Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2012) Volume 1, Issue 4
Cunila incisa is a perennial shrub with 1-2 m high with ramified branches. This highly aromatic species is commonly used in south brazilian folk medicine, growing wild at the margins of the forest in the highlands. Based on the medicinal and aromatic potential of this species the aim of this survey was to examine the genetic variation within and among populations. In this context, 13 C. incisa populations collected in Rio Grande do Sul, Brazil, were analyzed by RAPD.Cluster analysis afforded to discriminate populations under three groups. The C. incisa populations presented low intrapopulational variability, indicating that the populations form distinct genetic pools possible due to founder effects and low interpopulation gene flow.
Keywords: Cunila incisa; Lamiaceae; Genetic variability; Low gene flow; Founder effect
The genus Cunila D. Royen ex L. belongs to the Lamiaceae family (Nepetoideae; Mentheae) and has two geographic distribution centers, one in North America, and another in southern South America [1,2]. Based on growth habit and inflorescence morphology, Epling (1936) classified the South American species into three sections: Incanae, Incisae and Spicatae. Recently, studies started to address the taxonomical review of Cunila, indicating only two sections recognized for this genus: Spicatae and Incanae/Incisae [3].
Cunila incisa Benth. belongs to the section Incisae, and is classified as a perennial shrub with 1 - 2m high with ramified branches and ovoid to rhomboid leaves with incised margins. The small flowers are grouped in cimules fixed to the branches by short peduncles. Leaves, flowers and stalk are covered by glandular trichomes [4]. This highly aromatic species, which grows wild at the margins of the forest in the highlands (300 – 1000 m), shoots flowers between September and May [2].
Hydrodistillation of the aerial parts of Cunila incisa yielded 1 to 1.3% (w/w) of an essential oil with high content of 1.8-cineole (50-60%) [4,5]. This compound held responsible for the species’ secretolytic, secretomotoric, bronchospasmolytic, antiphlogistic, and antiseptic properties [6], that justify the popular application of this plant for the treatment of chronic cough and respiratory diseases [7]. Significant variation in the concentration of volatile compounds was detected within and among Cunila incisa populations, collected over the distribution area of the species in Rio Grande do Sul State (Brazil). Most populations exhibited low intra-populational variation indicating founder effect [5].
Considering the aromatic and medicinal potential of C. incisa, the present study aims to evaluate the genetic variability among plants collected in its native environment.
Plant material
Five plant samples were collected from each of 13 Cunila incisa populations (65 plant samples) in field excursions to different locations in the state of Rio Grande do Sul, Brazil (Figure 1). One sample belonging to C. incana, and one to C. angustifolia was used as out group.
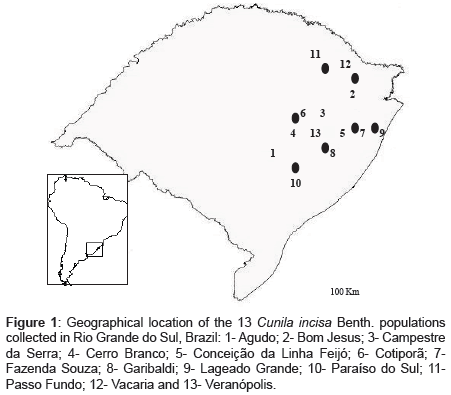
Figure 1: Geographical location of the 13 Cunila incisa Benth. populations collected in Rio Grande do Sul, Brazil: 1- Agudo; 2- Bom Jesus; 3- Campestre da Serra; 4- Cerro Branco; 5- Conceição da Linha Feijó; 6- Cotiporã; 7- Fazenda Souza; 8- Garibaldi; 9- Lageado Grande; 10- Paraíso do Sul; 11- Passo Fundo; 12- Vacaria and 13- Veranópolis.
