Journal of Pollution Effects & Control
Open Access
ISSN: 2375-4397
ISSN: 2375-4397
Research Article - (2016) Volume 4, Issue 2
Titanium dioxide nanoparticles (TiO2-NPs) have recently been of public health and scientific concern due to their widespread use in industrial and household applications. However, there is limited information concerning its in vivo cytogenotoxicity. In this study, the cytogenotoxic effects of TiO2-NPs on the somatic tissue using the mouse bone marrow micronucleus (MN) assay and on reproductive tissue using the mouse sperm morphology assay and testicular histopathology were investigated. Five concentrations of 9.38, 18.75, 37.50, 75.00 and 150.00 mg/kg bwt were administered intraperitoneally at 0.5 mL/mouse to mice for five and ten consecutive days in the MN assay; and for five consecutive days in the sperm morphology assay. Double distilled water and cyclophosphamide (20 mg/kg bwt) served as negative and positive controls, respectively. A significant (p<0.05) increase in MN was observed in bone marrow cells of treated mice at 37.50 mg/kg bwt concentration in the 5-day exposure and at all concentrations in the 10-day exposure. The sperm cells examined 5 and 10 weeks from the first day of exposure showed significant increase (p<0.05) in abnormal sperm cells at tested concentrations. Histopathologically, TiO2-NPs disrupted the normal cellular architecture of testicular tissues in exposed mice; as it caused severe lesions such as congestion of the interstitium oedema, vacuolation and necrosis. These suggest that the bone marrow and testicular cells may be potential targets for TiO2-NPs induced DNA damage and cytotoxicity in mice. This is of public health importance considering increasing exposure to TiO2-NPs in consumer products.
Keywords: Titanium dioxide nanoparticles; DNA damage; Micronucleus; Histopathology; Mouse sperm morphology
Nanotechnology as an emerging science in this millennium has led to the advancement in the production of nanoparticles [1]. It creates opportunities for engineers to manufacture superior and more durable devices and products [2], and boost scientific interest to ascertain their impact on the biotic and abiotic components of the ecosystem. The development and enlarging research interest in nanoparticles and nanomedicine have led to a huge potential for novel ways of rapid disease diagnosis, treatment and enhanced quality of life [2].
Nanoparticles (NPs) exist as naturally occurring nanoparticles (e.g. volcanic ash, ocean spray and storm dust) and engineered NPs (ENPs). ENPs include carbon based (e.g. fullerenes, carbon nanotubes), inorganic NPs such as [metal- e.g. silver, iron, copper, manganese; and metal oxides e.g. - titanium dioxide, zinc oxide, copper oxide, silicon oxide)] and quantum dots (e.g. cadmium and selenium) [3]. They have a small size and large surface area to volume ratio with high reactivity potential, as a result of these unique properties, they have being massively produced by industries that use them on a large scale [4,5]. The increased production of these particles enhances the probability of exposure through inhalation, oral and dermal penetration, both in the occupational and environmental settings [6-8].
Among the available metal oxide NPs used in the manufacturing of consumer products are titanium dioxide nanoparticles (TiO2-NPs), which are the earliest industrially produced nanomaterial’s [9] and one of the most highly manufactured in the world [10]. There are three different crystalline structures of TiO2-NPs: anatase, rutile and brookite [11]. Anatase is more chemically reactive and capable of generating reactive oxygen species [12] while rutile is the most natural form of TiO2 and is said to be chemically inert [11,12]. TiO2- NPs account for over 70% of the total production volume of nanoparticles worldwide. It is a white pigment and mostly used because of its brightness and very high refractive index. It is used in diverse areas of application such as ointments, toothpaste, plastics, rubber, printing inks, floor coverings, automotive products, food colorants, catalysts, adsorbents, semi-conductors, mortar, ceramics, whitening and brightening of food, especially for confectionary and certain powdered foods. They are also used in the pharmaceutical industry as an opacity agent, and in environmental decontamination of air, soil, and water [13-15]. In some of these products, the amount of TiO2-NPs is more than 10% by weight [16,17]. In spite of the increased application of TiO2-NPs especially in consumer products, little is known about the potential toxicities and the underlying mechanisms, and this has generated major concerns among scientists especially toxicologists on their potential genotoxic and cytotoxic effects [18-20].
