Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2016) Volume 4, Issue 2
Plant polyphenols quercetin and naringenin are considered healthy dietary compounds; however, little is known of their effect on the probiotic Lactobacillus rhamnosus GG (LGG). In this study it was discovered that both quercetin and naringenin produced temporary inhibition of LGG growth, particularly at 8 hours post inoculation, with LGG eventually recovering from this suppression. The observed growth inhibition was regarded as a phenotypic response of LGG to the polyphenols; we hypothesized that the subsequent recovery was due to unknown, underlying genetic factors. The molecular response of LGG to quercetin and naringenin was determined through RNA analysis using the Helicos single molecule sequencing platform. The expression profiles of LGG grown in the presence of either quercetin or naringenin were divergent from each other, with only a few similarities, indicating that these polyphenols inhibit growth through separate mechanisms. LGG treated with quercetin demonstrated upregulation of genes associated with DNA repair and transcriptional regulation, and a decrease in expression of genes involved in metabolism and protein movement through the cell wall. LGG treated with naringenin resulted in an increase of genes associated with metabolism, and a decrease in genes involved in stress response. Results from this study demonstrate that there is a clear interaction between the polyphenols quercetin and naringenin and the probiotic LGG. The RNA expression analysis provides unique insight into the molecular response of LGG to quercetin and naringenin, revealing an identifiable pattern of gene expression.
Keywords: Lactobacillus rhamnosus GG; LGG; Quercetin; Naringenin; Polyphenols; Probiotics; RNA single molecule sequencing; Helicos; Genetic expression analysis
Quercetin and naringenin are among the most studied plant polyphenols that exert beneficial effects on human health [1-4]. The antioxidant properties and radical scavenging activity of the polyphenols are the major determinants that have led research on their applications as potential therapeutics for the protection and treatment of cancers [5,6] and many chronic diseases, including cardiovascular disease [7], diabetes [8,9], and obesity [10,11]. However, research has also shown that polyphenols exhibit pro-oxidant behavior, and could be a carcinogen or mutagen [12-18]. It has been reported that factors such as pH value of media, the presence of transition metals, or enzymes [19-22], and, particularly, the complexity of each individual’s digestive system [23-25] have profound impact on these contradictory results.
The interaction of polyphenols with the intestinal microbiota has attracted increasing attention in regards to their impact on health. Quercetin and naringenin have lipophilic character; they are easily absorbed in the stomach. However, significant amounts of quercetin and naringenin are rarely found in the diet. Polyphenols in the diet mainly occur in glycosylated forms [25,26]. For example, quercetin 3- O-glucoside in onions, quercetin 3-O-rhamnoside in onions and cranberries, quercetin 3-O-rutinoside in buckwheat leaves, or naringenin in grapes and citrus fruits. The quercetin derivatives, containing a substituent other than glucose, are not hydrolyzed by endogenous human enzymes in the small intestine. Most polyphenols pass through the stomach and small intestine. They enter the cecum and colon where they are hydrolyzed by colonic microbiota to aglycone and sugar (glycone), and are further degraded into phenolic acids, or undergo other structural modifications to various derivatives. Knowledge of the interaction between polyphenols and the gut microbial community is still limited. Some of these reactions and their metabolites may insert beneficial effects, some may not. It has been reasonably questioned whether or not polyphenols, or the metabolites produced by gut microbial digestion of polyphenols, in the colon could potentially inhibit the growth of some species of bacteria, including beneficial microbiota, as well as alter the diversity and disturb the balance, or equilibrium, of the gut microbiota community [27-29].
It has been reported that polyphenols and phenolic acids exhibit inhibitory activity against bacteria, either Gram-negative or Grampositive, such as Fusobacterium nucleatum [30], Escherichia. Coli [31], and Listeria monocytogenes [32], etc. It has also been reported that polyphenols may have no impact on the growth of some bacteria, such as wine extracts that showed no effect on Lactobacillus spp., Enterococcus spp., and Bifidobacterium spp. [33], or even enhance bacterial growth. For example, tea extracts on probiotics Lactobacillus casei strain Shirota, Lactobacillus Rhamnosus GG (LGG), and Bifidobacterium [34], as well as quercetin or naringenin on the growth of Bifidobacterium [27]. These results were based on the observation of cell proliferation and/or instrumental analysis of metabolites. Therefore, the investigation on polyphenols vs. a bacterial species in a one-to-one simple system could provide significant insight of the interactions occurring in gastrointestinal tract.
Lactobacillus rhamnosus GG, known as LGG, (ATCC 53103) is a probiotic, isolated from the fecal sample of a healthy human being. The metabolite of LGG, protein P40, has been demonstrated effective in the prevention of inflammatory bowel disease, such as Crohn’s disease and ulcerative colitis, and other several clinical applications, and is a prevalent constituent in functional foods [35-37]. Therefore, a better understanding on the possible interaction between polyphenols and LGG is of importance, especially if the polyphenols adversely affect critical cellular functions or overall growth.
In the current study, quercetin and naringenin were used to evaluate the effects of polyphenols on LGG. LGG cultured in the presence of quercetin or naringenin were monitored over time in order to detect changes in cell growth. Alterations in growth due to the presence of either polyphenol provided the phenotypic response of LGG to quercetin and naringenin. In order to analyze the genotypic response, Helicos single molecule sequencing was used to generate the RNA expression profiles of LGG grown in the presence of either quercetin or naringenin. Analysis of the expression profiles revealed a pattern of change in genetic regulation in response to quercetin and naringenin. These results provide a more multi-dimensional understanding of how polyphenols may impact the growth of probiotics, in particular LGG.
