Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2017) Volume 8, Issue 3
The study assessed the protein sparing effect resulting from carbohydrate (CHO) and lipid (LIP) use and metabolic responses of juvenile tambaqui fed diets with different proportions of CHO and LIP. In a completely randomized 3 × 2 factorial design, the 6 diets tested combined 3 CHO (410 gkg-1, 460 gkg-1 and 510 gkg-1) and 2 LIP (40 gkg-1 and 80 gkg-1) levels. The 1080 fish tested (10.88 g ± 0.13 g mean body weight) were randomly distributed into twenty-four 500-L tanks (45 fish per tank). An increase in diet CHO and LIP levels improved feed conversion and protein efficiency ratio. The highest addition of CHO (510 gkg-1) and LIP (80 gkg-1) reduced diet intake and weight gain and increased serum triglyceride levels and fat deposition in muscle and liver. Glucose levels, serum protein and liver glycogen showed no difference between treatments with different CHO and LIP levels. Liver and muscle fat deposition were higher in the treatments with the highest diet LIP level, irrespective of diet CHO. The hepatosomatic index (HSI) and viscerosomatic fat index (VSI) exhibited no differences between treatments. Therefore, tambaqui can use both CHO and LIP to optimize protein use. The physiological responses indicate that the best CHO-LIP combination in tambaqui diets is 460 gkg-1 and 40 gkg-1, respectively. This combination promotes satisfactory growth without causing excess fat accumulation.
Keywords: Body composition; Non-protein energy; Nutrition; Plasma metabolites; Protein sparing effect
The energy required to feed different physiological processes in fish is produced from the catabolism of macronutrients, such as proteins, carbohydrates (CHO) and lipids (LIP) [1]. Protein, however, is the most expensive ingredient in the diet, and therefore, studies on nutrition aim to determine the minimum levels required to obtain the best fish performance, thereby reducing diet production costs in addition to minimizing ammonia excretion, which compromises water quality [2]. The inclusion of non-protein energy sources reduces the use of diet protein for energy production, a process called the protein sparing effect [1]. With low cost and high availability, CHO and LIP are the main non-protein energy sources in fish diet. CHO content in tropical fish diets is quite high, and its addition can improve pellet quality and optimize the use of other nutrients [3].
Diets with high CHO content are more efficient in omnivorous and/or herbivorous fish than carnivorous fish [4]. Tambaqui (Colossoma macropomum ) is an omnivorous Amazonian fish, with a digestive tract characteristic of diversified food items, dentition adapted to eating fruits and seeds and an efficient filtrating apparatus for capturing zooplankton [5]. Despite the efforts to develop technological solutions to farm this species, information on its nutrition and metabolism remain inconclusive.
According to Li et al. [6], a diet with an optimal proportion of CHO and LIP optimizes the use of these nutrients, whereas an imbalance of non-protein energy sources or inadequate amounts of nutrients can directly affect liver and body composition. Accordingly, LIP profile must be assessed in the liver, an essential metabolic organ, and white muscle because it is the commercially exploitable portion of fish. In addition, analysis of blood components and biochemical parameters is also important in monitoring metabolic equilibrium and diagnosing disturbances in farming conditions [7]. Thus, the present study aimed at testing the potential use of CHO and LIP, added to diet at different levels, as protein sparing agents. This assessment was based on growth parameters and metabolic intermediate quantification in juvenile tambaqui.
Fish and environmental conditions
The experiment was performed in the Aquatic Organism Nutrition Laboratory at the Aquaculture Center of São Paulo State University (UNESP) in Jaboticabal, São Paulo state, Brazil. The procedures applied were in accordance with the animal research ethical principles established by the Brazilian College of Experimentation (COBEA) and approved by the Animal Research Ethics Committee (CEUA), protocol # 018492/12.