Data regarding the genotoxicity studies of TiO2-NPs are inconsistent, as their toxicities are complex and depend on the physicochemical properties such as size, surface area, crystalline structure, surface properties, agglomeration and solubility [4,5,21,22]. There are studies on in vitro genotoxic effects of TiO2-NPs in human lymphocytes [23,24], human hepatoma HepG2 cells [25,26], Chinese Hamster Ovary Cells (CHO) [27,28], human bronchial epithelial cell line (BEAS 2B) [29], human epidermal cell line (A431) [30], human lung cancer cells (A549) [31], and human SHSY5Y neuronal cells [32]. Whether photo activated or not, TiO2-NPs (anatase or rutile) have been found to induce DNA damages [33-36], micronuclei formation [26,28,37], cell necrosis and apoptosis through the formation of reactive oxygen species [21,38- 40], impair cell function in human dermal fibroblasts and decrease cell area, cell proliferation, and cell mobility [41]. In vivo, TiO2-NPs have been reported to cause inflammatory reaction [42,43], oxidative DNA damage [44-47], pulmonary fibrosis [48] and serious damage to the liver, kidneys and myocardium in mice [49]. Information on in vivo genotoxicity of titanium dioxide nanoparticles is limited. This may be of importance for bone marrow and germ cells, where adverse impacts may affect the potentials for self-renewal and differentiation. With the enormous applications of TiO2-NPs in consumer products, they can be absorbed via inhalation, ingestion, and dermal penetration into the body systemic circulation and reached important viscera organs. Hence, in this study, we investigated the genotoxic effect of TiO2 NPs in mice using induction of micronucleus and abnormal sperm morphology as the genetic end points. Additionally, the toxic effect on the histology of the testes of exposed mice was also examined.
Test substance and preparation of TiO2-NPs stock solution
Titanium dioxide nanopowder [(TiO2-NPs, anatase, CAS number: 1317-70-0, product code - 637254), Purity: 99.7%, Average Particle Size: <25 nm, Specific Surface Area: 45 m2/g, Color: white, Morphology: powder, Bulk density: 0.04-0.06 g/mL and relative density: 3.9 g/mL] was obtained commercially from Sigma Aldrich Co. Germany. This NP was chosen because of its utilization in previous studies [30], and the physico-chemical characterization is as reported by Shukla et al. [30]. The TiO2-NPs were suspended in double distilled water at a stock concentration of 150 mg/kg body weight (adapted from IP LD50 in mice) [49] and ultrasonicated (BANDELIN Sonorex digitec Germany - DT 52H, 230 V – 50/60 Hz; 0.9 A; 60/240 W; 35 kHz) for 1h (3 min pulse on and 30 sec pulse off) at 60W. Freshly prepared working solutions were made by serial dilution from the stock solution; after vortex for five minutes before each dilution process to disperse the particles. Five concentrations of 9.38, 18.75, 37.50, 75.00 and 150.00 mg/Kg bwt of TiO2-NPs were utilized in this study.
Biological materials
Young male Swiss albino mice (Mus musculus, 5-6 and 8-12 weeks old) were obtained from the National Institute for Medical Research (NIMR) Lagos, Nigeria. They were acclimatized for two weeks in the animal house of the Department of Zoology, University of Ibadan, Nigeria with food (Ladokun® pelleted feed) and drinking water (tap water) supplied ad libitum. Mice of 7-8 weeks of age were used for the micronucleus assay; while those of 12-14 weeks were used for the sperm morphology assay. Ethical approval was obtained from the Animal Care and Use in Research Ethics Committee (ACUREC) of the University of Ibadan, Ibadan, Nigeria (UI-ACUREC/App/2015/005).