Preparation of broth and recovery of Lactobacillus rhamnosus GG
Lactobacillus rhamnosus GG (LGG) was acquired from ATCC (53103), cultured and grown in Difco Lactobacilli MRS broth purchased from Becton, Dickinson and Company. Prior to use, the MRS broth was autoclaved under pressure at 12°C for 30 minutes. After autoclaving, the broth was boiled under negative pressure using Nitrogen gas for 10 minutes to remove any oxygen, transferred into a Bactron anaerobic chamber and allowed to cool overnight. Prior to experimental use, this broth was aliquoted into hungate tubes purchased from Chemglass, 5 mls per tube, and stored in the anaerobic chamber at room temperature. Frozen aliquots of LGG were recovered through inoculation in anaerobic Lactobacilli MRS broth and grown overnight in the anaerobic incubator at 37°C. The bacteria were cultured at least two times in sequence prior to experimental use to ensure proper recovery from freezing.
The polyphenols quercetin and naringenin
Naringenin (W530098-500G) and quercetin (Q4951-100G) were purchased from Sigma-Aldrich and dissolved in DMSO to make stock solutions (made fresh for each experiment). The DMSO stock solutions containing either quercetin or naringenin were added to the anaerobic Lactobacilli MRS broth at a volume of 10 μl DMSO stock solution per 5 ml of Lactobacilli MRS broth (final cultures contained 1% DMSO).
For the initial experiment, quercetin was tested at a final concentration of 12.5, 2, 50 and 75 μg/mL and naringenin was tested at a final concentration of 50, 100, 150 and 200 μg/mL. Regarding cultures sent out for RNA sequencing, quercetin was used at a final concentration of 50 μg/mL and naringenin was used at a final concentration of 100 μg/mL. For each concentration tested, the polyphenol added to the Lactobacilli MRS broth without addition of LGG was used as a negative control. LGG cultured in Lactobacilli MRS broth containing DMSO without polyphenol was used as a positive control.
LGG and the polyphenols quercetin and naringenin:
All work for this experiment was performed in a Bactron anaerobic chamber. Anaerobic Lactobacilli MRS broth was aliquoted into hungate tubes the day before starting the experiment, 5 mLs per tube. The tubes were sealed with a rubber septa and screw cap lid, and left overnight in the anaerobic chamber. LGG was grown to confluency by inoculating 5 mLs of Lactobacilli MRS broth and incubating at 37°C for 16 hours under anaerobic conditions.
At the start of the experiment, each hungate tube containing prealiquoted Lactobacillus MRS broth was injected with DMSO containing either quercetin or naringenin to the desired final concentration (1 mL needle and 25 gauge syringe). The confluent culture of LGG was diluted to 0.5 Macfarland units (MU) over the media only read. 100 μLs of this culture was injected into each 5 mL hungate tubes containing Lactobacilli MRS broth and the desired concentration of polyphenol using a 1 mL needle and a 25 gauge syringe. After injection, each tube was briefly vortexed to ensure dispersion and the Macfarland units for each culture were determined using a densitometer (time 0 read). The cultures were then placed into the anaerobic incubator set to 37 C. At, 8, 12 and 24 hours post inoculation, each culture was removed from the incubator, briefly vortexed to ensure proper dispersion and Macfarland units measured using a densitometer. For our purposes, each concentration of polyphenol tested was considered a group and consisted of six hungate tubes of Lactobacilli MRS broth containing the desired polyphenol. Three tubes served as a negative control of broth containing the desired concentration of polyphenol only and three tubes which were also inoculated with LGG.
RNA extraction and RNA sequencing:
RNA was isolated from LGG grown in the presence of either 50 μg/mL of quercetin, 100 μg/mL naringenin or DMSO only. First, cultures of LGG were grown in Lactobacilli MRS broth containing either 50 μg/mL quercetin, 100 μg/mL naringenin or DMSO only, as described above. At 10 hours post inoculation the cultures were removed from the anaerobic incubator and the Macfarland units were determined using a densitometer. 15 mLs were added to a 50 mL falcon tube and spun for 10 mins at 5000 g to pellet the bacteria. After centrifugation, the supernatant was removed and 1 mL of Trizol reagent was added. These samples were then frozen and stored at -80°C until needed.
Prior to Single Molecule RNA sequencing using Helicos technology (SeqLL, Boston, MA), an RNA extraction was performed via the following Zymo Direct-zol RNA Miniprep protocol: 250 μL of Trizol was added to 100 μL of sample stored in Trizol. 350 μL of ethanol was added directly to the sample and homogenized. The sample/ethanol mixture was loaded into the Zymo-Spin II C column and centrifuged for 30 se at 16000 g. The column was washed with 400 μL of RNA wash buffer and then centrifuged for another 30 sec. 80 μL of DNase I reaction mixture (5 μL DNAse I and 75 μL of DNA digestion buffer) was added and incubated at 25°C for 15 minutes. 400 μL of Direct-zol RNA prewash was added to the column and centrifuged for 30 seconds and the flow through discarded. This step was repeated again. 700 μL of RNA wash buffer was added to the column and centrifuged for 2 minutes, repeated twice. The column was then transferred to an RNase free tube. 30 μL of RNase free water was added and centrifuged for 30 seconds. The final product was stored at -80°C until needed.