The 1080 juvenile tambaqui tested had an average weight of 10.88 g ± 0.13 g and average total length of 9.49 cm ± 0.24 cm. The fish were randomly distributed into twenty-four 500-L tanks (45 fish per tank) with constant air and water supply (~4.0 Lmin-1 renewal rate). Physicochemical water parameters were measured weekly and kept at a temperature of 29.68ºC ± 0.15ºC, dissolved oxygen of 5.85 mgL-1 ± 0.32 mgL-1, pH of 7.81 ± 0.7, total ammonia of 196.90 mgL-1 ± 40.76 μgL-1 [8] and total phosphorous of 42.85 mgL-1 ± 1565 μgL-1 [9].
Experimental diets
Six isoprotein diets (230 gkg-1 digestible protein) were produced, combining 3 CHO levels (410 gkg-1, 460 gkg-1 and 510 gkg-1 CHO) and 2 LIP levels (40 gkg-1 and 80 gkg-1 LIP). The diets were formulated based on the apparent digestibility coefficient of the ingredients, as determined in a previous study (unpublished data). The diets of each group were isocaloric, that is, crude energy of 17 MJkg-1 in diets containing 40 gkg-1 LIP and 18 MJkg-1 in those with 80 gkg-1 LIP) (Table 1).
| Ingredients (gkg-1) | Diets (Carbohydrates/Lipids) | |||||
|---|---|---|---|---|---|---|
| 410/40 | 460/40 | 510/40 | 410/80 | 460/80 | 510/80 | |
| Salmon meal (65% CP) | 191 | 191 | 191 | 191 | 191 | 191 |
| Corn gluten | 170 | 170 | 170 | 170 | 170 | 170 |
| Yeast | 20 | 20 | 20 | 20 | 20 | 20 |
| Corn | 145 | 145 | 145 | 145 | 145 | 145 |
| Sorghum | 50 | 50 | 50 | 50 | 50 | 50 |
| Corn starch | 190 | 270 | 340 | 210 | 280 | 350 |
| Cellulose | 199 | 119 | 49 | 140 | 70 | - |
| Corn Oil | 15 | 15 | 15 | 15 | 15 | 15 |
| Vitamin-mineral supplementa | 5 | 5 | 5 | 5 | 5 | 5 |
| Dicalcium phosphate | 11 | 11 | 11 | 11 | 11 | 11 |
| Antioxidant (BHT) | 1 | 1 | 1 | 1 | 1 | 1 |
| Antifungal (Filax) | 3 | 3 | 3 | 3 | 3 | 3 |
| Nutritional compositionb (gkg-1) | ||||||
| Dry matter | 928 | 928 | 928 | 928 | 928 | 928 |
| Crude protein | 260 | 260 | 260 | 260 | 260 | 260 |
| Digestible proteinc | 236 | 236 | 236 | 236 | 236 | 236 |
| Lipid | 40 | 40 | 40 | 80 | 80 | 80 |
| Mineral matter | 41 | 41 | 41 | 41 | 41 | 41 |
| Crude fiber | 171 | 111 | 58 | 127 | 74 | 22 |
| Starch | 316 | 386 | 447 | 333 | 395 | 456 |
| Carbohydrated | 413 | 466 | 513 | 418 | 464 | 510 |
| Crude energy (MJkg-1) | 17 | 17 | 17 | 18 | 18 | 18 |
| Digestible energyc (MJkg-1) | 15 | 15 | 15 | 16 | 15 | 15 |
| CE to CP ratioe (MJg-1) | 65 | 65 | 65 | 69 | 69 | 69 |
(a) Mineral mix (Premix Nutrifish Guabi®, Campinas/ SP, Brazil) per kg of product: folic acid (1250 mg); calcium pantothenate (12 000 mg); cupper (125 mg); iron (15 000 mg); iodine (375 mg); manganese (12 500 mg); selenium (87.5 mg); zinc (12 500 mg); cobalt (125 mg); vitamin A (2 500 000 IU); vitamin B12 (4000 mg); vitamin B1 (4000 mg); vitamin B2 (4000 mg); vitamin B6 (4000 mg); vitamin C (50 000 mg); vitamin D3 (600 000 IU); vitamin E (37 500); vitamin K3 (3750 mg); niacin (22 500 mg); biotin (15 mg); (b) Diet composition was calculated according to ingredient values; (c) Obtained from another study on digestibility for juvenile tambaquis (unpublished data); (d) Carbohydrate=dry matter - (crude protein+lipid+mineral matter+crude fiber); (e) CE=crude energy, CP=crude protein.
