Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2015) Volume 3, Issue 1
Anthropogenic inputs influence the community structure and activities of microorganisms, which may impinge the functioning of estuarine and coastal ecosystem. The aim of the present study was to understand the influence of dissolved heavy metals (Cr, Mn, Fe, Ni, Cu and Zn) on community structure of cyanobacteria in a nutrient rich tropical estuary, Cochin Estuary (CE), across the southwest coast of India. Dissolved heavy metals were higher in CE during dry season, with Zn as major pollutant. The level of dissolved Zn was higher in stations near river head during dry (130 μg L-1) and wet seasons (105 μg L-1). The community structure of cyanobacteria was higher in CE during wet season (H’ index: 1.4 to 3.1) compared to that in dry season (H’ index: 1.1 to 2.2). The 16S rRNA gene sequence analysis showed the prevalence of Synechococcus sp in CE. PCA analysis indicated the positive influence of Zn and nutrients on community structure of cyanobacteria. We postulate that the Zn pollution may be a critical factor to be monitored and controlled in nutrient rich estuaries, otherwise it may accelerate the eutrophication and associated ecological constraints.
<Keywords: Heavy metal; Cochin estuary; Eutrophication; Cyanobacteria; Phytoplankton
Influence of heavy metals on the community structure and activities of microorganisms in aquatic systems, varies depending on their concentration and available chemical forms [1,2]. At optimum concentration, heavy metals like Cu, Zn and Mn support major life processes such as maintenance of osmotic balance, stabilization of cellular organelles, they function as electron acceptors in many processes and cofactors for metallo-enzymes [3-5]. The heavy metal pollution can induce long lasting effects on health of estuaries as they are not easily or rapidly eliminated from these ecosystems by natural degradative processes. Estuaries, being the transition zone between fresh- and marine-water systems, function as receiving points of heavy metals and other pollutants of anthropogenic origin and regulate their supply to the coastal environment. Heavy metal pollution in estuaries has increased significantly in the recent past through enhanced discharge of industrial and domestic wastes [6-9]. Previous studies showed that heavy metal pollution causes unintended alterations in the functioning of estuarine ecosystems [10-13]. The direct impact of heavy metal pollution on microbial ecosystem includes the alterations in the physiology, diversity and abundance of microorganisms, which indirectly affect the biogeochemical cycles and ocean productivity [10-13].
Cyanobacteria are the major picoplankton distributed in estuarine environment, where they occupy a key position at the base of the food web and play a central role in carbon, nutrient and oxygen cycling [14]. The cyanobacteria Prochlorococcus and Synechococcus, alone are responsible for an estimated 20-40% of global carbon fixation, although they account for only one percent of photosynthetic biomass [15]. In spite of this, the effect of metal pollution on diversity of cyanobacteria is least studied from estuarine environments and most of the reported studies on heavy metal–cyanobacteria interactions have been conducted in small volume culture flasks in laboratories [2]. Field level observations that include the whole microbial food web are important to understand the impact of heavy metal pollution on cyanobacterial diversity. In a pristine environment heavy metals are transported across the cell membrane through energy independent or dependent pathways to participate in cellular processes. If this open gate condition pertains under polluted environment, heavy metals may accumulate in cytoplasm leading to toxic effects at cellular and subcellular level. Interestingly, microorganisms acquired several mechanisms to maintain the homeostasis between the available metal concentration and microbial physiology [12,16,17]. In oligotrophic marine environments, cyanobacteria secrete ligands that bind with biologically important metals like copper, iron, cobalt and zinc [18]. The laboratory scale experiments showed that trace metals above optimum levels can influence the general growth, pigment composition, photosynthesis and enzyme expression of cyanobacteria [19]. Several strains of cyanobacteria have shown the ability to accumulate, detoxify or metabolize heavy metals up to certain concentration range [20]. Cyanobacteria are effective biological metal sorbents, which represent an important sink for metals in aquatic environments and have the ability to control the speciation of many biologically active metals [20-22]. Although metal accumulation is a promising strategy for the survival of cyanobacteria in metal polluted environments, their impact on the food web will be disturbing.
