phases: (a) Establishment of the shell-founder cell; (b) Invagination then evagination of the shell field; (c) Successive expansion of the shell field to cover the entire embryo at the veliger stage; and (d) Secretion of the shell matrices and calcification of the shell field [34]. The oyster C. gigas is an interesting model for developmental biology and its single annotated bivalve genome is available (http://www.oysterdb.com). Zhang et al. [25] annotated 28,027 genes from 38 developmental stages (egg to juvenile) of C. gigas . They posted their data on the Oyster Genes website (http:// www.oysterdb.com). The analysis of gene expression in several developmental stages revealed that many of the predicted proteins are involved in shell formation and adaptation to environmental stress. For example, genes encoding several adhesome components from the annotated oyster genome database were published. These were found in non-metazoan and metazoan [6,7]. Expression of the main ECM components, including interstitial/connective ECM elements, basement membrane components, ECM receptors, and certain matrix metalloproteinases, was identified in the Oyster Genes website.
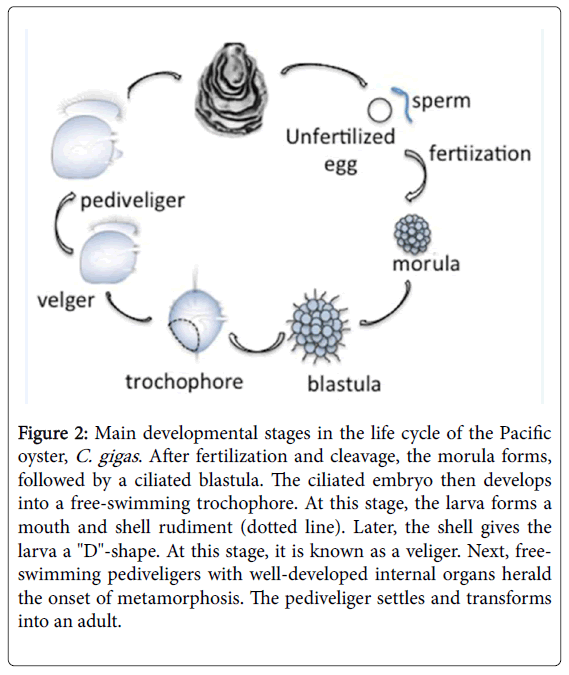
Figure 2: Main developmental stages in the life cycle of the Pacific oyster, C. gigas . After fertilization and cleavage, the morula forms, followed by a ciliated blastula. The ciliated embryo then develops into a free-swimming trochophore. At this stage, the larva forms a mouth and shell rudiment (dotted line). Later, the shell gives the larva a "D"-shape. At this stage, it is known as a veliger. Next, freeswimming pediveligers with well-developed internal organs herald the onset of metamorphosis. The pediveliger settles and transforms into an adult.
Among the most highly conserved metazoan components of interstitial/connective ECM, the expression of fibrillar collagen-LP (CGI_10007860) and fibrillin-LP (CGI_10001572) starts to increase at later stages of oyster development. The matricellular protein SPARCLP (Secreted Protein Acidic and Rich in Cysteine, CGI_10005088) has a low expression level in D-veliger larvae but its expression increases by the umbo and pediveliger larval stages. Matrix metalloproteinases (MPP-LP, CGI_10025346), disintegrin, and metalloprotease (ADAMLP, CGI_10027133) have low expression levels throughout oyster development. In silico-predicted basement membrane components laminins (α: CGI_10026821; β: CGI_10005310), collagen-LP IV (CGI_10025803) are highly expressed during the early stages of oyster development. The basement membrane proteoglycans, namely a large agrin-LP (CGI_10007223) and heparan sulfate proteoglycan (perlecan- LP, CGI_10025660), are expressed differently from other genes in oyster development. Perlecan-LP is expressed at very low levels in the gastrula stage and remains expressed at low levels until late stages (spat and juvenile). Proteoglycan agrin-LP is expressed in the late D-veliger stage and reaches maximum expression at the umbo stage. The expression of the basement membrane component nidogen-LP (CGI_10007863) does not significantly change throughout oyster development.