Table 1: Composition of test diets (based on natural matter).
The diets were produced in an extruder at the feed manufacturing facility of the Agricultural and Veterinary School of UNESP, Jaboticabal campus. Pellet size ranged from 4 mm to 6 mm. The diets were provided until apparent satiety 3 times a day, for 120 days. The amount of feed consumed in each tank was calculated weekly by the difference in food container weight before and after feeding.
Productive performance parameters
Fish from each tank were weighed and measured at the onset and end of the experiment to assess mean weight gain, apparent feed conversion, mean daily food intake and protein efficiency ratio, according to the following equations:
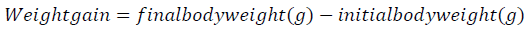
Feed conversion rate = mean feed intake (g)/mean weight gain (g)
Mean daily feed intake = feed intake per fish (g)/time (days)
Protein efficiency ratio = mean weight gain (g)/crude protein intake (g)
Morpho-somatic indices were determined at the end of the experiment. To that end, 3 fish from each tank were collected (12 fish per treatment), feed-deprived for 24 h and then killed by deepening anesthesia with 0.2 gL-1 benzocaine. Fish liver and visceral fat were removed and weighed to calculate the hepatosomatic index (HSI) and viscerosomatic fat index (VSI), as follows:
Hepatosomatic index (%) = (Wliver/Wtfish) x 100
Viscerosomatic fat index (%) = (Wfat/Wtfish) x 100
where: Wliver: mean liver weight; Wfat: mean visceral fat weight and Wtfish: mean fish weight.
Energy metabolism
To evaluate energy metabolism of CHO and LIP, 3 fish from each tank (12 per treatment) were collected at the end of the experimental period and feed-deprived for 24h. Fish were then anesthetized with 0.2 gL-1 benzocaine and blood was collected by caudal puncture. Blood samples were transferred to 1.5 mL centrifuge microtubes, half without anticoagulant and the other half with 15 μL EDTA solution+potassium fluoride (Glistab®). Samples with anticoagulant were centrifuged at 3000 rpm for 10 min to separate plasma and determine blood glucose levels using a commercial kit (GOD-Trinder method, Labtest® *Reference: 133a). Blood samples without anticoagulant were kept at room temperature for 3h and then centrifuged at 3000 rpm for 10 min to separate serum, which was used to determine triglycerides, and total protein and cholesterol levels, using Labtest® kits (*References: 87b, 99c and 13d, respectively). After blood collection, fish were killed by immersion in water and ice, and liver and muscle fragments were removed and stored at -20°C for future determination of tissue fat content [10] and liver glycogen [11]. Blood and tissue analyses were carried out at the Department of Animal Morphology and Physiology of the Agricultural and Veterinary Sciences School (FCAV) of UNESP, in Jaboticabal.
Experimental design and statistical analyses
The experiment was arranged in a randomized 3 × 2 factorial design, with 6 treatments combining 3 CHO and 2 LIP levels, with 4 replications per treatment. Analyses of variance (two-way ANOVA) of the results were tested for error normality and variance homoscedasticity (Cramer Von-Mises and Brown-Forsythe test, respectively). Significant statistical differences were compared by applying the Tukey test (p<0.05). Statistical analyses were performed using Statistical Analysis System SAS® v.9.0 software (SAS Institute Inc., Cary, North Carolina, USA).
The increased levels of CHO included in the diet reduced weight gain (Figure 1a) and daily food intake (Figure 1b), with the lowest means observed in fish fed 510/80 (CHO/LIP), although they did not differ from 510/40 and 460/80 treatments. Fish fed the 410/40 diet showed worse feed conversion (Figure 1c) and lower protein efficiency ratio (Figure 1d) than those fed the 460/80 diet. The other treatments exhibited intermediate values and neither value differed statistically. The results showed no significant differences as a function of LIP levels or interaction effect of the factors (p>0.05).