Cochin Estuary (CE) is the largest wetland ecosystem along the southwest coast of India, covering an area of ~25600 ha, extending from 9° 30’-10° 12’ N to 76° 10’-76° 29’ E. CE is reported to be nutrient rich, and its composition is greatly influenced by anthropogenic and terrestrial inputs from six rivers, seawater influx from two bar mouths and prolonged monsoon [23]. CE also receives high concentrations of industrial effluents (104 × 103 m3 per day) and untreated domestic waste (260 m3). Like most estuaries, the anthropogenic contribution of heavy metals has increased significantly in CE over the past three decades [9]. However, the influence of heavy metal pollution on diversity of photosynthetic microorganisms are not studied till date. In a previous study we reported the influence of heavy metal pollution on the diversity and enzyme expression profile of heterotrophic bacteria in CE [9]. In the present study, we report the influence of dissolved heavy metals on the community structure of cyanobacteria in CE.
Sample collection
Water samples were collected from four locations across a pollution gradient in CE using 5 L capacity Niskin samplers during dry (February) and wet (September) season. The positions of the sampling stations, S1, S2, S3 and S4 are given in Figure 1. S1 is situated near the Cochin bar mouth and is designated as least polluted station. S4 is designated as a grossly polluted station in this study and located near the point where the river Periyar, carrying effluents, joins the estuary. S2 and S3 are considered as intermediately polluted stations.
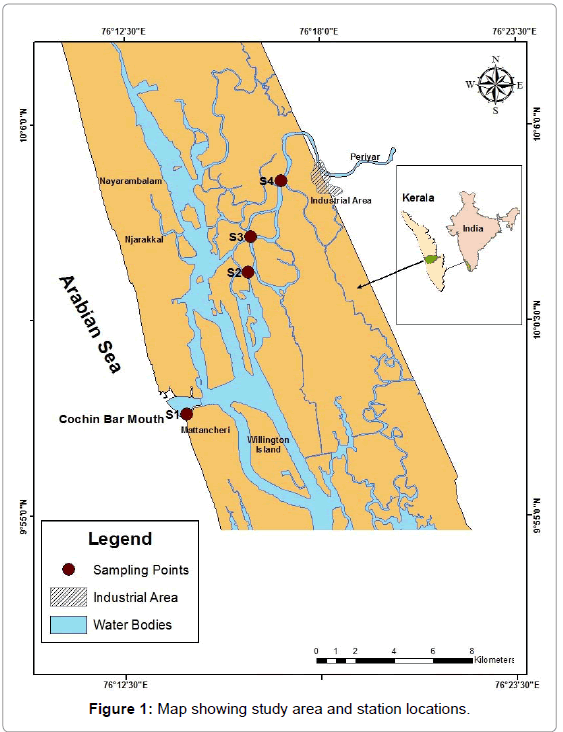
Figure 1: Map showing study area and station locations.
Analysis of environmental parameters
Environmental variables were measured following standard protocol. Salinity was determined using a Digi Auto Salinometer (Model TSK, accuracy ± 0.001) and the pH using an ELICO LI 610 pH meter. Dissolved oxygen content in the ambient water was determined following Winkler’s titration method [24]. Samples for nutrients (ammonia, nitrite, nitrate, phosphate and silicate) were filtered and estimated spectrophotometrically within six hours of sampling [24]. Suspended particulate matter (SPM) was collected on a pre-combusted 0.45 μ GF/F filter paper (Whatman, USA) and measured gravimetrically after achieving constant weight at 70°C.
Analysis of heavy metals
Water samples were analyzed for dissolved heavy metals, Cr, Mn, Fe, Ni, Cu and Zn, using inductively coupled plasma optical emission spectroscopy (ICP-OES) following standard protocol [24]. Heavy metals present in water samples (400 ml) were acidified using concentrated nitric acid. The dissolved metals were extracted using ammonium pyrrolidine dithiocarbamate (APDC, 2%) and chloroform at acidic pH, brought back to aqueous layer by back-extraction with concentrated nitric acid and made up to 20 ml with sterile di-ionized water. The precision and accuracy of the analytical procedure was monitored by the repeated analysis of standard reference material BCSS-1.