RNA Sequencing data analysis:
RNA sequencing was carried out in a Helicos sequencer by SeqLL (Boston, MA). In order to quantify the gene expression of LGG through its different treatments, we download the full assembled genomes from the National Center of Biological Information [38]. Then, we mapped the reads of each sample to its corresponding genome using UCLUST [39]. In order to increase the matching specificity, reads that were aligned to multiple locations were assigned to their corresponding best match. Thus, genes are depicted by its number of unique reads for each sample. Due to the different levels of abundance through the samples, we normalized them by its abundance and length using the RPKM [40] metric (reads per kilo base of mapped reads). Finally, the gene expression levels are computed by the log transformation of the RPMK abundances respect to each controllog 2 (RPKM [sample]/RPKM[control]). For analysis purposes, all hypothetical proteins and genes with less than 50 reads were not considered. Genes considered artifacts were removed. The fold change in expression was determined through comparison of the experimental group to the control group.
Quercetin and naringenin inhibit growth of LGG in a dose dependent manner
LGG was grown in the presence of different concentrations of either quercetin or naringenin in Lactobacilli MRS broth. Quercetin was tested at a final concentration of 12.5, 25, 50 and 75 μg/mL and naringenin was tested at a final concentration of 50, 100, 150 and 200 μg/mL. The Macfarland unit (MU) for each culture was determined using a densitometer at 0, 4, 8, 12 and 24 hours post inoculation (Figure 1). The results were plotted as a growth curve indicating the MU for each concentration at each time point (Figure 1A and Figure 1C). The percent of control was determined for each concentration at 8, 12 and 24 hours post inoculation by dividing the MU of the experimental group by the MU of the control group and multiplying by 100 (Figure 1B and Figure 1D). Time points 0 and 4 hours post inoculation are not considered for percent of control because the MU readings were too low for this type of analysis.
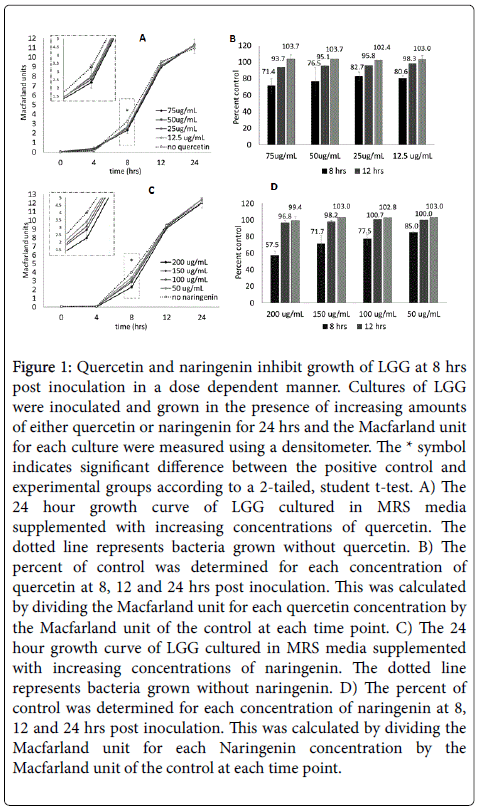
Figure 1: Quercetin and naringenin inhibit growth of LGG at 8 hrs post inoculation in a dose dependent manner. Cultures of LGG were inoculated and grown in the presence of increasing amounts of either quercetin or naringenin for 24 hrs and the Macfarland unit for each culture were measured using a densitometer. The * symbol indicates significant difference between the positive control and experimental groups according to a 2-tailed, student t-test. A) The 24 hour growth curve of LGG cultured in MRS media supplemented with increasing concentrations of quercetin. The dotted line represents bacteria grown without quercetin. B) The percent of control was determined for each concentration of quercetin at 8, 12 and 24 hrs post inoculation. This was calculated by dividing the Macfarland unit for each quercetin concentration by the Macfarland unit of the control at each time point. C) The 24 hour growth curve of LGG cultured in MRS media supplemented with increasing concentrations of naringenin. The dotted line represents bacteria grown without naringenin. D) The percent of control was determined for each concentration of naringenin at 8, 12 and 24 hrs post inoculation. This was calculated by dividing the Macfarland unit for each Naringenin concentration by the Macfarland unit of the control at each time point.
Cultures of LGG were inoculated and grown in the presence of increasing amounts of either quercetin or naringenin for 24 hrs and the Macfarland unit for each culture were measured using a densitometer. The * symbol indicates significant difference between the positive control and experimental groups according to a 2-tailed, student t-test. A) The 24 hour growth curve of LGG cultured in MRS media supplemented with increasing concentrations of quercetin. The dotted line represents bacteria grown without quercetin. B) The percent of control was determined for each concentration of quercetin at 8, 12 and 24 hrs post inoculation. This was calculated by dividing the Macfarland unit for each quercetin concentration by the Macfarland unit of the control at each time point. C) The 24 hour growth curve of LGG cultured in MRS media supplemented with increasing concentrations of naringenin. The dotted line represents bacteria grown without naringenin. D) The percent of control was determined for each concentration of naringenin at 8, 12 and 24 hrs post inoculation. This was calculated by dividing the Macfarland unit for each Naringenin concentration by the Macfarland unit of the control at each time point.