Figure 1: (a) Carbohydrate (CHO) and lipid (LIP) effects on weight gain (b) mean daily feed intake, (c) feed conversion rate, (d) protein efficiency ratio of juvenile tambaqui (mean ± SD). Uppercase letters compare the 6 treatments, and lowercase letters the LIP levels within each CHO level. Means followed by a same letter do not differ according to the Tukey test (p>0.05).
CHO and LIP levels did not affect HSI (Figure 2a) or VSI (Figure 2b). Blood glucose, total serum protein and liver glycogen levels did not differ statistically (p>0.05) as a function of CHO and LIP levels. With an increase in LIP levels to 80 gkg-1, serum cholesterol levels and fat deposition in the liver and muscle increased. CHO and LIP levels exhibited an interaction effect in serum triglyceride levels (Table 2).

Figure 2: (a) Hepatosomatic index (HSI), (b) viscerosomatic fat index (VSI) of juvenile tambaqui fed diet with different carbohydrate (CHO) and lipid (LIP) levels (mean ± SD). Uppercase letters compare the treatments, and lowercase letters the LIP levels within each CHO level. Means followed by a same letter do not differ according to the Tukey test (p>0.05).
| Factors | Blood glucose (mgdL-1) |
Serum triglycerides (mgdL-1) |
Serum total protein (gdL-1) |
Serum cholesterol (mgdL-1) |
Liver glycogen (%) |
Liver lipid (%) |
Muscle lipid (%) |
|---|---|---|---|---|---|---|---|
| Carbohydrate level | |||||||
| 410 gkg-1 | 58.96 ± 10.47 | 458.84 ± 172.93 | 2.69 ± 0.26 | 160.81 ± 41.61 | 12.98 ± 1.54 | 6.80 ± 0.41 | 7.69 ± 4.51 |
| 460 gkg-1 | 64.63 ± 7.20 | 486.75 ± 128.82 | 2.67 ± 0.13 | 193.41 ± 46.95 | 13.40 ± 1.47 | 7.16 ± 1.00 | 7.80 ± 4.26 |
| 510 gkg-1 | 63.86 ± 6.64 | 826.67 ± 351.56 | 2.72 ± 0.22 | 197.69 ± 43.65 | 13.66 ± 1.42 | 7.25 ± 0.60 | 8.01 ± 3.40 |
| Lipid level | |||||||
| 40 gkg-1 | 63.35 ± 8.55 | 518.87 ± 180.37 | 2.63 ± 0.17 | 163.68 ± 31.54 b | 13.50 ± 1.61 | 6.77 ± 0.54 b | 4.95 ± 1.81 b |
| 80 gkg-1 | 61.86 ± 8.06 | 645.73 ± 358.59 | 2.76 ± 0.22 | 204.26 ± 49.15 a | 13.19 ± 1.29 | 7.37 ± 0.76 a | 10.98 ± 2.96 a |
| Effects (F-value; ANOVA) | |||||||
| Carbohydrate (CHO) | 1.93NS | 16.01** | 0.14NS | 2.23NS | 0.41NS | 1.01NS | 0.14NS |
| Lipid (LIP) | 0.67NS | 9.32** | 2.06NS | 6.78* | 0.24NS | 4.73* | 37.69** |
| CHO x LIP interaction | 4.39NS | 8.02** | 0.56NS | 1.65NS | 0.99NS | 0.63NS | 2.33NS |
| Coefficient of variation (%) | 11.38 | 38.52 | 7.83 | 20.74 | 11.29 | 9.46 | 30.38 |
Different letters in a column indicate a significant difference according to the Tukey test (p<0.05); NS=non- significant; *=p<0.05 (Tukey test); **=p<0.01 (Tukey test).
Table 2: Energy metabolism (mean ± SD; N=4) and corresponding F-values of juvenile tambaqui diets containing different carbohydrate and lipid levels.
