Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2014) Volume 2, Issue 4
Twenty–six moderately halophilic bacteria were isolated from the sea coast of Sanya city Hainan province China, and screened for their ability to secrete halophilic proteases. Among them, a moderately halophilic strain Pontibacillus sp. SY–8 was investigated for the production of extracellular protease. 16S rRNA gene analysis identified it as Pontibacillus chungwhensis. To enhance protease production, Plackett–Burman (PB) design and response surface methodology (RSM), were used to determine the optimal concentrations. Based on PB design experiments, casein, yeast extract powder, and NaCl were determined as the main factors influencing protease production. The optimal concentration of these components was determined by response surface methodology, which is based on the central composite design. Under the proposed optimized conditions, the protease experimental yield (1441 U ml-1) closely matched the yield predicted by the statistical model (1472.59 U ml-1) with R2 =0.990. An overall 3.56–fold increase in protease production was achieved in the optimized medium as compared with the unoptimized medium.
<Keywords: Moderately halophilic bacterium; haloalkaliphilic protease; Optimization; Response surface methodology Moderately halophilic bacterium; haloalkaliphilic protease; Optimization; Response surface methodology
Microbial proteases are important class of enzymes, which constitute more than 65% of the total industrial enzyme market. A challenge faced by industrial application of microbial proteases is optimal activity and stability in a wide range of salinity. Halophilic proteases, which are active both at low and high concentrations of NaCl, are needed. Given their potential use, there has been renewed interest in the discovery of proteases with novel properties [1-4].
Halophiles are microorganisms which grow over an extended range of salt concentrations (3–30% NaCl, w/v), include the halophilic bacteria and archaea. They are found in salt marshes, marine ecosystems, salted meats, hypersaline seas, salt evaporation pools and salt mines [5]. Moderately halophilic bacteria grow well over a wide range of salt concentrations, 3-15% [6], and sometimes even at close to saturated NaCl [5]. Their exoenzymes exhibit unique structural, biochemical characteristics, and stable under extreme high saline conditions [7-9]. The exoenzymes from halophiles are not only interesting from the basic scientific viewpoint, but they may also be of potential interest in many industrial and biotechnological applications, owing to their stability and activity at low water levels.
Although an optimal strain is important for the application of protease, culture medium components and fermentation conditions play a significant role in protease production [10,11]. In recent years, the use of statistical approaches involving PB design and RSM has gained momentum in medium optimization. These methods are also useful for understanding interactions among various physicochemical parameters using a minimal number of experiments. The PB design allows for the screening of the main factors from a large number of variables, and this information can be retained in further optimization. RSM is a collection of statistical techniques that is useful for designing experiments, building models, evaluating the effects of different factors and searching for optimal conditions of studied factors for desirable responses. RSM has been successfully applied in many areas of biotechnology, such as laccase production and protease production [12-14].
In view of that, we isolated a moderately halophilic bacterium (SY- 8) which produced haloalkaliphilic protease and aimed to optimize the significant variables on its protease production.
Samples
For the isolation of halophilic microorganisms, seawater and sediment samples were collected from was isolated from the sea coast of Sanya city Hainan province China (109°28′E, 18°12′N).
Culture medium and culture conditions
Seed culture medium consisted (g L–1): glucose 10, yeast extract powder 5, NaCl 100, KCl 2.0, (NH4)2SO4 1.0, pH 7.5. Culture medium was sterilized at 121° for 20 min. Solid culture medium: add 15 g L–1 agar in Seed culture medium.
Casein agar plate: proteolytic production of the bacterial strains was screened on agar plates supplemented with adding 10 g L–1 casein in solid culture medium. The plates were incubated overnight at 30 °C. The protease producing strains were selected based on the zone of clearance.
Fermentation medium contained (g L–1): casein 5, glucose 10, yeast extract powder 5, NaCl 100, KCl 2.0, (NH4)2SO4 1.0, pH 7.5, sterilization at 121 °C for 20 min.
Isolation of moderately halophilic bacteria for protease production
Isolation of moderately halophilic bacteria was done by samples employing seed culture medium, the cultures were diluted serially and plated on solid culture medium. The pure isolates were picked up and streaked on casein agar plate. Isolates showing clear zones of proteolytic activity were selected as protease producer moderately halophilic bacteria. Fermentation medium was inoculated with 5% inoculum (v/v) (A600nm = 1.05). Erlenmeyer flasks (250 mL) containing 50 mL of the medium were used in both seed and fermentation medium. Fermentation medium was maintained at the same conditions for 72 h.