Cultures grown in different concentrations of quercetin exhibited no statistical difference from the control group of LGG grown without quercetin at hours 0 or 24 post inoculation (Figure 1A). However, there was an observed, statistically significant, suppression of growth for cultures grown in the presence of all concentrations of quercetin at both 8 and 12 hours post inoculation, with the exception of 50 μg/mL at 8 hours (Figure 1A). The most inhibition was observed at 8 hours, where LGG grown without quercetin had a MU reading of 3.27 and cultures grown with 75 μg/mL of quercetin had a MU reading of 2.5, 50 μg/mL had a MU reading of 2.5, 25 μg/mL had an MU reading of 2.7 and 12.5 μg/mL had an MU reading of 2.6 (Figure 1A). This data suggests that the inhibition of growth is dose dependent. This is demonstrated at 8 hours post inoculation where cultures grown in 75 μg/mL quercetin were only at 71.4% of control, 50 μg/mL were at 76.5% of control, 25 μg/mL were at 82.6% of control and 12.5 μg/mL were at 80.6% of control (Figure 1B). By 12 hours post inoculation the experimental groups were on their way to recovery, although still significantly inhibited (Figure 1A and Figure 1B). By 24 hour post inoculation, all groups of LGG grown with quercetin were approximately 100% of control (Figure 1B).
Similar to the results of quercetin, cultures grown in different concentrations of naringenin demonstrated no statistical difference from the control group of LGG grown without naringenin at hours 0 or 24 post inoculation (Figure 1C). There was an observed statistically significant suppression of growth for cultures grown in the presence of all concentrations of naringenin at 8 hours post inoculation (Figure 1C). At 12 hours post inoculation, the experimental groups were recovered and only 200 μg/mL of naringenin statistically inhibited growth (Figure 1C). The most inhibition was observed at 8 hours, where LGG grown without naringenin had a MU reading of 4.0 and cultures grown with 200 μg/mL of naringenin had a MU reading of 2., 150 μg/mL had a MU reading of 2.9, 100 μg/mL had an MU reading of 3.1 and 50 μg/mL had an MU reading of 3.4 (Figure 1C). The inhibition of growth for naringenin also appeared to be dose dependent. At 8 hours post inoculation, cultures grown in 200 μg/mL naringenin were only at 57.5 % of control, 150 μg/mL were only at 71.7% of control, 100 μg/mL were at 77.5 % of control and 50 μg/mL were at 85.0% of control (Figure 1D). By 24 hours post inoculation, all groups were 99% of control or greater (Figure 1D).
The effects of quercetin and naringenin on genetic expression of LGG
In order to determine what effects quercetin and naringenin have on LGG gene expression, cultures were grown in either 50 μg/mL of quercetin, 100 μg/mL of naringenin or no polyphenol for 10 hours. These doses were selected because they were the minimal amount of polyphenol required to have a moderate inhibition of growth. The 10 hour time point was selected because growth was inhibited the most at 8 hours post inoculation, but by 12 hours post inoculation the bacteria were able to recover. This indicated that in between 8 and 12 hours post inoculation, LGG is able to counteract the effects of the polyphenols. It was hypothesized that whatever mechanism LGG used to do this, would be observed in the overall RNA expression.
Cultures of LGG were grown in the presence of either 50 μg/mL of quercetin, 100 μg/mL of naringenin or no polyphenol. The Macfarland unit for each culture was determined using a densitometer at 0 and 10 hours post inoculation (Figure 2). The results were plotted as a growth curve indicating the MU for each concentration at both time points (Figure 2A). The percent of control was determined for both quercetin and naringenin by dividing the MU of the experimental group by the MU of the control group and multiplying by 100 (Figure 2B). These samples were then harvested and sent out for RNA single molecule sequencing via the Helicos Platform (SeqLL; Boston, MA).
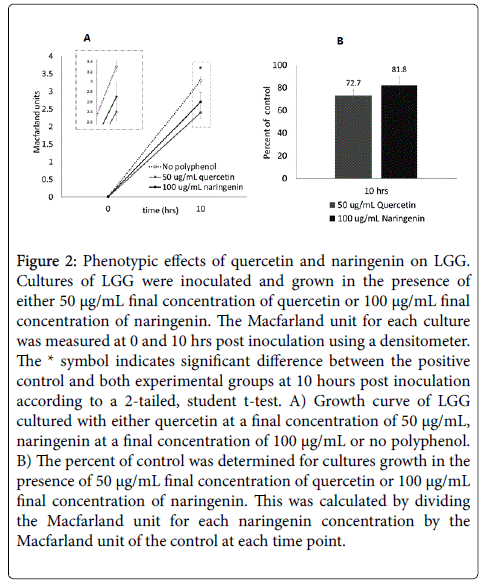
Figure 2: Phenotypic effects of quercetin and naringenin on LGG. Cultures of LGG were inoculated and grown in the presence of either 50 µg/mL final concentration of quercetin or 100 µg/mL final concentration of naringenin. The Macfarland unit for each culture was measured at 0 and 10 hrs post inoculation using a densitometer. The * symbol indicates significant difference between the positive control and both experimental groups at 10 hours post inoculation according to a 2-tailed, student t-test. A) Growth curve of LGG cultured with either quercetin at a final concentration of 50 µg/mL, naringenin at a final concentration of 100 µg/mL or no polyphenol. B) The percent of control was determined for cultures growth in the presence of 50 µg/mL final concentration of quercetin or 100 µg/mL final concentration of naringenin. This was calculated by dividing the Macfarland unit for each naringenin concentration by the Macfarland unit of the control at each time point.
Cultures of LGG were inoculated and grown in the presence of either 50 μg/mL final concentration of quercetin or 100 μg/mL final concentration of naringenin. The Macfarland unit for each culture was measured at 0 and 10 hrs post inoculation using a densitometer. The * symbol indicates significant difference between the positive control and both experimental groups at 10 hours post inoculation according to a 2-tailed, student t-test. A) Growth curve of LGG cultured with either quercetin at a final concentration of 50 μg/mL, naringenin at a final concentration of 100 μg/mL or no polyphenol. B) The percent of control was determined for cultures growth in the presence of 50 μg/mL final concentration of quercetin or 100 μg/mL final concentration of naringenin. This was calculated by dividing the Macfarland unit for each naringenin concentration by the Macfarland unit of the control at each time point.