In treatments with 40 g kg-1 LIP, an increase in CHO levels did not affect serum triglyceride levels. In the group fed diets containing 80 gkg-1 LIP, however, the inclusion of 510 gkg-1 CHO increased mean triglyceride levels to 1231.31 mgdL-1 ± 116.80 mgdL-1, which is 2 to 3 times higher than those obtained in treatments with 460 and 410 gkg-1 CHO, respectively. In animals fed diets with 510 gkg-1 CHO, triglyceride levels also increased when LIP rose from 40 g to 80 g kg-1. This difference was not observed when 410 gkg-1 and 460 gkg-1 of CHO was added (Table 3).
| Lipid (gkg-1) | Carbohydrate (gkg-1) | |||
|---|---|---|---|---|
| 410 | 460 | 510 | ||
| Serum triglyceride | 40 | 509.58 ± 209.77 aA | 422.67 ± 101.35 aA | 624.35 ± 194.22 bA |
| (mg dL-1) | 80 | 408.09 ± 137.52 aB | 572.20 ± 123.33 aB | 1231.31 ± 116.80 aA |
Different lowercase letters in a column and uppercase letters in a row indicate a significant difference according to the Tukey test (p<0.05).
Table 3: Interaction effect of diet carbohydrate and lipid levels on serum triglyceride levels (mean ± SD) of juvenile tambaquit.
The lower weight gain of fish fed the 510/80 (CHO/LIP) diet (Figure 1a) likely resulted from the significant decrease in food intake (Figure 1b), which reduced the consumption of essential nutrients such as proteins. The decreased growth was possibly caused by enzymatic limitation when CHO is used as energy source [12], particularly in the treatments containing 510 gkg-1 CHO.
A reduction in voluntary feed intake after an increase in diet energy with CHO and LIP was reported for the juvenile rainbow trout Oncorhynchus mikiss [13], juvenile largemouth bass Micropterus salmoides [14], piracanjuba Brycon orbignyanus [15], jundiá Rhamdia quelen [16] dorado Salminus brasiliensis [8] and dentex Dentex dentex [17]. In the present study, weight gain and food intake tended to improve when diet CHO increased from 410 gkg-1 to 460 gkg-1 and worsen when it rose from 460 gkg-1 to 510 g kg-1 in diets containing 40 gkg-1 LIP (Figure 1a and 1b).
The results of the present study showed that an increase in CHO to 460 gkg-1, improved feed conversion and increased the protein efficiency ratio, without affecting mean daily food intake and weight gain. A similar food conversion and protein efficiency ratio was observed in another omnivorous fish (Piaractus mesopotamicus ) fed diets containing 410 gkg-1, 460 gkg-1 or 500 gkg-1 CHO [18]. However, high CHO levels in the diets can promote fat deposition by stimulation of lipogenic enzymes in fish liver [19]. In the present study, this phenomenon was manifested by an increase in serum triglyceride levels in treatments containing an increase in CHO to 510 gkg-1 (Table 3).
According to Hemre et al. [20], diet CHO promotes fish growth if the amount added does not exceed the limits that meet metabolic demands and does not result in intolerance or nutrient deficiency. Wang et al. [21] tested corn starch at six inclusion levels (60, 140, 220, 300, 380 and 460) gkg-1 in juvenile Nile tilapia (Oreochromis niloticus x O. aureus ) diets and found that fish can use up to the highest level without compromising growth, but fish fed diets with the lowest levels (60 gkg-1 and 140 gkg-1) used protein as energy source, and not for carcass deposition.
One of the main functions of liver is to sequester blood glucose, which is stored as glycogen and lipid and can be hydrolyzed and released when required [22]. According to Table 2, blood glucose and liver glycogen did not change with an increase in CHO and LIP content in diet, suggesting that blood glucose was available to supply the energy requirements for growth and was not stored as liver glycogen. Accordingly, Corrêa et al. [23] did not detect changes in blood glucose levels in tambaqui fed diets containing different CHO levels. In the present study, blood glucose homeostasis is related to the absorption of diet CHO. Other studies show the adjustment capacity of digestive and metabolic enzymes in response to different dietary CHO sources and levels [23-25]. A noteworthy aspect is that before blood sampling the fish tested were feed deprived for 24 h, and omnivorous and herbivorous fish are known to be able to regulate an increase in blood glucose in a few hours. This blood glucose dynamic is related to the ability of some fish to achieve glycemic homeostasis, as observed in Piaractus mesopotamicus [26]. Likewise, matrinxã (Brycon cephalus ) was found to exhibit high blood glucose levels up to 24 h after being fed, and reduced levels after feeding restriction for 72 h [27]. A recent study showed that even the carnivorous rainbow trout (Oncorhynchus mykiss ) can regulate an increase in blood glucose within a few hours, which is much faster than previously reported, and that it can also completely halt the production of liver glucose [28].