Transmission electron microscopy (TEM)
The cells of log phase were harvested by centrifugation at 10,000×g for 10 min at 4°C. The pellets were washed with 0.1 M phosphate buffer (pH7.5). The washed cells were fixed in modified Karnovsky’s fluid and processed as per the procedure of David et al [15].
Identification of the bacterial strain
The protease producing strain was identified by various biochemical test and 16S rRNA gene sequencing. Genomic DNA was extracted as per the standard protocol and it was amplified by using the following universal 16S rRNA primers: 27F(5′-AGAGTTTGATCMTGGCTCAG–3′), 1492R(5′-GGYTACCTTGTTACGACTT–3′) [16]. 16S rRNA gene sequences obtained from strain SY–8 was compared with other bacterial sequences by using NCBI Mega BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) for their pair wise identities. It was related to Pontibacillus by 16S rRNA gene sequencing and submitted in GenBank, NCBI, USA.
Multiple alignments of these sequences were carried out by Clustal W 1.83 version of EBI (http://www.ebi.ac.uk/cgi–bin/clustalw/). Phylogenetic tree was constructed in MEGA 5.0 version (http://www.megasoftware.nte) using neighbor–joining (NJ) method with a bootstrap value of 1000 [17].
Protease assay
Protease activity was measured by caseinolytic assay [18]. The suitably diluted enzyme (0.5 ml) was added to 3.0 ml casein (0.6%, w/v in 20 mM borax–NaOH buffer, pH 10.0, containing 5%, w/v NaCl). The mixture was incubated at 30 °C for 20 min. The reaction was stopped by adding 3.2 ml of TCA mixture (0.1 M trichloro acetic acid, 0.2 M sodium acetate, 0.3 M acetic acid) and incubating the mixture at 30 °C for 30 min. The resultant precipitates were removed by filtration through Whattman filter paper and absorbance of the filtrate was measured at 280 nm. One unit of protease activity was defined as the amount of enzyme liberating 1 μg of tyrosine min–1 under assay conditions. Enzyme units were calculated using tyrosine standard curve made by 0–100 μg ml–1 tyrosine in 20 mM glycine–NaOH buffer, pH 10.0. A blank was run in the same manner, except the enzyme was added after the addition of TCA mixture.
Screening of critical media components using the PB design
The PB design allows the evaluation of N variables in N+1 experiments; each variable was examined at two levels: –1 for a low level and +1 for a high level. In the present study, the following variables were selected: glucose, casein, yeast extract powder, NaCl, KCl, (NH4)2SO4, pH, and each factor had two levels in the experimental design (Table 1).
| Symbol code | Variables | Units | Level | |
|---|---|---|---|---|
| −1 | +1 | |||
| A | glucose | g/l | 10 | 20 |
| B | NaCl | g/l | 50 | 100 |
| C | Dummy | − | − | − |
| D | casein | g/l | 10 | 20 |
| E | yeast extract powder | g/l | 5 | 10 |
| F | Dummy | − | − | − |
| G | pH | − | 7.0 | 7.5 |
| H | KCl | g/l | 1 | 2 |
| J | Dummy | − | − | − |
| K | (NH4)2SO4 | g/l | 0.5 | 1 |
| L | Dummy | − | − | − |
Table 1: Experimental variables at different levels used for lipase production by SY–8 using the PB design.
Optimization of medium using RSM
The three independent factors were studied at five different levels (−1.682, −1, 0, +1, and +1.682). The model was statistically analyzed to evaluate the analysis of variance (ANOVA). Quality of the polynomial model equation was statistically confirmed by the coefficient of determination, R2, and its statistical significance was determined by F test. Significance of the regression coefficients was tested by t test.
Validation of the experimental model
The statistical model was validated with respect to all the three variables within the design space. Experiments predicted by the point prediction feature of the Design Expert software were conducted in triplicates. Six combinations of medium constituents were used to determine lipase production, and the results were compared with the predicted values.