DNA extraction and pcr amplification
Equal amounts (0.1 g dry weight) of leaf tissue from each sample were placed in porcelain mortars refrigerated with liquid nitrogen and ground to a fine powder. Total genomic DNA was extracted according to the CTAB method described by Doyle and Doyle [8]. DNA samples were spectrophotometrically quantified at 260 nm.
RAPD-PCR amplifications were performed in a total reaction volume of 25 μl containing 10 mM Tris-HCl (pH 8.3), 50 mM KCl, 1.25 mM each dNTP, 3 mM MgCl2, 30 ng of primer, 1.5 unit of Taq polymerase (Invitrogen), 0.25% of Triton-X-100 and 50 ng of genomic DNA. The amplification conditions were one initial 5-min step at 92°C followed by 40 cycles of 45 s at 92°C, 1 min 40 s at 40°C and 2 min at 72°C. The reactions were completed by a final extension step of 5 min at 72°C. DNA amplifications were performed using a thermocycler (MJ Research PT 100, Watertown, Mass.).
The amplification products were separated by electrophoresis in 1.5% agarose gel in 1X TBE buffer (50 mM Tris, 50 mM boric acid, 2.5 mM EDTA, pH 8.3). The size of the amplified products was determined by comparison with lambda DNA digested with EcoRI and HindIII restriction enzymes.
For DNA amplification, initially 60 random primers of the OPA, OPC, and OPZ kits of Operon Technologies (Alameda, California) were tested. The pattern of amplifications was screened for the number of bands, band intensity, and reproducibility, and allows to select 14 primers to perform the study (Table 1).
| Primer | Sequence (5’>3’) | No. of Bands | Polymorphic Bands (%) Including Outgroup | Polymorphic Bands (%) Excluding Outgroup |
|---|---|---|---|---|
| OPZ 03 | CAGCACCGCA | 8 | 75 | 37.5 |
| OPZ 05 | TCCCATGCTG | 3 | 33.3 | 0 |
| OPZ 13 | GAGCACCGCA | 15 | 86.6 | 46.6 |
| OPZ 16 | TCCCCATCAC | 11 | 100 | 63.6 |
| OPZ 17 | CCTTCCCACT | 8 | 100 | 37.5 |
| OPA 01 | CAGGCCCTTC | 15 | 100 | 53.3 |
| OPA03 | AGTCAGCCAC | 13 | 84.6 | 15.4 |
| OPA 10 | GTAATCGCAG | 4 | 75 | 50 |
| OPC 02 | GTGAGGCGTC | 16 | 81.2 | 25 |
| OPC 06 | GAACGGACTC | 16 | 100 | 62.5 |
| OPC 08 | TGGACCGGTG | 9 | 77.8 | 66.7 |
| OPC 11 | AAAGCTGCGG | 4 | 75 | 25 |
| OPC 12 | TGTCATCCCC | 11 | 100 | 27.3 |
| OPC 15 | GACGGATCAG | 14 | 92.8 | 42.8 |
| Total | 147 | 89.1 | 42.2 |
Table 1: RAPD primers with respective number of bands and polymorphic bands.
Statistical analyses
Bands were scored as a binary variable, (1) for presence and (0) for absence. The binary matrix (1/0) was used to calculate the Jaccard’s similarity coefficient between each pair of samples. The relationships among and within populations were evaluated by constructing dendrograms using the UPGMA (Unweighted Pair Group Method Using Arithmetic Averages) algorithm and the analysis of principal components assisted by the NTSYS Package [9] and SPSS software (SPSS Inc., Chicago).
Sixty primers were previously tested for RAPD amplification, but only 14 primers were selected for further analyses, based on the quality of profiles. The selected primers generated 147 amplified fragments, 89.1% of which were polymorphic, including outgroup. Considering only C. incisa samples the polymorphism reached 42.2% (Table 1). The number of amplicons per primer varied from three to 16 (Table 1) and size ranged from 150 to 1400-bp. The amplification pattern revealed the existence of species-specific markers for the three species analyzed, being 14 specific bands for C. incana, 17 bands for C. angustifolia and 11 specific bands shared by all populations representing C. incisa. The analyses allowed discriminating all C. incisa populations conferring to RAPD markers great power in the genotype identification of this species.