Using systematic data processing and bioinformatic analysis, each read of RNA was matched to its corresponding gene. This provided information on which genes were being expressed and to what extent. Once the genetic expression profiles were generated, they were compared, looking for disparities in gene expression. The genetic changes between LGG grown with or without quercetin or naringenin were calculated as a fold increase or fold decrease (Tables 1-4). Each table has a column indicating the gene ID number, a column with the gene name, a column providing the fold change in expression levels and a column categorizing the known gene function. Genes that were downregulated in response to quercetin are shown in Table 1 and genes upregulated in response to quercetin can be found in Table 2. Genes that decreased in response to naringenin are provided in Table 3 and genes increased in response to naringenin are presented in Table 4.
Our results demonstrated that both quercetin and naringenin effectively suppressed growth of LGG at 8 hours post inoculation. This inhibition appeared to be dose-dependent, with the most suppression occurring at the higher doses, however, all doses were effective. Compared to quercetin, naringenin seemed to inhibit growth more radically. The high dose of naringenin produced growth equal to 57.5 % of control, while the high dose of quercetin produced growth equal to 71.43% of control. Where naringenin may have been more effective at 8 hours post inoculation, data indicated that LGG was able to recover from this inhibition more quickly compared to quercetin. At 12 hours post inoculation, LGG treated will all concentrations of quercetin still had a statistically significant inhibition of growth (Figure 1A and Figure 1B), whereas only the high dose of naringenin still significantly inhibited growth (Figure 2C and 2D).
At 24 hours post inoculation, the presence of either quercetin or naringenin was ineffectual, suggesting that LGG was able to recover from this initial growth inhibition. We hypothesized that whatever mechanism LGG used to counteract this effect, would be observed in levels of gene expression. Using the Helicos single molecule RNA sequencing platform we identified over 1100 genes expressed by LGG and compiled this information to produce genetic expression profiles. Using this information, we were able evaluate the effects of quercetin and naringenin on the genotypic level. The changes in gene numbers were calculated as fold change between groups. The threshold for evaluation was limited to a 2 fold change, and both the increase and the decrease in expression were considered, results shown in Tables 1-4.
| Gene ID | Gene name | Fold ▼ | Affected function |
|---|---|---|---|
| LGG_RS10280 | Phosphoglyceromutase | 2.7 | Metabolic function |
| LGG_RS09525 | Tat pathway signal protein | 2.7 | Molecular translocation |
| LGG_RS08335 | Ribosome silencing factor RsfS | 2.3 | Protein synthesis |
| LGG_RS13265 | Serine hydrolase | 2.3 | Stress response |
| LGG_RS08530 | Thiol reductase thioredoxin | 2.3 | Stress response |
| LGG_RS08220 | Iron-sulfur cluster biosynthesis protein | 2.3 | Protein synthesis |
| LGG_RS12110 | Glutamate synthase | 2.2 | Protein synthesis |
| LGG_RS08005 | Ribosome biogenesis GTPase RsgA | 2.1 | Protein synthesis |
| LGG_RS07925 | Signal recognition particle-docking protein FtsY | 2.0 | Molecular translocation |
| LGG_RS07490 | Peptide-methionine (R)-S-oxide reductase | 2.0 | Molecular translocation |
| LGG_RS13135 | PTS mannose transporter subunit IID | 2.0 | Metabolic function |
| LGG_RS12665 | Holin | 2.0 | Stress response |
Table 1: Genes downregulated in response to treatment of LGG with quercetin.
| Gene ID | Gene name | Fold ▲ | Affected function |
|---|---|---|---|
| LGG_RS06455 | MerR family transcriptional regulator | 2.7 | Transcriptional regulation |
| LGG_RS10850 | Recombinase RecR | 2.5 | DNA repair |
| LGG_RS11270 | Phosphoglycerate mutase | 2.5 | Metabolic function |
| LGG_RS08040 | DNA-directed RNA polymerase subunit omega | 2.5 | Transcriptional regulation |
| LGG_RS07570 | Membrane protein | 2.4 | Structural function |
| LGG_RS09870 | UDP-galactopyranose mutase | 2.4 | Protein synthesis |
| LGG_RS12855 | L-rhamnose mutarotase | 2.2 | Metabolic function |
| LGG_RS14075 | RNA-binding protein | 2.2 | Transcriptional regulation |
| LGG_RS01265 | Alkaline phosphatase | 2.2 | Cell signaling |
| LGG_RS13930 | PTS mannitol transporter subunit IIA | 2.1 | Metabolic function |
| LGG_RS11220 | dUTP diphosphatase | 2.1 | DNA repair |
| LGG_RS06890 | Phosphoribosyl-AMP cyclohydrolase | 2.1 | Protein synthesis |
| LGG_RS05165 | Branched-chain amino acid ABC transporter ATP-binding protein | 2.1 | Molecular translocation |
| LGG_RS03150 | DeoR family transcriptional regulator | 2.1 | Transcriptional regulation |
| LGG_RS07425 | ABC transporter ATP-binding protein | 2.0 | Molecular translocation |
| LGG_RS06220 | Tellurite resistance protein TelA | 2.0 | Stress response |
| LGG_RS02165 | ATP-dependent endonuclease | 2.0 | Stress response |
| LGG_RS04960 | Acetyltransferase | 2.0 | Protein synthesis |
| LGG_RS07090 | 3-phosphoglycerate dehydrogenase | 2.0 | Protein synthesis |
| LGG_RS05215 | Pyridoxamine 5-phosphate oxidase | 2.0 | Protein synthesis |
| LGG_RS05495 | XRE family transcriptional regulator | 2.0 | Transcriptional regulation |
| LGG_RS07815 | Ribosome recycling factor | 2.0 | Protein synthesis |
| LGG_RS03425 | 2-Cys peroxiredoxin | 2.0 | Stress response |
| LGG_RS12250 | Isomerase | 2.0 | Cell signaling |
| LGG_RS07970 | Phosphate acyltransferase | 2.0 | Protein synthesis |
| LGG_RS08350 | RNA-binding protein | 2.0 | Transcriptional regulation |
Table 2: Genes upregulated in response to treatment of LGG with quercetin.