Isolation, screening, and identification of protease producer
In this study, moderately halophilic bacteria from the sea coast of Sanya city Hainan province China, have been screened for the presence of protease activity. A total of twenty−six halophilic isolates were purified by salt enrichment from the water samples of the offshore. The protease producers among the isolates were identified by casein agar plates. Out of these, four identified strains namely, SY−8 (GenBank accession no. JN016852), WJ5−1 (GenBank accession no. JN624832), WJ5−3 (GenBank accession no. JN624835) and WJ6−1 (GenBank accession no. JN624834) showed 405, 337, 210 and 305 U/ml protease activity in fermentation medium, respectively. SY−8 was found to be Gram positive rods, strictly aerobic. It grew at 15–40 °C (optimum, 30 °C), pH 6.0–10.0 (optimum, pH 7.0–7.5) and 1–15% (w/v), NaCl (optimum, 5–10%). The transmission electron micrographs (TEM) of the isolate clearly exhibited rod–shaped morphology with peritrichous flagella on the cell surface (Figure 1). Biochemical tests showed positive reactions for catalase, and negative for oxidase, nitrate reduction.
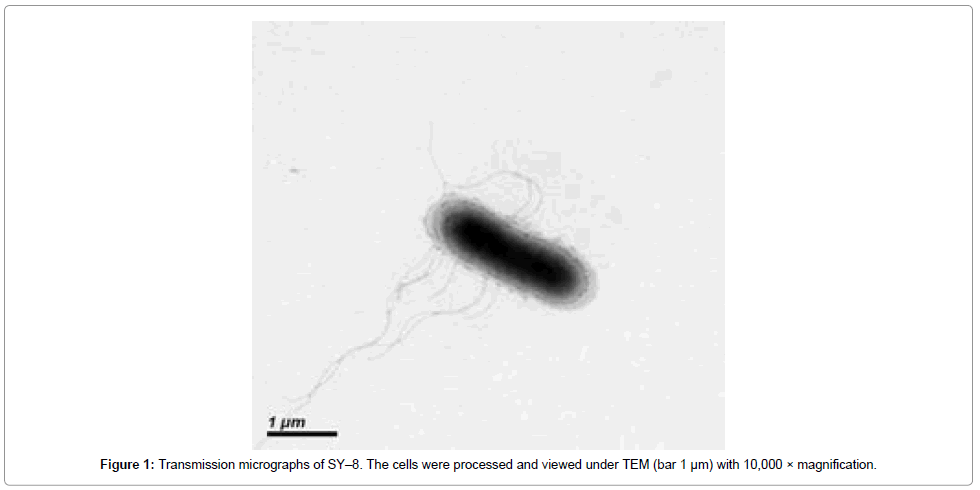
Figure 1: Transmission micrographs of SY–8. The cells were processed and viewed under TEM (bar 1 μm) with 10,000 × magnification.
Based on the morphological, biochemical characters and 16S rRNA gene sequence showed that the strain SY–8 belonged to the genus Pontibacillus. The phylogenetic tree (Figure 2) constructed by the NJ method indicated that the isolate SY–8 was part of the cluster within the genus Pontibacillus [19].
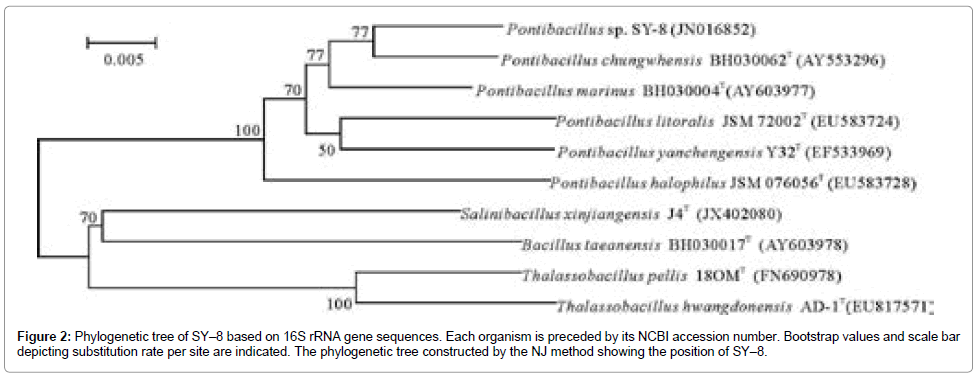
Figure 2: Phylogenetic tree of SY–8 based on 16S rRNA gene sequences. Each organism is preceded by its NCBI accession number. Bootstrap values and scale bar depicting substitution rate per site are indicated. The phylogenetic tree constructed by the NJ method showing the position of SY–8.