Micronucleus (MN) assay
Two exposure durations of 5- and 10- days were considered. There were five concentrations of 9.38, 18.75, 37.50, 75.00 and 150.00 mg/kg bwt (corresponding to × 0.0625, × 0.125, × 0.25, × 0.5 and × 1 of IP LD50 of TiO2 NPs) per exposure period, and four mice (weight range of 22– 30 g) per concentration. Double distilled water and cyclophosphamide (20 mg/kg; Cadila Healthcare Limited, Ponda, Goa) served as negative and positive controls respectively. Each mouse per concentration was intraperitoneally (IP) administered 0.5 mL of the respective dosage per day for 5 or 10 consecutive days depending on the exposure period. Bone marrow cells preparation for MN assessment was according to Schmid [50] as modified by Bakare et al. [51]. Briefly, the animals were sacrificed by cervical dislocation. The femurs were removed and bone marrow flushed with Fetal Bovine Serum (FBS) (PAA Laboratories GmbH, PAA-Strasse 1, Pasching. Austria). Cells were centrifuged at 2000 rpm for 5 min and slides were stained with May-Grunwald and Giemsa stains. At least 2000 erythrocytes per mouse were scored at ×1000 for MN in polychromatic erythrocytes (MNPCE) and normochromatic erythrocytes (MNNCE).
Sperm morphology assay
Induction of sperm abnormalities was done in accordance with standard procedures [52,53]. Mice of 12-14 weeks of age (weight range of 20-36 g) were utilised. They were divided into two groups based on the exposure periods of 5 and 10 weeks. In each exposure period there were 5 dosage groups (6 mice per dosage group) just as in the MN assay. A single intraperitoneal (IP) injection of 0.5 mL of the respective dosage group was administered to the mice daily for 5 consecutive days. The same volume but of cyclophophasmide (20 mg/kg) and double distilled water was administered to mice in the positive and negative control groups respectively. Sperm cells were sampled from the cauda epididymes at 5 and 10 weeks from the first exposure. Four mice per dosage group were sacrificed by cervical dislocation and their cauda epididymes removed; sperm suspensions were then prepared from the cauda of each testis by mincing the cauda in normal saline and 1% eosin Y stain. The slides were air-dried and coded for subsequent microscopic examination at x1000. For each mouse, 1000 sperm cells were assessed for morphological abnormalities according to the criteria of Wyrobek and Bruce [54].
Histopathological analysis of the testes
Testes were carefully excised from two mice randomly selected from each dosage group exposed to 9.38, 37.50 and 150.00 mg/Kg bwt of TiO2-NPs, and the negative and positive control groups at the end of 5 and 10 weeks exposure periods respectively. Blood and body fluids were rinsed from the testes using normal saline and were fixed in Bouin’s fluid for 48 hours. They were then dehydrated in ascending grades of ethanol (70%, 80%, 95%, and 100%), cleared using xylene and embedded in paraffin wax using Leica Histokinete tissue processor for 6 hours. Serial sections of 4 μm thickness were obtained on labeled glass slides using a rotatory microtome. The deparaffinized sections were stained routinely with haematoxylin and eosin (H & E) and mounted. The slides were scored randomly at 400X and photomicrographs taken accordingly.
Statistical analysis
The Statistical Package for the Social Sciences (SPSS®) version 16.0 and Microsoft Excel® 2007 were used for data analysis. Data obtained were expressed as percentage frequency and mean ± standard error. Significance at different concentration-level was tested using oneway ANOVA test and Duncan’s New Multiple Range Test (DMRT). Correlation analysis was done to establish relationship between the frequency of induced MN and exposure period. Difference between the negative control-group and individual concentration-groups were analyzed at the 0.05 and 0.01 probability level.