| Gene ID | Gene name | Fold▼ | Affected function |
|---|---|---|---|
| LGG_RS04185 | UDP-diphosphatase | 3.9 | Peptidoglycan synthesis |
| LGG_RS08530 | Thiol reductase thioredoxin | 3.2 | Stress response |
| LGG_RS03360 | AI-2E family transporter | 3.1 | Quorum sensing |
| LGG_RS03460 | Magnesium-transporting ATPase | 2.8 | Metabolic function |
| LGG_RS02730 | Branched-chain amino acid transporter II carrier protein | 2.8 | Protein synthesis |
| LGG_RS08235 | Membrane protein | 2.8 | Structural function |
| LGG_RS10790 | Ribosomal-protein-alanine N-acetyltransferase RimI | 2.7 | Protein synthesis |
| LGG_RS01690 | PTS lactose transporter subunit IIC | 2.7 | Metabolic function |
| LGG_RS12085 | PadR family transcriptional regulator | 2.7 | Stress response |
| LGG_RS01415 | Branched-chain amino acid transporter II carrier protein | 2.5 | Protein synthesis |
| LGG_RS10720 | Amino acid permease | 2.3 | Molecular translocation |
| LGG_RS00375 | Amino acid ABC transporter permease | 2.3 | Molecular translocation |
| LGG_RS13250 | Glycerol-3-phosphate ABC transporter permease | 2.3 | Molecular translocation |
| LGG_RS00825 | Membrane protein | 2.3 | Structural function |
| LGG_RS00885 | Glyoxalase | 2.3 | Stress response |
| LGG_RS00140 | ABC transporter ATP-binding protein | 2.2 | Molecular translocation |
| LGG_RS13580 | XRE family transcriptional regulator | 2.2 | Transcriptional regulation |
| LGG_RS04565 | ABC transporter permease | 2.2 | Molecular translocation |
| LGG_RS14005 | Hydrolase | 2.2 | Metabolic function |
| LGG_RS00545 | Acetyldiaminopimelate deacetylase | 2.2 | Protein synthesis |
| LGG_RS05110 | Glucosyl transferase family 2 | 2.1 | Metabolic function |
| LGG_RS13900 | Membrane protein | 2.1 | Structural protein |
| LGG_RS02940 | ABC transporter ATP-binding protein | 2.1 | Molecular translocation |
| LGG_RS03715 | Mechanosensitive ion channel protein MscS | 2.1 | Stress response |
| LGG_RS13295 | PTS sorbose transporter subunit IIC | 2.1 | Metabolic function |
| LGG_RS02990 | Ammonia permease | 2.0 | Molecular translocation |
| LGG_RS12960 | Membrane protein | 2.0 | Structural function |
| LGG_RS03330 | PadR family transcriptional regulator | 2.0 | Stress response |
| LGG_RS00445 | Membrane protein | 2.0 | Structural function |
| LGG_RS05490 | Antibiotic biosynthesis monooxygenase | 2.0 | Metabolic function |
| LGG_RS04690 | ABC transporter permease | 2.0 | Stress response |
Table 3: Genes downregulated in response to treatment of LGG with naringenin.
| Gene ID | Gene name | Fold ▲ | Affected function |
|---|---|---|---|
| LGG_RS11035 | 1_4-dihydroxy-2-naphthoate octaprenyltransferase | 2.4 | Metabolic function |
| LGG_RS12315 | PTS galactitol transporter subunit IIA | 2.4 | Metabolic function |
| LGG_RS09440 | Transposase | 2.3 | Genome stability |
| LGG_RS09525 | Tat pathway signal protein | 2.3 | Molecular translocation |
| LGG_RS07270 | Primase | 2.2 | DNA synthesis |
| LGG_RS08810 | Recombinase RecX | 2.1 | DNA Repair |
| LGG_RS10320 | Glycine/betaine ABC transporter | 2.1 | Molecular translocation |
| LGG_RS08750 | Alpha-galactosidase | 2.1 | Energy utilization |
| LGG_RS10825 | Initiation-control protein | 2.0 | DNA synthesis |
| LGG_RS11330 | Gamma-glutamyl kinase | 2.0 | Protein synthesis |
| LGG_RS10155 | Acetyl-CoA carboxylase carboxyl transferase subunit alpha | 2.0 | Protein synthesis |
Table 4: Genes upregulated in response to treatment of LGG with naringenin.
According to our analysis, the changes in genetic expression due to the addition of either quercetin or naringenin were divergent from each other, with only a few similarities (Tables 1-4). This difference in response indicated that quercetin and naringenin suppress growth through alternate mechanisms. If quercetin and naringenin function through separate pathways, it is reasonable that LGG would respond through the upregulation and downregulation of different sets of genes. Of note, it was determined that neither quercetin nor naringenin had an effect on expression of P40 (data not shown).