Characterization of SY–8 protease
The purified protease by SY–8 was confirmed to be single purified protein molecule by using SDS–PAGE (data not shown). The protease activity was observed in all the tested concentrations of NaCl and this enzyme requires 5–10 NaCl for maximum activity. In the control group, it retained only 73.5% activity. At concentrations between 1–5% the protease activity retained above 80%, and at concentrations above 10% the protease activity drastically decreased (Figure 3A). The effect of pH on protease activity was investigated with varying pH range of 5-12 and the result showed that pH9–10 was the optimum range and it was especially high at pH10 (Figure 3B). From the result, it was confirmed that the protease was a haloalkaliphilic protease.
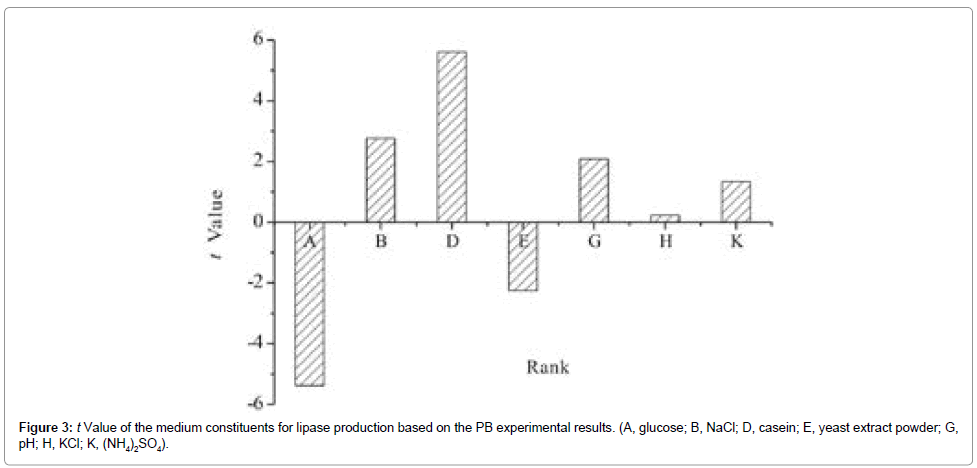
Figure 3: t Value of the medium constituents for lipase production based on the PB experimental results. (A, glucose; B, NaCl; D, casein; E, yeast extract powder; G, pH; H, KCl; K, (NH4)2SO4).
Selection of the most suitable carbon and nitrogen sources by the one–variable–at–a–time approach
A series of experiments were carried out to study the effects of various simple and complex carbon and nitrogen sources on protease production by SY–8. Cultures were first conducted using fermentation culture medium. In addition, the different carbon sources were added at a concentration of 10 g L–1. SY–8 produced a high level of protease in the presence of complex organic carbon sources with glucose (data not shown). The maximum levels of protease activity were carried out with glucose (405 U ml–1). Easily assimilated simple carbon sources like fructose, lactose, sucrose and maltose resulted in weak protease production, 125, 132, 270 and 145 U ml–1, respectively.
In general, both organic and inorganic nitrogen sources were used efficiently for protease production. In the present study, different organic (yeast extract powder, casein, soy peptone, and pastone) and inorganic ((NH4)2SO4 and NH4Cl) nitrogen sources, at a concentration of 5 g L–1, were tested in fermentation culture medium containing glucose at 10 g L–1 as the carbon source. Higher protease production levels were obtained with casein (310 U ml–1) compared to yeast extract powder, soy peptone, and corn syrup (284, 230, and 232 U ml–1, respectively). Inorganic nitrogen sources ((NH4)2SO4 and NH4Cl) showed weak enzyme production levels, 165 and 112 U ml–1, respectively (data not shown). Complex nitrogen sources (casein, yeast extract powder, and (NH4)2SO4) showed high enzyme production levels (405 U ml–1). Among the various carbon and nitrogen sources tested, glucose, casein, yeast extract powder, and (NH4)2SO4 were the most suitable substrates for the production of proteolytic enzymes. Thus, these substrates were selected for further optimization steps.