The ECM receptor glypican-LP (CGI_10016108) is a major heparan sulfate proteoglycan and together with syndecan-LP (CGI_10004071) is highly expressed from the early stages until the early trochophore stage. Another ECM receptor, dystroglycan (CGI_10020032), is one of the dystrophin-associated glycoproteins. Its expression starts from the four-cell embryo stage, reaches a maximum at the Trochophore stage, and decreases during the veliger stage. Fibronectin-LP (CGI_10016964) expression starts at the gastrula stage of oyster and reaches its first peak at the trochophore stage, followed by another maximum at the D-veliger stage. Integrin-LPs are the most heterogeneously expressed during oyster development. Most of the α and β-chains are expressed at early stages of oyster development, and all integrins are highly expressed at the juvenile stage. The reason of non-uniform expression of integrin isoforms could be explained by a tissue specific expression of proteins during development and their unique role in the formation of basal membranes and receptor-ligand interactions involving in growth and differentiation processes.
The ECM and its receptors make various contributions to development. There are many similarities between invertebrate and vertebrate ECM components. However, certain ECM components are unique to invertebrates, such as the cuticlins in C. elegans , tiggrins in D. melanogaster , and shell matrix proteins in bivalves. Immunodetection was not performed for the aforementioned bivalve ECM components because specific antibodies were lacking for them, and the bivalve larvae were small and difficult to cultivate. Nevertheless, a few studies detected certain components of the ECM in bivalve development and in vitro [32,35,36].
In our previous study, we evaluated the localization of a β-integrinlike protein (β-integrin-LP) during the early development of the bivalve M. trossulus [32]. The first β1-like integrin-positive signal appeared at the early trochophore stage (19 hpf) in 5% of the Mytilus larvae. Later, at the Trochophore stage (24 hpf), β1-like integrin was detectable in the developing stomach area. β1-like integrin was localized on the membranes of the separate digestive mass cells forming the gastric lumen. The hepatopancreatic β1-like integrinimmunopositive antigen became more visible at the veliger stage (51 hpf) because the larval stomach was well-formed by that time. Another component of ECM is Fibronectin (FN), which is one of the most important ligands for β1-integrins found in bivalve genome [25,27]. It binds an arginine-glycine-aspartic acid tripeptide and regulates vertebrate cell morphogenesis [37]. We detected FN-LP in the oyster blastula stage (12 hpf). The signal intensity increased sharply during the trochophore (30 hpf) and veliger (56 hpf) stages. The β1-like integrin oyster larval cells do not express FN-LP. We propose, therefore, that FN-LP and β1-like integrin do not interact with each other either during M. trossulus development or in adult tissues. For this reason, FN-LP is not a relevant β1-like integrin ligand [32]. The analysis of the expression of FN-LP and integrin β-LP is summarized in Figure 3.
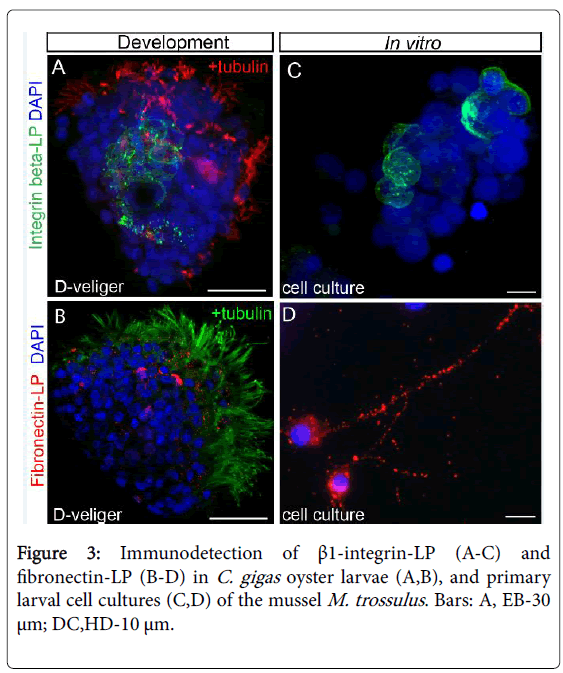
Figure 3: Immunodetection of β1-integrin-LP (A-C) and fibronectin-LP (B-D) in C. gigas oyster larvae (A,B), and primary larval cell cultures (C,D) of the mussel M. trossulus . Bars: A, EB-30 μm; DC,HD-10 μm.