Of the genes evaluated, the only one downregulated for both treated groups was the thiol reductase thioredoxin gene, with a 2.3 fold decrease for LGG treated with quercetin (Table 1) and a 3.2 fold decrease for LGG treated with naringenin (Table 3). Thiol reductase thioredoxin is involved in the cellular defense against oxidative stress by actively regenerating thioredoxin, a protein that reduces thiols after exposure to oxygen [41]. In this way, the thioredoxin system counteracts the accumulation of oxidized thiols, which can be toxic to anaerobic bacteria [42]. However, LGG is microaerophilic and can tolerate low levels of oxygen. Therefore, it is possible that this reducing system is not essential, and can be downregulated in an effort to divert energy to more important cellular functions. This may explain why there was a decrease in expression of thiol reductase thioredoxin for LGG treated with either quercetin or naringenin, even if they inhibit growth through separate mechanisms.
According to our analysis, the recombinase pathway was the only pathway displaying a greater than 2 fold increase in expression for groups treated with quercetin or naringenin compared to the control. For quercetin it was observed as a 2.5 fold increase in recombinase RecR (Table 2), and for naringenin it was demonstrated as a 2.1 fold increase in recombinase RecX (Table 4). While these genes are not identical, their products, RecR and RecX, both interact with the protein RecA [43]. RecA is responsible for repairing DNA breaks through homologous recombination, the reactivation of a stalled replication fork, and initiation of the SOS response [44].
While both RecX and RecR are accessory proteins for RecA, it has been previously determined that they have opposite functions. RecR is required for efficient homologous recombination, allowing RecA to repair DNA and initiate the SOS response [45]. Alternately, research has revealed that RecX inhibits the action of RecA in Escherichia coli, inhibiting initiation of the SOS response [46]. This information indicates that treatment with quercetin may induce DNA repair and the SOS response, while treatment with naringenin inhibits this response. This supports the proposal that quercetin and naringenin function through separate mechanisms, resulting in a different type of cellular response. It may also explain why naringenin treatment was more inhibiting at 8 hours post inoculation and why LGG was able to recover more quickly compared to quercetin treatment (Figure 1B and Figure 1D).
Interestingly, there were two genes identified that were affected inversely by quercetin and naringenin. First, the twin-arginine protein translocation (Tat) pathway signal protein was downregulated 2.7 fold in response to quercetin (Table 1), yet upregulated 2.3 fold in response to naringenin (Table 4). The Tat system is responsible for the movement of large, folded proteins into or through the cell wall [47,48]. Second, the ABC transporter ATP-binding protein was upregulated 2 fold for LGG treated with quercetin (Table 2) and downregulated 2.2 fold for LGG treated with naringenin (Table 3). The ABC pathway is involved in multi-drug resistance, due its ability to translocate molecules, or efflux molecules out of the cell [49,50]. Taken together, this data indicates that while treatment with quercetin may inhibit protein export from the cell, it enhances the efflux of smaller molecules. Conversely, treatment with naringenin promotes the extracellular transport of proteins, but inhibits movement of smaller molecules.
Lastly, it was recognized that the XRE family transcriptional regulator was increased by 2 fold for LGG treated with quercetin (Table 2) and decreased by 2.2 fold for LGG treated with naringenin (Table 3). However, the XRE family transcriptional regulator gene ID numbers for LGG treated with quercetin and LGG treated with naringenin were different. This means that while these two transcription factors belonged to the same family, they may recognize different promoter elements. Because this is the largest family of transcription factors found in Lactobacillaceae, consisting of 298 transcription factors [51], we are unable to determine which regulons were affected by these genes. It is likely that the genes regulated by these transcription factors were not related at all, and for this reason no conclusions can be drawn.
The expression profiles of LGG treated with quercetin and the control group had a total of 1,130 genes in common. Of these genes, 640 were considered upregulated in response to quercetin, with 26 genes exhibiting a greater than 2 fold increase. Conversely, 490 genes were considered down regulated in response to quercetin, with 12 genes exhibiting a greater than 2 fold decrease in expression. Because we know that LGG is able to overcome the effects of quercetin between 8 and 12 hours post inoculation (Figure 1), it would be assumed that all of the genes downregulated are either non-essential or expendable, at least for a short period of time. In general, genes that were downregulated in response to quercetin were involved in processes such as metabolic function, molecular translocation, protein synthesis and stress response (Table 1).
The two genes that decreased the most in response to quercetin, a 2.7 fold decrease, were the phosphoglyceromutase gene and the Tat pathway signal protein gene (Table 1). Phosphoglyceromutase is a necessary component of glycolysis and gluconeogeneisis [52], and the Tat pathway is involved in large protein movement into or through the cell wall [47,48]. A decrease in phosphoglyceromutase would result in a decrease in overall cellular metabolism. The observed decrease in phosphoglyceromutase may be due to direct interference by quercetin, but may also be just a side effect of stalled growth. It stands to reason that if LGG is not growing, or is growing at a slower rate compared to the control, that the metabolism would also be decreased in comparison. This means that the decrease in phosphoglyceromutase would be an indirect effect of quercetin treatment. Likewise, a decrease in the Tat pathway signal protein may be directly due to inhibition by quercetin. However, it may also be an indirect effect of slower growth.