Screening of significant factors using the PB design
To optimize protease production, a total of seven medium components were screened using the PB design through 12 experimental runs with two levels each (Table 2). Design Expert® 8.0.6 (Stat−Ease, Inc., Minneapolis, USA) was used to analyze the experimental PB design. The PB design serves as a valuable tool for initial screening to determine the effects of various factors in a small number of experiments for reliable short−listing of relevant factors, indicating how each factor affects the production process for further optimization [20].
| Run | A | B | C | D | E | F | G | H | I | J | K | Protease activity (U/ml) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | −1 | 1 | −1 | |
| 2 | −1 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | −1 | 1 | |
| 3 | 1 | −1 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | −1 | |
| 4 | −1 | 1 | −1 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | |
| 5 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | 1 | 1 | 1 | −1 | |
| 6 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | 1 | 1 | 1 | |
| 7 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | 1 | 1 | |
| 8 | 1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | 1 | |
| 9 | 1 | 1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | −1 | |
| 10 | −1 | 1 | 1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | 1 | |
| 11 | 1 | −1 | 1 | 1 | 1 | −1 | −1 | −1 | 1 | −1 | 1 | |
| 12 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 |
Table 2: The PB experimental design for evaluation of 11 components with coded values for protease production by SY–8.
The t value of the effects of each medium component was performed (Figure 4), which serve as a measure to view the contributions of individual component to production. This was estimated on the basis of the difference between the average of measurements taken at the high level (+1) and the low level (−1) of each component. The medium components affecting lipase production was ranked as casein > glucose > NaCl > yeast extract powder > pH >(NH4)2SO4> KCl.
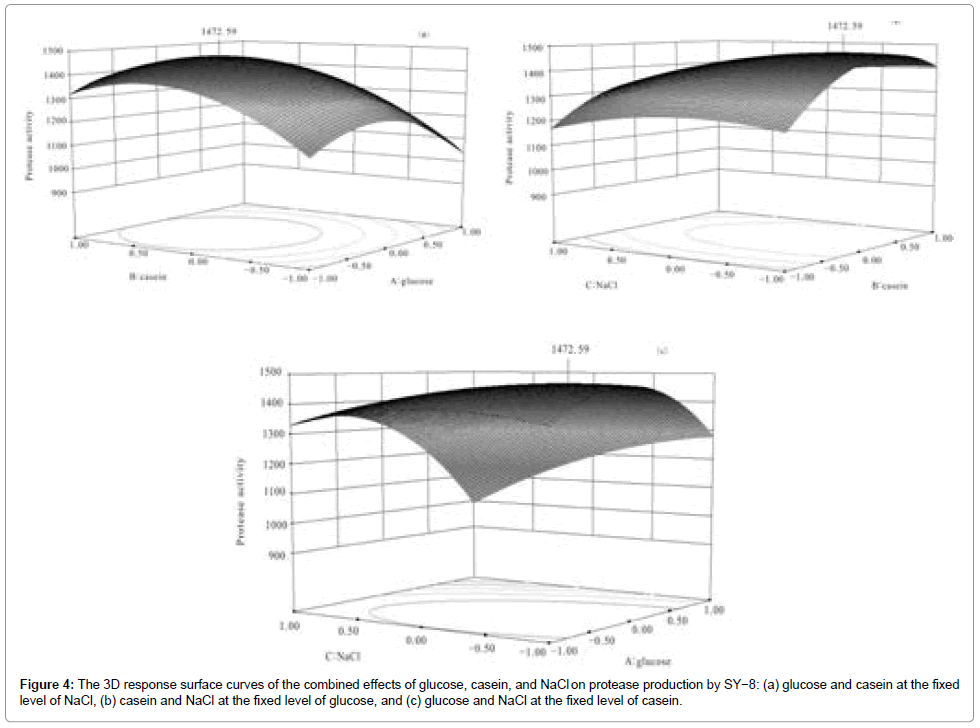
Figure 4: The 3D response surface curves of the combined effects of glucose, casein, and NaCl on protease production by SY−8: (a) glucose and casein at the fixed level of NaCl, (b) casein and NaCl at the fixed level of glucose, and (c) glucose and NaCl at the fixed level of casein.