Genetic proximity inferred from Jaccard’s similarity index between C. incisa and C. incana was 0.305, between C. incisa and C. angustifolia was 0.370 and at last between C. incana and C. angustifolia was 0.224. The interspecific genetic similarity was in agreement with that found based on ISSR markers [3] and with the first classification made by [10] in which C. incisa and C. angustifolia belong to section Incisae and C. incana belongs to section Incanae. Genetic similarity calculated for each pair of populations varied from 0.801 to 0.98. The highest similarity value between populations was detected for the comparison between Fazenda Souza and Conceição da Linha Feijó populations (0.98), both of which are located at Caxias do Sul region and separated each other by 20 km. High genetic similarity within other Cunila species were previously reported using RAPD [11] and ISSR markers [3,12,13]. Following those authors, the high similarity found may be associated to restricted genetic bases reflecting founder effect and low gene flow among populations.
Cluster analyses afforded to discriminate populations under three groups (Figure 2). Group 1 was formed by populations from Fazenda Souza, Conceição da Linha Feijó, Veranópolis, Cotiporã, Lageado Grande, Vacaria, Bom Jesus, Garibaldi and Agudo. Excepting for Agudo, located in central Rio Grande do Sul state, all the populations are situated in the northeastern mountains of Rio Grande do Sul state and in the high grasslands of the Atlantic range. The second group was formed by two populations, Paraíso do Sul and Cerro Branco, both native to central regions of Rio Grande do Sul state and showing high similarity in the essential oil’s chemical composition [5]. At last, Group III was formed by the population native to Passo Fundo, located in the Rio Grande do Sul plateau, and by the population representative of Campestre da Serra, situated in the border of highlands.
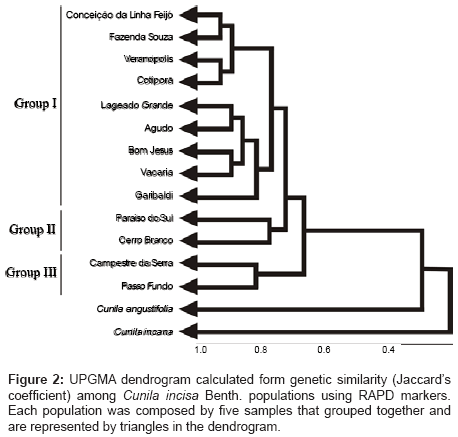
Figure 2: UPGMA dendrogram calculated form genetic similarity (Jaccard’s coefficient) among Cunila incisa Benth. populations using RAPD markers. Each population was composed by five samples that grouped together and are represented by triangles in the dendrogram.
A low but positive and significant correlation (p > 0.01) was found between the distances obtained based on the essential oil analysis [5] and the similarity index calculated based on RAPD markers. This correlation suggests that the essential oil composition observed in C. incisa is under genetic control but quantitative variations are under environmental control and that the C. incisa populations present individual genetic and chemical characteristics, possible due to founder effects and low interpopulation gene flow. Similar results were obtained by [14] analyzing the chemical and genetic variation among different C. galioides populations. For C. galioides, different chemotypes were found, fact that can influence the pollination process and explain the chemical, geographical and genetic correlation observed [14]. However, in C. incisa, no chemotypes were found and the quantitative variation in the chemical composition observed is probably related to environmental conditions [5].
Considering the protandry, low rate of fertilization, and pollination by insects, the reduced gene flow among populations appears to be a characteristic of the genus Cunila. This fact may be due to the discontinuous distribution of the species, which usually occur in very specific environments, associated with the short distance reached by insects that act as pollinators of these species.
The authors are grateful to the Universidade de Caxias do Sul for financial support and permission to use its facilities, and CAPES for the fellowship provided to G. Agostini.