According to Wilson [29], diet CHO is digested slowly, which may result in a stable glycemic response and improved growth. Rawles et al. [30] however, report that improved weight gain (increase in body mass and decrease in body fat) has been difficult to achieve in fish fed large amounts of CHO because control of glycemic homeostasis is associated with several factors such as insulin secretion [31] occurrence of insulin receptors in the cells [32], receptor type [33] and the presence of glucose carriers and enzymes involved in glucose metabolization [31,33].
Similarly, no statistical differences in total serum protein levels were detected (Table 2), possibly because the diets were isoprotein. This was also observed by Ren et al. [34] in an evaluation of different CHO diet sources for blunt snout bream (Megalobrama amblycephala ).
An increase in diet LIP to 80 gkg-1 enhanced the deposition of liver and muscle fat and increased blood cholesterol levels (Table 2). In salmonids and mammals, adipose is the main tissue used for fat storage, but other fish use the liver and muscle as storage sites [35]. The relative importance of these storage sites varies among species. No statistical difference was found in hepatosomatic index (HSI) or viscerosomatic fat index (VSI) (Figure 2a and 2b), confirming that tambaqui deposits fat primarily in the liver and muscle rather than in the mesenteric adipose tissue, since these are more important storage sites in fish [36], and because of the high muscle lipid levels detected (4% to 10%).
According to Araújo-Lima et al. [37], muscle fat in farmed tambaqui ranges from 2% to 6%. Diet LIP content directly affects the chemical composition of fish and therefore, plays an important role in the final quality of the meat, especially in the muscle. Depending on the species, season, nutrition status and farming system, muscle fat can range from 2% to more than 20%, and from a human nutrition standpoint, fish can be classified as lean (<2% fat), slightly fat (2%-4%), moderately fat (4%-8%) and very fat (>8%) [38].
As in mammals, cholesterol in fish can be obtained from the diet or synthetized, primarily in the liver and the intestine [39]. Under normal feeding conditions, endogenous cholesterol is directly regulated by diet cholesterol levels. As such, serum cholesterol concentrations can vary according to the nutritional status of fish [40]. Therefore, the increase in serum cholesterol detected in the present study indicates a more active endogenous LIP transport in response to the increase of this nutrient in the diet.
The increase in serum triglyceride observed in the 510/80 (CHO/ LIP) treatment (Table 3) suggests the occurrence of lipogenesis through the synthesis of hepatic triglycerides followed by transport to plasma and, finally, incorporation in storage tissues (liver, muscle and adipose). This result suggests that higher LIP levels in the diet compromised the use of the fat consumed, channeling triglycerides to storage tissues, especially when CHO level increased to 510 gkg-1. In this respect, Tocher [41] reports that the availability of substrates derived from glucose metabolism favors the synthesis of triglycerides that are subsequently enveloped in lipoprotein particles in the liver, and then proceed to extra-hepatic tissues such as adipose and muscle tissue. Although many studies show that fat deposition is related to diet LIP levels, optimum inclusion levels have yet to be established, especially with respect to the quality of the final product.
Although tambaqui is a tropical frugivorous-herbivorous species, the inclusion of LIP and CHO in its diet must consider inclusion limits in order to maximize protein use, because higher levels of CHO and LIP are related to a decline in performance parameters, affect triglyceride levels and increase fat deposition in storage tissues. In the present study, however, the CHO and LIP proportions tested did not compromise performance, allowing fish to efficiently use protein for growth. The findings suggest that the best balance was obtained with diets containing 40 gkg-1 LIP and 460 gkg-1 CHO, which did not impaired growth or cause excess fat accumulation.
To the São Paulo Research Foundation (FAPESP) for fellowship (grant #2012/15656-6) and regular financial support (grant #2013/00254-2).