It is clear that glucose and yeast extract powder affect protease production at negative levels, and casein, NaCl, pH, (NH4)2SO4, and KCl affect production at positive levels. Based on the design, protease activity ranges from 205 U ml–1 to 1196 U ml–1, indicating a strong influence of medium components on enzyme production. The highest activity was found in run 10 where all the medium components were at the level with the same affect level of protease production; run 5 showed the lowest activity of 205 U ml–1. This could be related to the absence of casein which was the inducer of protease production.
The standard ANOVA results were calculated from the experimental runs (Table 3). The model F−value (11.32) for lipase activity implied that the model was significant, and P (0.0168) indicated that the model terms were significant. Casein, glucose, and NaCl were selected as significant model terms on the basis of ANOVA results. Moreover, the value of coefficient of determination (R2) for the factorial model was 0.952. The predicted R–squared values were in reasonable agreement with the adjusted R−squared values for all equations. Consequently, on the basis of these results, the three most significant variables were further evaluated using RSM.
| Variable | Sum of Squares | Coefficient Estimate | Mean Squares | Standardized Effects | F value | P | |
|---|---|---|---|---|---|---|---|
| Code | Factors | ||||||
| model | 1.454 E+006 | 563.25 | 2.077 E+005 | 11.32 | 0.0168 * | ||
| A | glucose | 5.238 E+005 | −208.92 | 5.238 E+005 | −417.83 | 28.56 | 0.0059 |
| B | NaCl | 1.485 E+005 | 111.25 | 1.485 E+005 | 222.50 | 8.10 | 0.0466 |
| D | casein | 5.664 E+005 | 217.25 | 5.664 E+005 | 434.50 | 30.88 | 0.0051 |
| E | yeast extract powder | 1.007 E+005 | −91.58 | 1.007 E+005 | −183.17 | 5.49 | 0.0792 |
| G | pH | 82502.08 | 82.92 | 82502.08 | 165.83 | 4.50 | 0.1012 |
| H | KCl | 520.08 | 6.58 | 520.08 | 13.17 | 0.028 | 0.8744 |
| K | (NH4)2SO4 | 31518.15 | 51.25 | 31518.15 | 102.50 | 1.72 | 0.2601 |
Table 3: Statistical analysis of medium components for protease production using the PB design.
Optimization using RSM
On the basis of the PB experiments, CCD under RSM was used to determine the optimal conditions for the three significant factors: glucose, casein, and NaCl. For each run, a total of 20 experiments with different combinations of the three selected variables were performed (Table 4). The results demonstrated that the maximal amount of protease from SY−8 (1412 U ml–1 to 1448 U ml–1) is produced from the runs representing the center points (runs 15, 16, 17, 18, 19, and 20); run 8 showed minimum production (843 U ml–1).
| Run | Factors | Protease activity (U/ml) | |||
|---|---|---|---|---|---|
| (g/l) | (g/l) | (g/l) | Experimental | Predicted | |
| 1 | 8(−1) | 20(−)1 | 80(−1) | 1131 ± 6.3 | 1122.35 |
| 2 | 12(1) | 20(−1) | 80(−1) | 1010 ± 4.1 | 1041.41 |
| 3 | 8(1) | 30(1) | 80(−1) | 1277 ± 3.5 | 1302.86 |
| 4 | 12(1) | 30(1) | 80(−1) | 1235 ± 11.5 | 1217.67 |
| 5 | 8(−1) | 20(−1) | 100(1) | 1136 ± 5.0 | 1155.35 |
| 6 | 12(1) | 20(−1) | 100(1) | 864 ± 6.7 | 840.41 |
| 7 | 8(−1) | 30(1) | 100(1) | 1262 ± 12.9 | 1232.45 |
| 8 | 12(1) | 30(1) | 100(1) | 843 ± 4.7 | 883.51 |
| 9 | 6.636(−1.682) | 25(0) | 90(0) | 1173 ± 3.3 | 1187.84 |
| 10 | 13.364(1.682) | 25(0) | 90(0) | 869 ± 3.9 | 851.54 |
| 11 | 10(0) | 16.59(−1.682) | 90(0) | 926 ± 2.5 | 933.73 |
| 12 | 10(0) | 33.41(1.682) | 90(0) | 1157 ± 3.7 | 1146.67 |
| 13 | 10(0) | 25(0) | 73.18(−1.682) | 1398 ± 10.3 | 1398.36 |
| 14 | 10(0) | 25(0) | 106.82(1.682) | 1173 ± 6.8 | 1170.18 |
| 15 | 10(0) | 25(0) | 90(0) | 1441 | 1431.24 |
| 16 | 10(0) | 25(0) | 90(0) | 1422 | 1431.24 |
| 17 | 10(0) | 25(0) | 90(0) | 1429 | 1431.24 |
| 18 | 10(0) | 25(0) | 90(0) | 1448 | 1431.24 |
| 19 | 10(0) | 25(0) | 90(0) | 1412 | 1431.24 |
| 20 | 10(0) | 25(0) | 90(0) | 1435 | 1431.24 |
Table 4: Experimental design using CCD of three independent variables with their actual and coded values and six center points showing the experimental and predicted responses.