Community structure of cyanobacteria
The community structure of cyanobacteria in CE was studied using nested PCR-DGGE technique. Genomic DNA was extracted from water samples following standard protocol with slight modification [25]. Here the water samples (2 L) were passed through 0.2 μ polycarobonate membrane filter (Milliipore; GTTP2500), followed by incubation at 37°C for 1 hr in lysis buffer (NaCl 400 mM, Sucrose 750 mM, EDTA 20 mM and Tris HCl 50 mM) containing 1 mgml-1 lysozyme. Subsequently, SDS (1%) and proteinase K (100 μgml-1) were added to the solution and incubation was continued for 5 hr at 55°C. Further DNA was precipitated with 0.6 volume of isopropanol at -20°C for 60 min. DNA pellet was washed copiously with 70% ethanol, dissolved in TE buffer and stored at -20°C until used.
Approximately 2000 bp of 16S rRNA gene of cyanobacteria were amplified using primers 359F (GGGGAATTTTCCGCAATGGG) and 23S30R (CTTCGCCTCTGTGTGCCTAGGT). The genomic DNA was amplified with 10 picomol each of forward and reverse primers in a 25 μl reaction volume containing 1 μl of DNA, 2.5 μl 10 × Taq polymerase buffer (NEB, Canada), 0.5 U Taq DNA polymerase (NEB, Canada), and 200 μM each dNTP (Sigma Aldrich, USA). The reaction mixture was subjected to thermal cycling conditions as follows: initial denaturation at 95°C for 2 min, followed by cycle denaturation at 95°C for 40 sec, annealing at 58°C for 40 sec, and extension at 72°C for 15 sec for a total of 30 cycles. Nested PCR was done using cyanobacterial specific DGGE primers CYA359F and an equimolar combination of reverse primers, CYA781R (a) (GACTACTGGGGTATCTAATCCCATT) and CYA781R (b) (GACTACAGGGGTATCTAATCCCTTT), following PCR conditions of initial denaturation at 95°C for 2 min, followed by cycle denaturation at 95°C for 40 sec, annealing at 58°C for 40 sec, and extension at 72°C for 2 min for a total of 30 cycles. The resulting 400bp PCR products were run on a denaturing gradient gel (30-50% formamide) prepared in 8% polyacrylamide for 17.5 hr at constant voltage of 75 V in 1X TAE buffer (40 mM tris-HCl, 20 mM acetic acid, 1 mM EDTA). The bands formed in the gel were stained with SYBR green and observed in a gel documentation system (BioRAD, USA). The bands were picked, incubated overnight in a 50 μl TE and re-amplified. The PCR products were treated with EXOSAP-IT (Affymetrix, Ohio USA) and used as template for sequencing on an ABI sequencer. Sequence data obtained were analyzed and edited using Sequencher V4.10.1 (GeneCodes, USA). Subsequently, the sequences were compared with those in the GenBank database using the Basic Local Alignment Search Tool (BLAST) algorithm to determine approximate phylogenetic affiliations. The nucleic acid sequences showing the closest similarities were used as Reference sequences while constructing a neighbor -joining tree using the software MEGA (version 5.0). DGGE approach will give an insight into the overall distribution of cyanobacteria in the CE. We acknowledge the limitation of DGGE which originated from efficiency of DNA extraction methodology, selective amplification of genes from mixed DNA samples and low resolution in the band pattern of phylogenetically related species. Maximum care was taken to keep these errors to the minimum possible levels. Despite these shortcomings, the PCR-DGGE approach has been proved to be a powerful method allowing a comprehensive picture of the community structure and constraints associated with it.
Statistical analysis
Difference in band pattern between stations was compared using Bionumerics software. The software carried out a density profile analysis, detected the bands from each lane and calculated the relative contribution of each band to the total lane intensity. Numbers of Operational Taxonomic Units (OTUs) in each sample were counted as number of DGGE bands. Gels were cross-checked visually as well as for number of bands per lane. The relative intensity of each band was used to calculate the Shannon Wiener diversity index (H’ index). Cluster analysis of DGGE bands based on square root transformed community data matrix through Bray-curtis similarity were performed. Multivariate statistics, Principal Components Analysis (PCA), was carried out to understand the influence of environmental variables and heavy metals on DGGE band diversity of cyanobacteria. All statistical analysis were performed with the software Statistica (Version 6.0).