Micronucleus assay
Table 1 shows the frequency of MNPCE (Figure 1) observed in the bone marrow of mice exposed to the different concentrations of TiO2-NPs for five and ten days. TiO2-NPs increased the frequency of micronucleus in mice at all the tested concentrations compared to the negative control both in the 5 and 10 days exposure periods. At the respective tested concentrations, MNPCE increased with a 3.2, 1.5, 4.8, 3.2, and 4.1 folds at the 5 days exposure period; and 6.5, 4.5, 14.3, 15.8 and 22.0 folds at the 10 days exposure period. Significant difference (p<0.05) was however observed at 37.50 mg/kg for the 5 days exposure and at all the tested concentrations (p<0.01) except 18.75 mg/kg for the 10 days exposure. A concentration dependent increase in the frequency of MNPCE was observed in the 10 days exposure. There was a positive correlation between the frequency of MNPCE and days of exposure (r = 0.693).
| Concentration (mg/kg) | 5-days Mean ± SE | 10-days Mean ± SE |
| DDW | 10.25 ± 2.17 | 9.50 ± 2.66 |
| 9.38 | 33.25 ± 4.21 | 6 61.75 ± 8.42** |
| 18.75 | 15.25 ± 4.71 | 42.75 ± 9.69 |
| 37.5 | 49.00 ± 7.49* | 13 135.75 ± 12.04** |
| 75 | 33.00 ± 9.53 | 149.75 ± 4.37** |
| 150 | 41.50 ± 9.43 | 208.75 ± 13.33** |
| Cyp | 36.50 ± 29.73 | 24.75 ± 1.97 |
Table 1: Frequencies (Mean ± SE) of micronucleated polychromatic erythrocytes in bone marrow of mice exposed to Titanium dioxide nanoparticles.
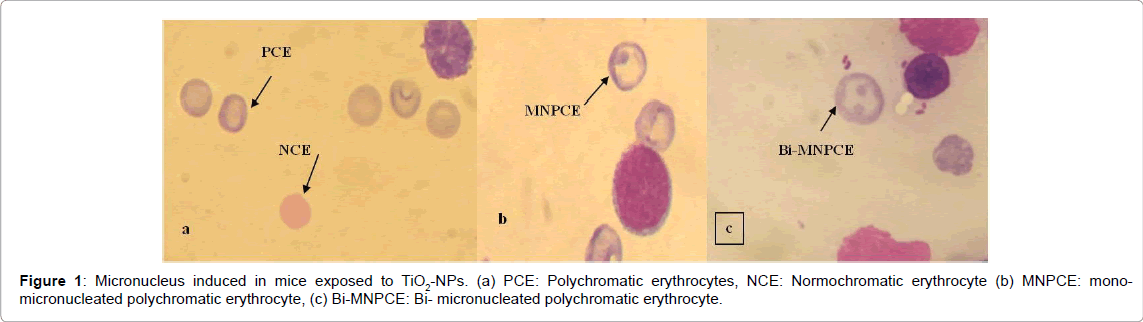
Figure 1: Micronucleus induced in mice exposed to TiO2-NPs. (a) PCE: Polychromatic erythrocytes, NCE: Normochromatic erythrocyte (b) MNPCE: monomicronucleated polychromatic erythrocyte, (c) Bi-MNPCE: Bi- micronucleated polychromatic erythrocyte.
Sperm morphology assay
The effects of the different concentrations of TiO2-NPs on the sperm morphology at the end of 5 and 10 weeks exposure periods are presented in Table 2. After 5 weeks of exposure, the percentage abnormal sperm cells were 16.44, 13.94, 15.22, 21.64 and 21.72% for 9.38, 18.75, 37.50, 75.00 and 150.00 mg/kg of TiO2-NPs respectively, which were statistically significant (p<0.05) at all the tested concentrations compared to the negative control (8.40%). At the same concentrations for the 10 weeks exposure period, there were 11.65, 8.73, 15.98, 13.95 and 15.05% abnormal sperm cells which were significant (p<0.05) at all concentrations (except at 18.75 mg/kg) compared to the negative control value of 8.20%. Generally, the mean of abnormal sperm cells induced in mice at each concentration after 5 weeks of exposure were greater than after 10 weeks of exposure. Abnormal sperm cells such as double hook, double heads, knobbed hook, double tails, pin head, banana shaped, amorphous head, folded and wrong tail attachment were observed in the mice exposed to TiO2-NPs at the two exposure periods (Figure 2). Folded sperm cells were the most predominant while sperm cells with double hooks were the least.