A second–order regression equation showed the dependence of SY–8 protease activity on the medium constituents. The parameters of the equation were obtained by multiple regression analysis of the experimental data. An empirical relationship between the response and the screened variables was expressed in terms of the second–order polynomial equation:
Y (Protease activity, U ml–1) = 1431.24 – 99.97A + 63.30B – 67.83C – 8.50AB – 66.00AC − 33.25BC – 145.47A2 – 138.22B2 – 51.95C2
The response (Y) represents the amount of lipase and A, B, and C are the concentrations of glucose, casein, and NaCl, respectively. The experimental results revealed that this mathematical tool could satisfactorily explain the effect of the concentration of the variables (glucose, casein, and NaCl) on protease production by SY–8 in shake flask cultures. ANOVA for the Quadratic model is shown in Table 5. The model variables P of <0.05 implies that the model is significant [21]. In this case A, B, C, AC, BC, A2, B2, and C2 are significant model terms. The lack of fit P of 0.0157 showed that the lack of fit is significant.
| Source | Sum of square | df | Coefficient Estimate | Mean Squares | F value | P |
|---|---|---|---|---|---|---|
| Model * | 8.364 E+005 | 9 | 1431.24 | 92935.27 | 110.30 | <0.0001 |
| A | 1.365 E+005 | 1 | −99.97 | 1.365 E+005 | 161.99 | <0.0001 |
| B | 54723.45 | 1 | 63.30 | 54723.45 | 64.95 | <0.0001 |
| C | 62841.94 | 1 | −67.83 | 62841.94 | 74.59 | <0.0001 |
| AB | 578.0 | 1 | −8.50 | 578.0 | 0.69 | 0.4268 |
| AC | 34848.0 | 1 | −66.00 | 34848.0 | 41.36 | <0.0001 |
| BC | 8844.50 | 1 | −33.25 | 8844.50 | 10.50 | 0.0089 |
| A2 | 3.049 E+005 | 1 | −145.47 | 3.049 E+005 | 361.93 | <0.0001 |
| B2 | 2.753 E+005 | 1 | −138.22 | 2.753 E+005 | 326.77 | <0.0001 |
| C2 | 38894.48 | 1 | −51.95 | 38894.48 | 46.16 | <0.0001 |
| Residual | 8425.49 | 10 | 842.55 | |||
| Lack of Fit | 7574.65 | 5 | 1514.93 | 8.90 | 0.0157 | |
| Pure Error | 850.83 | 5 | 170.17 | |||
| Cor Total | 8.448 E+005 | 19 |
Table 5: ANOVA for the Quadratic model.
R2 closer to 1 denotes a high correlation between the observed and predicted values. The higher R2 (0.9900) and adjusted R2 (0.9811) values also indicate the efficacy of the model, suggesting that 99.0% and 98.11% variation could be accounted for the model equation, respectively. Thus, for a good statistical model, R2 should be in the ranges of 0–1.0; the closer the value is to 1.0, the better fit the model is deemed to be [22]. Adequate precision measures the signal–to–noise ratio and a value of >4 are considered appropriate for desirable models. The adequate precision value of 28.794 for lipase production indicates that the model can be used to navigate the design space. Moreover, the coefficient of variation (CV) measures the degree of precision with which the treatments are compared, and the low CV values indicate the reliability of the experiment. A relatively low CV value (2.41) in this study suggests a good precision and reliability. P is used as a tool to check the significance of each coefficient, which also indicates the strength of the interaction between each independent variable; the smaller the P, the better the significance of the corresponding coefficient. The responses revealed that all of the three quadratic coefficients are significant (P < 0.05) and have remarkable effects on the overall production.