Environmental variables in CE
We observed higher levels of nutrients in CE during wet season (Table 1). Ammonia was the major nutrient in CE and its level ranged from 1.9 to 5.2 and 4.8 to 11.3 μmol L-1 respectively in dry and wet season. Nitrate (4.4. to 19.3 μmol L-1), silicate (31 to 36.1 μmol L-1) and phosphate (1.3 to 3.3 μmol L-1) were also higher in the CE during wet season compared to dry season (nitrate 0.5 to 1.4, silicate 8.5 to 36.3 and phosphate 1.05 to 3.9 μmol L-1). The level of dissolved nitrogen (sum of ammonia, nitrite and nitrate) are increased four to six times in the estuary during wet season. Nutrient levels are reported to be higher in tropical estuaries during wet season, and is attributed to increased river water discharge and rainfall associated with southwest monsoon [26-28]. CE receives 60–70% of its annual freshwater discharge i.e., 22.41 × 103 Mm3 during southwest monsoon [29], which carries the nitrogen rich effluents discharged from fertilizer and fish processing industries [30]. It is estimated that 260 million litre per day of effluents are produced in the industrial belt of Cochin and a significant fraction of it reaches CE through river Periyar [31]. Also, 235 million litre per day of untreated sewage are generated in Cochin city and a significant fraction of this may reach the estuary through land runoff and other nonpoint sources [30]. CE has been recognized as a nutrient rich estuary and the nutrient levels are always maintained at sufficient levels to support the nourishment of phytoplankton [32]. The nutrient levels in the estuary are comparable with Godavari estuary, India [33], Carolina’s Neuse river estuary (NRE) USA [34], Seine estuary in France [35] and Schelde estuary in Belgium [36]. Higher SPM levels in CE during dry seasons was attributed earlier to high seawater influx and mixing of water column [37]. The distribution of nutrients in the estuary are directed by seasonal changes in river water discharge and seawater influx. CE is dominated by seawater during dry season and its influence was recorded up to station S4 (salinity 15) (Table 1), while it remains limnetic during wet season due to heavy river water inflow and the influence of seawater was restricted at station S2 (salinity 9). The average river water discharge was reported to be minimum in CE from January to May in a year, it increases in June with the onset of southwest monsoon and reaches its plateau during August [29]. Accordingly the CE shifts between seawater and fresh water dominance in dry and wet season respectively.
| Environmental factors | ||||||||
|---|---|---|---|---|---|---|---|---|
| Dry season | Wet season | |||||||
| S1 | S2 | S3 | S4 | S1 | S2 | S3 | S4 | |
| pH | 7.8 | 7.8 | 7.9 | 7.8 | 7.8 | 7.8 | 7.6 | 7.1 |
| Salinity | 34.5 | 32 | 8 | 15 | 30 | 9 | 0 | 0 |
| SPM(mg L-1) | 32.4 | 26.8 | 23.2 | 17.6 | 36 | 18 | 6.8 | 2.8 |
| NH3 (μmolL-1) | 1.9 | 2.5 | 3.2 | 5.2 | 11.3 | 7.9 | 16.1 | 4.8 |
| NO2 (μmolL-1) | 0.2 | 0.1 | 0.1 | 0.2 | 0.5 | 0.3 | 0.3 | 0.2 |
| NO3 (μmolL-1) | 1 | 0.5 | 1.4 | 0.5 | 4.4 | 6 | 13.6 | 19.3 |
| SiO4 (μmolL-1) | 10.3 | 8.5 | 36.3 | 22.6 | 31 | 56.7 | 35.4 | 36.1 |
| PO4 (μmolL-1) | 1.9 | 1.6 | 1.05 | 3.9 | 3.3 | 2.1 | 2.4 | 1.3 |
Table 1: Environmental variables observed in Cochin estuary during the study period.