In vitro studies
Immunodetection analysis of bivalve larval ECM components and primary cell cultures from the Embryo (blastula) or Trochophore (larva) stage are very useful approaches for investigating ECM components in cellular morphogenesis. Previously, we developed a molluscan in vitro technique that allows us to study cellular differentiation (Myogenic, Neurogenic, and Ciliary differentiation) [36]. We also demonstrated that ECM components (Fibronectin, poly- L-lysine, and carbon) may promote cell differentiation and proliferation in primary cell cultures of the mussel M. trossulus . These ECM components may even support contractile activity in cell culture. Larval mussel cells raised on collagen I attached to the substrate but did not spread. These larval cells inhibit myo-differentiation and cell specialization in general. In contrast, larval cells that were spread on coverslips coated with fibronectin, poly-L-lysine, or carbon were, in fact, able to differentiate into muscle cells (Figure 4) [35].
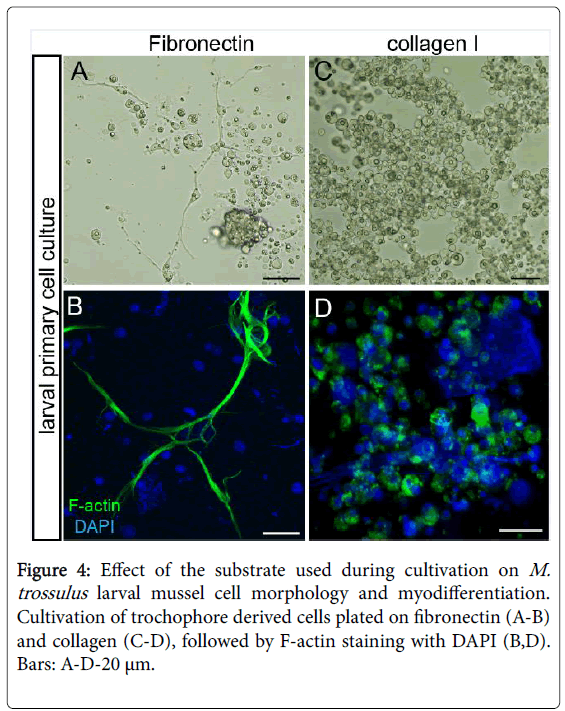
Figure 4: Effect of the substrate used during cultivation on M. trossulus larval mussel cell morphology and myodifferentiation. Cultivation of trochophore derived cells plated on fibronectin (A-B) and collagen (C-D), followed by F-actin staining with DAPI (B,D). Bars: A-D-20 μm.
Larval mussel M. trossulus cells. They begin to express musclespecific proteins and assemble into sarcomere-like structures [36]. These observations suggest that the inhibition of larval cell myodifferentiation by collagen is a reversible process. Moreover, the differentiation mechanisms can be regulated by ECM components even in highly deterministic animals such as the Mollusca. Therefore, the differentiation and proliferation of embryonic and larval mussel (M. trossulus ) cells may be regulated by individual ECM components isolated from vertebrates. The effects of substrates isolated from vertebrates on mollusc larval cell morphology are well described. These matrices were found to be suitable for cell attachment, differentiation, and proliferation in vitro [36,38]. Nevertheless, the cultivation of embryonic and larval bivalve cells on ECM components does not necessarily generate long-term cell lines. For this purpose, then, it is preferable to use ECM components isolated from invertebrates in general and molluscs in particular. The investigation of ECM synthesis in tissue and primary cells cultures is also a future research priority [39-42].
The author reports no conflict of interest.
I wish to thank Maria Maiorova for her assistance with the confocal microscopy, and Dr. Ekaterina Selivanova and Dr. Svetlana Goncharova for moral support. This work was supported by the Russian Science Foundation (Grant No. 14-50-00034).