The regression equation, maximized by Design Expert 8.0.6, allowed us to obtain the following optimal coded units with a high model accuracy for test variables: A = −0.21, B = 0.31, and C = −0.62, corresponding to the optimal concentrations of 9.58 g L–1 glucose, 26.55 g L–1 casein, and 83.8 g L–1 NaCl for the maximal lipase activity of 1472.59 U ml–1.
The 3D plots (Figure 5) were based on the function of concentration of two variables with the other variable being at its optimal level. Significance of the interaction between the corresponding variables is indicated by an elliptical or saddle nature of the contour plots [23]. The shape of the response surface curves indicates a moderate interaction between the tested variables. Thus, it can be concluded that the optimal combination of the selected medium components exhibit strong synergistic effects on lipase production. Medium manipulation offers a better alternative for enzyme overproduction, because secretion of metabolism products is an important component of the survival strategy among some microorganisms under certain environments [24].
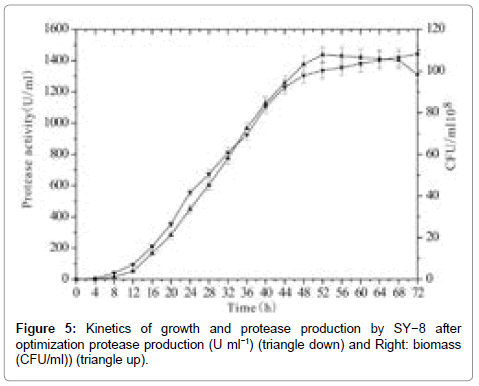
Figure 5: Kinetics of growth and protease production by SY−8 after optimization protease production (U ml−1) (triangle down) and Right: biomass (CFU/ml)) (triangle up).
Validation of the model
To further verify the predicted results, a set of experiments with three replicates in each was performed according to the media constituents (Table 6). Some of the experimental results were slightly higher than the predicted results. This minor deviation may likely be due to experiments conducted in the border zone of the technological space considered in CCD. After optimization, a successful and significant improvement (3.56−fold) in the production of protease by SY−8 was accomplished using the PB design and RSM.
| Experiment | Glucose (g/l) | Casein (g/l) | NaCl (g/l) | Protease activity (U/ml) | |
|---|---|---|---|---|---|
| Experimental | Predicted | ||||
| 1 | 9.58 | 26.55 | 83.8 | 1441 ± 11.6 | 1472.59 |
| 2 | 12 | 25 | 100 | 1033 ± 8.3 | 1000.02 |
| 3 | 10 | 25 | 100 | 1257 ± 4.7 | 1311.46 |
| 4 | 12 | 27.5 | 90 | 1147 ± 9.5 | 1178.39 |
| 5 | 9 | 30 | 90 | 1385 ± 6.8 | 1374.19 |
| 6 | 9 | 12 | 4 | 1274 ± 11.2 | 1290.53 |
Table 6: Validation of the experimental model.
Time course of protease production and cell growth during the fermentation
The time course of protease activity and the growth of SY-8 for the optimized (casein 26.55, glucose 9.58, yeast extract powder 5, NaCl 83.8, KCl 2.0, (NH4)2SO4 1.0, pH 7.5) conditions are shown in Figure 5. These results showed that the secretion of the protease was associated with the cell growth. Protease activity increased during the exponential growth phase and reached activity of 1305 U ml−1 at 48 h. The production of proteases increased slightly at the beginning of the stationary phase. By optimizing the medium composition, the production of halophilic protease by SY−8 was enhanced from 405 to 1441 U ml−1.
It is highly important to discover new bacterial−strains that produce enzymes with novel properties that could be of industrial value. Enzymes from halophiles are considered novel for industrial application because of their catalytic ability at high salt environment and low water requirement. The present study introduces high levels of haloalkaliphilic protease by a newly isolated moderately halophilic bacterium SY−8. Using RSM, we got an optimal culture medium for protease production by SY−8. After optimization, the protease production was increased from 405 U ml−1 to 1441 U ml−1, the highest yield has been reported by moderately halophilic bacterium.
This work was financially supported by Hi–Tech Research and Development Program of China (863 program of China; 2012AA092103), China Ocean Mineral Resources R&D Association (DY125–15–T–06).