Heavy metal pollution in CE
The concentrations of dissolved heavy metals in the water column of CE are given in Table 2. The dissolved heavy metals were higher in CE during dry season, which may be attributed to changes in flushing rate. Flushing activity would be inactive in Indian estuaries during dry season, while it experisences multiple flushing in addition to heavy rain fall during wet season [38]. In concurrence with this, the level of dissolved heavy metals were found higher during dry season and lower during wet season. Although the fresh water influx and industrial discharge are higher during wet season, the high flushing rate, i.e., one flushing in 1-2.5 day [39], will result in the dilution of dissolved heavy metals in CE during wet season. We recorded higher levels of Zn in stations near to the river head i.e., S3 and S4 during dry (48. 28 and 130.5 μgL-1 respectively) and wet season (73. 65 and 105.85 μgL-1 respectively). The S4 is located geographically close to the industrial belt of Cochin and functions as discharge point of effluents carried by river Periyar. The level of heavy metal pollution gradually reduces from S4 at river mouth to S1 at bar mouth, owing to its accumulation in sediment [9]. Earlier studies recorded accumulation of heavy metals in the sediment samples and benthic organisms collected from CE [9,23,40-42]. In a previous study, we recorded an 18 times increase in the level of Zn in sediments of CE during the past three decades [9]. Although the heavy metals accumulated in sediment samples can cause serious problems to both estuarine and human health, phytoplanktons are most affected by dissolved heavy metals. Since the purpose of this work was to understand the impact of heavy metal pollution on community structure of cyanobacteria and the level accumulated in sediments are recorded in previous studies, only the dissolved heavy metals are monitored in the present study.
| Heavy metal | Concentration (μg L-1) at different stations | |||||||
|---|---|---|---|---|---|---|---|---|
| Dry season | Wet season | |||||||
| S1 | S2 | S3 | S4 | S1 | S2 | S3 | S4 | |
| Cr | 0.22 | 0.08 | 0.34 | 0.2 | 0.47 | 0.07 | 0.09 | 0.09 |
| Cu | 1.28 | 0.38 | 1.74 | 0.81 | 0.68 | 0.27 | 0.44 | 0 |
| Fe | 262.3 | 274.2 | 161.7 | 55.7 | 799 | 45.37 | 68.1 | 6.62 |
| Mn | 0.24 | 0.21 | 0.39 | 0.12 | 0.63 | 0.13 | BDL | BDL |
| Ni | 3.55 | 2.97 | 4.21 | 2.02 | 4.56 | 4.57 | 5.51 | BDL |
| Zn | 8.84 | 6.55 | 48.28 | 130.5 | 4.36 | 6.36 | 73.65 | 105.85 |
Table 2: Distribution of heavy metals in sampling stations during dry and wet season.
The level of Fe and Mn showed interesting fluctuations in CE. Although their levels were higher during dry season, maximum concentrations of Fe and Mn were recorded in S1 during wet season. The higher levels of Mn and Fe during dry season was reported earlier in CE [43], and was attributed to the higher salinity, which facilitates the formation of comparatively soluble Fe and Mn sulphides [44]. Under normal conditions, the heavy metals discharged to the estuary are immobilized in the sediment or accumulated in benthic organisms. Disturbing the system by either physical or chemical forces, i.e., resuspension of sediments by dredging operation or tidal mixing or adjusting the pH or redox conditions by discharging acidic wastes, may trigger the release of significant amount of heavy metals to the water column [45-47]. The resuspension of sediment due to continuous dredging operations could be the reasons for pulsed level of heavy metals in S1 during wet season. Previous studies placed CE among the most impacted estuaries in the world based on high levels of Zn, and recorded sediment as a sink of heavy metals [48].
Community structure of cyanobacteria in CE
We used standard PCR-DGGE technique to identify the community structure of cyanobacteria in CE. PCR DGGE have many advantages over morphological or culture dependent methods in analyzing the community structure of cyanobacteria. The morphological methods mainly depend on the size and pigmentation of cyanobacteria, which may vary with the nutrient compositions while culture dependent method is limited by the lack of a universal media which can accommodate all cyanobacteria. In PCR-DGGE, the PCR product of each species from the sample is segregated into distinct bands in a gradient gel [49]. Hence each band in a gradient gel can be considered as an operational taxonomic unit (OTU). Twenty representative DGGE bands were eluted, cloned in TOPO cloning vector and sequenced. The sequences were submitted to NCBI GenBank (accession numbers KP694048 – KP694067). The number of OTUs of cyanobacteria in the water column of CE was higher during wet season (4- 8) compared to dry season (2-4) (Figure 2). The Shannon Weiner diversity index calculated from the DGGE image also showed comparatively higher diversity of cyanobacteria in CE during wet season. During this season, maximum Shannon Weiner diversity index was observed in S4 (3.1), followed by S1 and S2 (1.8) and S3 (1.4). During dry season, maximum diversity of cyanobacteria was observed in S1 (H’ 2.2) followed by S2 (H’ 1.6). The lowest diversity of cyanobacteria was recorded in samples collected from S3 and S4 (H’ 1.1) during dry season. Based on the similarity of PCR–DGGE band pattern, the stations were clustered differently during dry and wet season (Figure 2). During dry season, S3 remained as a different cluster while other stations showed nearly 40% similarity in DGGE band pattern. The dissimilarity in cyanobacterial diversity may be due to the difference in both environmental variables and heavy metals at different stations in the estuary. DGGE band pattern of S1 and S2 showed more than 60% similarity during wet season, while S2 and S3 remained different clusters. The phylogenetic tree derived from PCR-DGGE analysis showed the presence of Synechococcus sp in all the stations (Figure 3). This is in agreement with a previous culture dependent study from CE [50]. A recent study based on pigment fluorescence also confirmed Synechococcus sp as a dominant picoplankton in CE [51]. Synechococcus sp has been reported from many estuaries and bays [52,53] and its bloom has been reported in Chesapeake Bay during summer when the salinity was high [52]. The Synechococcus sp has been recorded as a dominant contributor of biomass and phytoplankton productivity in nutrient rich North Carolina’s Neuse River Estuary (NRE) [34]. Our study also confirm the prevalence of Synechococcus sp in nutrient rich CE.