| Concentrations (mg/kg) | % abnormalities | Mean ± S.E. | ||
| 5 weeks | 10 weeks | 5 weeks | 10 weeks | |
| Distilled water | 8.4 | 8.2 | 84.00 ± 6.44 | 82.0 ± 8.76 |
| 9.375 | 16.44* | 11.65* | 164.40 ± 21.01* | 116.5 ± 12.16* |
| 18.75 | 13.94* | 8.73 | 139.40 ± 11.18* | 87.3 ± 12.56* |
| 37.5 | 15.22* | 15.98* | 152.20 ± 20.61* | 159.8 ± 43.17* |
| 75 | 21.64* | 13.95* | 216.40 ± 43.36* | 139.5 ± 5.07* |
| 150 | 21.72* | 15.05* | 217.20 ± 30.07* | 150.5 ± 11.07* |
| Cyp | 11.66* | 16.08* | 116.60 ± 20.49* | 160.8 ± 9.28* |
Table 2: The frequency (%) and mean (± S.E) of morphologically abnormal sperm cells induced in mice exposed to different concentrations of TiO2-NPs for 5 and 10 weeks.
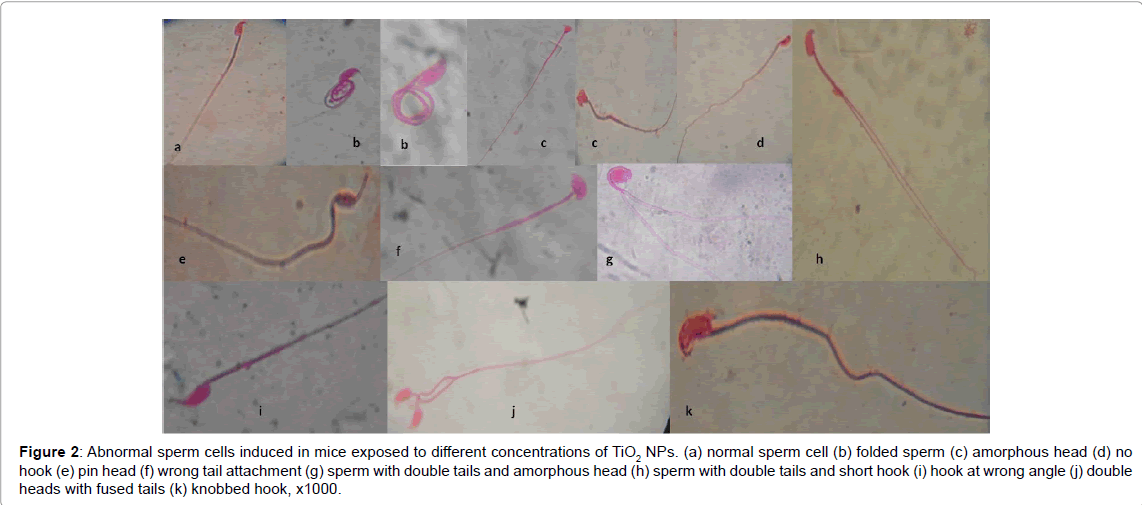
Figure 2: Micronucleus induced in mice exposed to TiO2-NPs. (a) PCE: Polychromatic erythrocytes, NCE: Normochromatic erythrocyte (b) MNPCE: monomicronucleated polychromatic erythrocyte, (c) Bi-MNPCE: Bi- micronucleated polychromatic erythrocyte.
Histopathological assessment of the testes
Microscopic examination of the testis in the negative control group showed a normal cellular architecture of the testicular tissues while the treated groups at the two exposure periods showed the disruption of the normal cellular architecture of the testicular tissues by TiO2-NPs (Figure 3). There were severe histopathological lesions such as congestion of the interstitium oedema, congestion of the interstitial blood vessels, reduced height of germinal epithelium, numerous spermatocytes and elongate spermatids (Figure 3). The lesions were most severe in mice exposed to 37.50 mg/kg of TiO2-NPs for 10 weeks as there were numerous variably-sized seminiferous tubules (ST) many of which were cystic and had irregular outlines, while the affected seminiferous tubules were severely depleted of spermatogenic cells. There were also marked necrosis and vacuolation of spermatogenic cells within the STs at this concentration.