Of the 26 genes upregulated by quercetin, 2 are involved in DNA repair and 6 play a role in transcription regulation. An upregulation of genes associated with these cellular processes supports previous findings that quercetin inhibits the function of DNA gyrase [53]. DNA gyrases, only present in prokaryotes, are responsible for coiling of bacterial DNA. Quercetin is known to bind DNA gyrase, resulting in DNA breaks [54]. Our analysis found that in response to quercetin treatment there was a 2.5 fold increase in the Recombinase RecR gene and a 2.1 fold increase in the dUTP diphosphatase, both of which are involved in DNA repair pathways (Table 2).
LGG treated with quercetin also demonstrated an increase in overall transcription regulation. Six genes were identified that are known to be involved in transcription, a MerR family transcriptional regulator, the DNA-directed RNA polymerase subunit omega, two RNA-binding proteins, a DeoR family transcriptional regulator and an XRE family transcriptional regulator (Table 2). Interestingly, the MerR family transcriptional regulator gene had the highest increase in expression, with a 2.7 fold increase. The MerR family is traditionally identified as responding to mercury and works to detoxify the cells of heavy metals. However, this family is indicated in responding to not only mercury and heavy metals, but also to reactive oxygen species [55,56]. The DeoR family of transcriptional regulators play a role in sugar metabolism and usually function as a repressor [57]. However, because these families of transcription regulators are large it is unclear which genes they are actually regulating. However, a general upregulation in transcription would make sense if the bacteria is undergoing DNA repair in response to quercetin treatment.
The expression profiles of LGG treated with naringenin and the control group had a total of 1,139 genes in common. Of these genes, 677 were considered upregulated in response to quercetin, with 11 genes exhibiting a greater than 2 fold increase. 462 genes were considered downregulated in response to quercetin, with 31 genes exhibiting a greater than 2 fold decrease in expression. Similar to quercetin, we know that LGG o overcome the effects of naringenin, so again, it would be assumed that genes with a decrease in expression are not important or dispensable, at least for some time. It is interesting that, unlike LGG treated with quercetin, for LGG treated with naringenin there are approximately 3 times more genes downregulated compared to upregulated. Genes that were downregulated in response to naringenin included 6 that were involved in stress response, 6 involved in metabolic function, 4 involved in protein synthesis, 7 involved in molecular translocation, 5 involved in structural function, 1 involved in quorum sending and 1 involved in peptidoglycan synthesis (Table 3).
The gene with the most change in response to naringenin was the UDP-diphosphatase, with a 3.9 fold decrease in expression (Table 3). UDP- diphosphatase plays a role in peptidoglycan and techoic acid synthesis [58]. It is possible that naringenin inhibits cell wall synthesis by blocking the action of UDP-diphosphate, initially inhibiting growth. However, it is also possible that the decrease in cell wall synthesis is due to overall slower growth compared to the control group. In this case, naringenin may be directly or indirectly effecting expression of this gene.
Interestingly, there is 3.1 fold decrease in the AI-2 or, Auto- Inducer-2 gene. This gene is involved in quorum sensing for both gram positive and gram negative bacterial cells [59]. Previous reports have demonstrated that naringenin effects the quorum sensing ability of Psuedomonas aeruginosa by inhibiting production of the acylhomoserine lactones [60]. Taken together, this suggests that naringenin can adversely affect LGG’s quorum sensing capability by inhibiting expression of the A1-2E family transporter. Perturbations in quorum sensing can interrupt vital, cell-cell communications on population density and collective gene expression [61]. A decrease in genetic expression of the A1-2E family transporter may affect the growth of LGG. This may explain why growth was inhibited more severely for the naringenin treated groups compared to the quercetin treated groups at T=8 hrs (Figure 1).
There is an overall decrease in stress response genes for LGG treated with naringenin. This is observed as a decrease in Thiol-reductase thioredoxin, two PadR family transcriptional regulators, glyoxalase, mechanosensitive ion channel protein MscS and an ABC transporter permease. Of interest is the decrease in two of the PadR family transcriptional regulators. The PadR transcription system is known to function as a response to stress caused by phenolic acids in gram positive bacteria [62,63]. A decrease in this expression indicates that naringenin is not initiating a phenolic acid response. Taken together, it seems that while naringenin is able to inhibit growth of LGG, it is not initiating a stress response. So the cells are growing slower, but not stressed, which is supported by the fact that naringenin inhibits initiation of the SOS response.
There were only 11 genes found to be upregulated by LGG treated with naringenin. They consist of genes involved in pathways such as metabolic function, genome stability, molecular translocation, DNA synthesis and repair, protein synthesis and energy utilization. There were two genes with a 2.4 fold increase in expression, both involved in metabolic function. The first gene is 1_4-dihydroxy-2-napthoate octaprenyltransferase, which is involved in vitamin biosynthesis [64]. The second gene is the PTS galactitol transporter subunit IIA, which is involved in sugar uptake [65]. Taken together, this data indicates that treatment of LGG with naringenin does not initiate a stress response, but does lead to an increase in metabolism. These findings further support the idea that quercetin and naringenin inhibit growth through separate mechanisms and that LGG responds through alternate pathways.
LGG is a commonly used probiotic that has a number of healthy qualities. We have been able to characterize the interactions between LGG and the polyphenols quercetin and naringenin through phenotypic and genotypic analysis. Interestingly we find that LGG is able to overcome the initial inhibitory effects of these polyphenol through up regulation and down reguation of multiple genes. This is also a rare demonstration of how single molecule sequencing technology can be used for microbiota expression analysis. The results of our genetic analysis are remarkably consistent with the observed LGG growth. The omission of any amplification step in RNA sequencing likely improve the accuracy of our analysis.