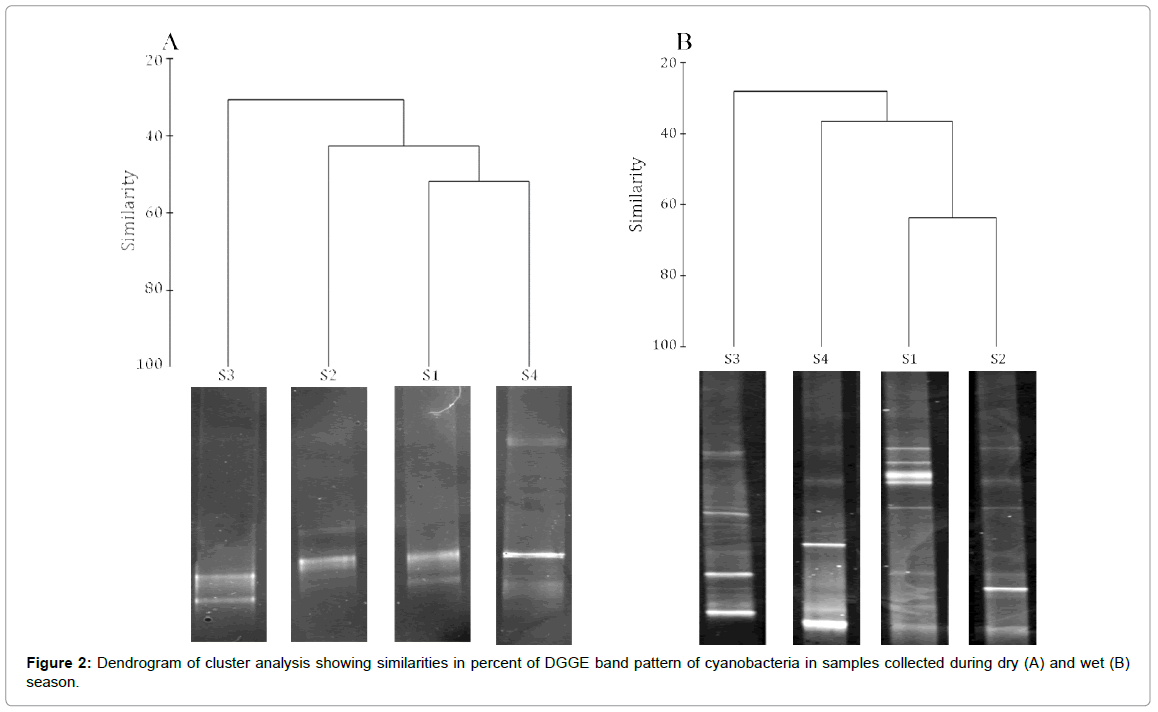
Figure 2: Dendrogram of cluster analysis showing similarities in percent of DGGE band pattern of cyanobacteria in samples collected during dry (A) and wet (B) season.