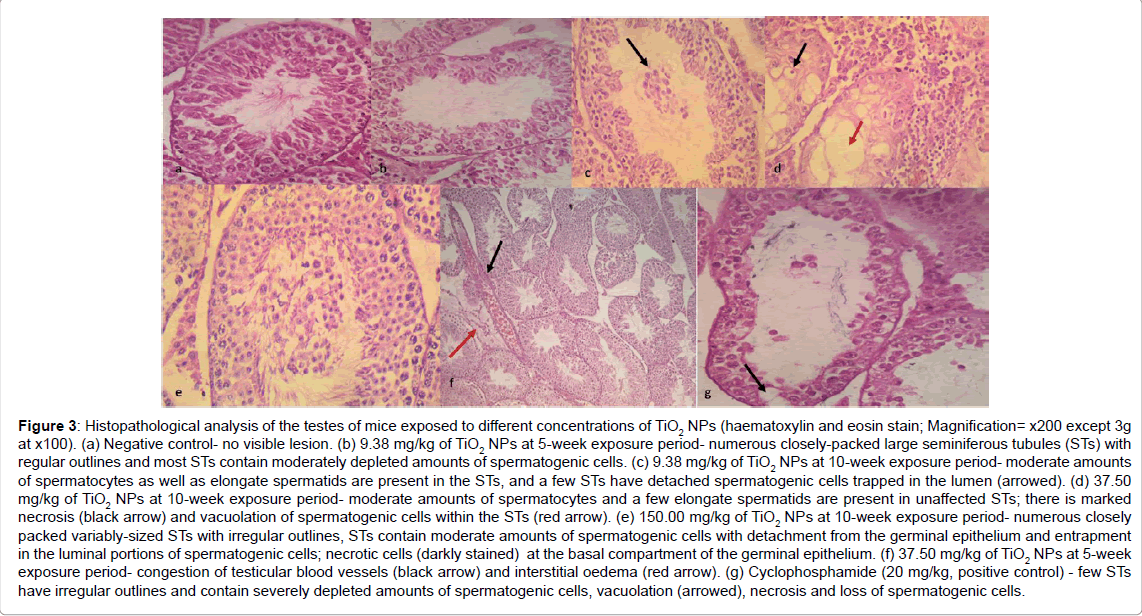
Figure 3: Histopathological analysis of the testes of mice exposed to different concentrations of TiO2 NPs (haematoxylin and eosin stain; Magnification= x200 except 3g at x100). (a) Negative control- no visible lesion. (b) 9.38 mg/kg of TiO2 NPs at 5-week exposure period- numerous closely-packed large seminiferous tubules (STs) with regular outlines and most STs contain moderately depleted amounts of spermatogenic cells. (c) 9.38 mg/kg of TiO2 NPs at 10-week exposure period- moderate amounts of spermatocytes as well as elongate spermatids are present in the STs, and a few STs have detached spermatogenic cells trapped in the lumen (arrowed). (d) 37.50 mg/kg of TiO2 NPs at 10-week exposure period- moderate amounts of spermatocytes and a few elongate spermatids are present in unaffected STs; there is marked necrosis (black arrow) and vacuolation of spermatogenic cells within the STs (red arrow). (e) 150.00 mg/kg of TiO2 NPs at 10-week exposure period- numerous closely packed variably-sized STs with irregular outlines, STs contain moderate amounts of spermatogenic cells with detachment from the germinal epithelium and entrapment in the luminal portions of spermatogenic cells; necrotic cells (darkly stained) at the basal compartment of the germinal epithelium. (f) 37.50 mg/kg of TiO2 NPs at 5-week exposure period- congestion of testicular blood vessels (black arrow) and interstitial oedema (red arrow). (g) Cyclophosphamide (20 mg/kg, positive control) - few STs have irregular outlines and contain severely depleted amounts of spermatogenic cells, vacuolation (arrowed), necrosis and loss of spermatogenic cells.
Nanotechnology has effectively improved a number of consumer products, through the manufacturing and use of Nanoparticles. Therefore, assessment of the toxicological effects of Nanoparticles on the human health and environment is inevitable as these nanoparticles through their small size, large surface area to volume ratio, and other physicochemical properties are able to disrupt the biochemical and physiological functions of the cell. In the present study, the potential genotoxic effect of TiO2-NPs was evaluated in the somatic and germ tissues of mice using the mouse bone marrow micronucleus and sperm morphology assays. The data obtained in this study showed TiO2-NPs to be genotoxic in mice.
The results of the MN assay showed that TiO2-NPs are clastogenic and aneugenic. It caused chromosomal damage in dividing cells of exposed mice. The frequency of MN induction was directly proportional to the exposure periods; the longer the number of days of exposure; the higher the chromosomal damage induced by TiO2-NPs. Mice exposed for 10 days had higher genomic damages in the form of micronuclei formation than those exposed for 5 days. This suggests that TiO2-NPs might have accumulated in a dose-dependent manner, thus affecting the bone marrow PCE for a longer time [55]; it has also been reported that the retention halftime of TiO2-NPs in vivo is long because of its difficult excretion [56]. TiO2-NPs may have interacted (directly or indirectly) with the genetic material of the bone marrow cells producing primary and/or secondary genotoxicity resulting in accentric chromosome fragments or chromosome loss. Several factors may account for the calstogenic and aneugenic characteristics of TiO2- NPs. A direct interaction between TiO2-NPs and the genetic material is a possibility. Previous studies have shown TiO2-NPs to have access to the cell membrane, without using a specific transporter or penetrate through the nuclear pore complex. They are able to produce titanium ion in the cell cytoplasm, having the potential of generating intracellular reactive oxygen species, of which the stable and diffusible forms such as hydrogen peroxide or lipid peroxidation intermediates could affect the nuclear DNA [57]. Another possible reason for the aneugenic effect of TiO2-NPs may be explained by the physical interaction with the components of the mitotic spindle during cell division or the interaction with proteins directly or indirectly involved in chromosome segregation [58]. They may physically interact with the mitochondrial membranes causing loss of the mitochondrial membrane potential, the opening of the permeability transition pores and ROS production [5,55].
Our result is in accordance with those of previous studies wherein mice [33,44,59], human peripheral blood lymphocytes [60,61] and cell lines [62] were used as test systems. However, it differs from those of Lindberg et al. [63], Sadiq et al. [64], Xu et al. [65] and Kim et al. [66] wherein TiO2-NPs were reported to be non-clastogenic/aneugenic. This could be because of differences in exposure duration, degree of agglomeration, particle size, and chemical composition of the NPs.
The result of the sperm morphology assay showed the spermatotoxic effect of TiO2-NPs. The mouse sperm morphology assay has potential in identifying chemicals that induce spermatogenic dysfunction and perhaps heritable mutations [52]. Sperm abnormalities have long been associated with male infertility and sterility in most species and the structure play a substantial role in both fertilization and pregnancy outcome [67]. Two sperm cells collection periods were considered: 5 and 10 weeks. The 5-week assessed sperm cells were exposed as differentiating and mitotically dividing spermatogonia, while the 10- week assessed sperm cells were exposed as mitotically dividing stem cells. Higher rate of sperm abnormalities was recorded for the 5-week assessed cells compared to the 10-week assessed cells. This could be an indication that differentiating and mitotically dividing spermatogonia are more susceptible to TiO2-NPs damage than mitotically dividing stem cells.
The exact mechanism for the increase in the frequency of abnormal sperm is not clear and opinions on this subject differ. The induction of abnormal sperms is assumed to be as a result of an abnormal chromosome [68], minor alteration in testicular DNA [69], and point mutation [70]. According to several studies [71-74], small deletions, point mutations, and abnormal chromosomes are proposed as possible genetic causes of such alterations. Bruce and Heddle [75] attributed the occurrence of sperm head abnormalities to the chromosomal aberrations that occur during the packaging of genetic material in the sperm head or occurrence of point mutation in testicular DNA [75]. Sperm abnormalities may also arise as a consequence of mistakes in the spermatozoa-differentiating process during spermatogenesis or by physiological, cytotoxic or genetic mechanisms or alterations in testicular DNA which in turn disrupts the process of differentiation of spermatozoa [52].
This report supports the chromosomal damage as possible genetic cause of such alterations, since the TiO2-NPs were found to be clastogenic in the MN assay. The reproductive toxicity of TiO2-NPs herein is in accordance with previous reports of other types of NPs. Yoshida et al. [76] showed adverse effects of carbon NPs on the male reproductive systems of adult mice. Also, Gromadzka-Ostrowska et al. [77] observed a decrease in sperm count, increased DNA damage and a change in testis seminiferous tubule morphometry of male rats exposed to Ag NPs. Au NPs was also reported to cause a drop in sperm motility and increase sperm fragmentation [78].
The result of the histopathology of the exposed mice testes showed that TiO2-NPs caused pathological changes to the testes. Administration of TiO2-NPs caused severe histopathologic lesions such as congestion of the interstitium oedema, moderate/mild congestion of the interstitial blood vessels, reduced height of germinal epithelium, fairly numerous spermatocytes and few elongated spermatids. The degenerative changes observed in the seminiferous tubules such as necrosis, vacuolation and congestion of the interstitial/testicular blood vessels are evidence of the toxicity of TiO2-NPs to the mouse male reproductive system. Degenerative changes in the seminiferous tubules indicate that TiO2- NPs may directly interfere in the process of spermatogenesis [79]. This was probably a pre-requisite for the observed abnormal sperm morphology produced by the damaged testes. This result is consistent with previous reports on histopathological damage or lesions in the male testes of mouse/rat by TiO2-NPs [80,81] but in contrast to the report of Guo et al. [82] who reported no obvious pathological changes in the testis of male mice exposed to TiO2-NPs. That TiO2-NPs induced abnormalities in mouse sperm cells and the testes suggests that the same could happen in other exposed male animal species especially mammals. DNA-damaged spermatozoa may introduce damaged genome into the oocytes with dangerous drawbacks on fertilization, embryonic, foetal and post-natal development [83,84]. Although the clinical significance of sperm morphology is still a matter of debate, it has been recently recognized that an accurate definition of morphological anomalies plays a very important role in the determination of male fertility potential [85].
One of the possible mechanisms for TiO2-NPs induced genotoxicity herein is oxidative stress [86] as TiO2-NPs are photocatalytic and have been implicated to directly generate free radicals [87,88]. NPs are able to generate reactive oxygen species caused by secondary mechanical processes associated with inflammatory responses ultimately causing cell damage and eventually cell death [33,44]. Likewise, the small particle size and large surface area of TiO2-NPs enables them to easily penetrate cells and cellular components thus interfering with several sub-cellular mechanisms and biomolecules causing lipid peroxidation, mitochondria disruption, immune reactivity and protein damage [5].
Our study has shown that TiO2-NPs have the capacity to interact with mice genetic materials / machinery under the test condition. This is of public health importance considering industrial and household applications of TiO2-NPs. Chemically induced genetic damage has been implicated in the etiology of many diseases; thus, there is need for stringent policies as regards the use of nanoparticles in human consumable and cosmetic products as well as their disposal into the environment.
We appreciate the support of Department of Zoology, University of Ibadan vote for postgraduate studies; and the technical assistance of Naomi Adeyemo.
Bakare, Alimba and Alabi conceived and designed the experiments. Udoakang, Anifowose, Fadoju, Ogunsuyi and Oyeyemi performed the experiments and analyzed the data. Udoakang and Anifowose drafted the manuscript; while Bakare, Alimba and Alabi critically revised it, gave final approval and agrees to be accountable for all aspects of work ensuring integrity and accuracy.