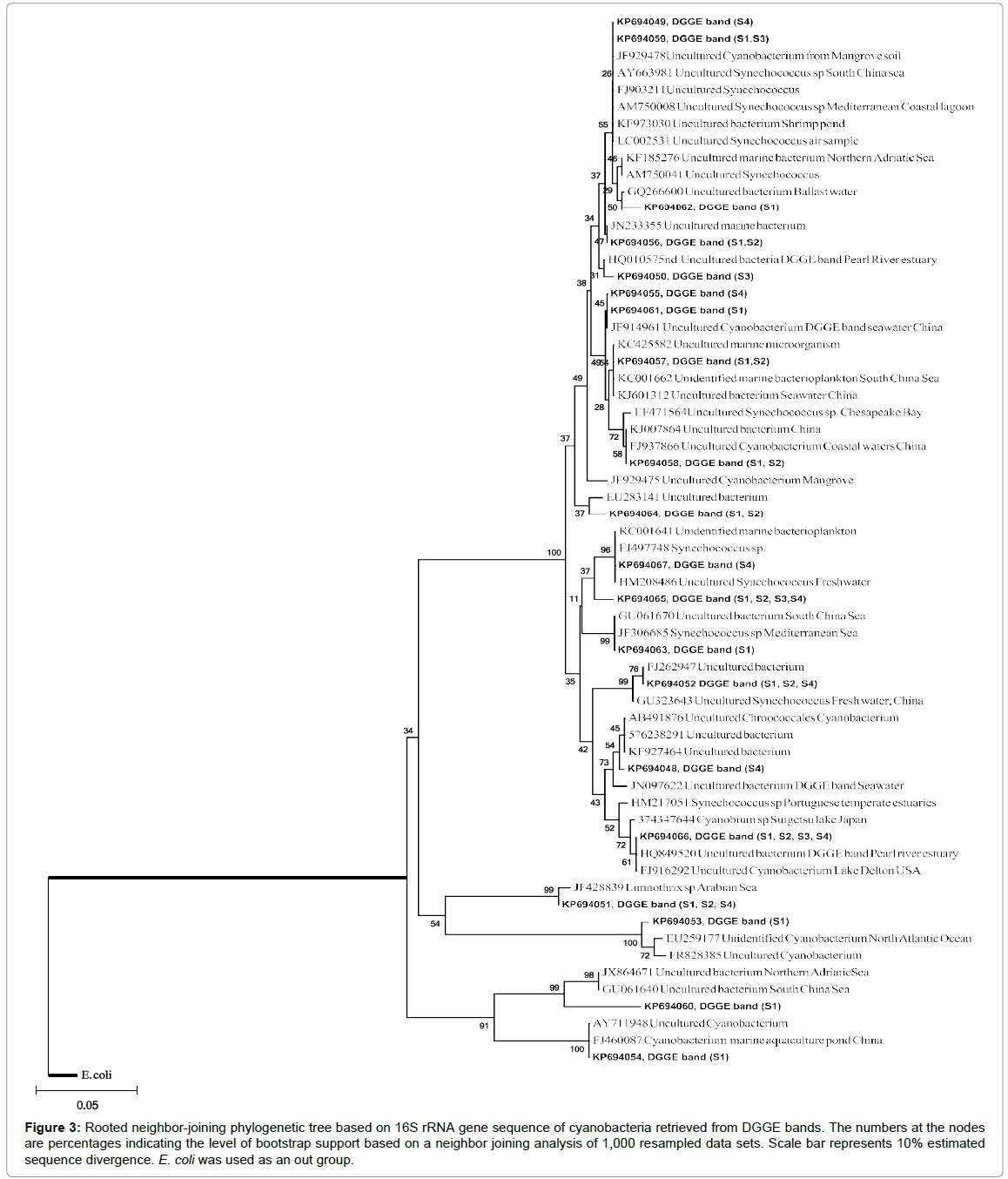
Figure 3: Rooted neighbor-joining phylogenetic tree based on 16S rRNA gene sequence of cyanobacteria retrieved from DGGE bands. The numbers at the nodes are percentages indicating the level of bootstrap support based on a neighbor joining analysis of 1,000 resampled data sets. Scale bar represents 10% estimated sequence divergence. E. coli was used as an out group.
Here we used three 16S rRNA gene primers specific for cyanobacteria Cya359F, Cya781R (a) and Cya781R (b), for the assessment of community structure of cyanobacteria in CE. The reverse primer Cya781R(a) preferentially targets filamentous cyanobacteria whereas Cya781R(b) targets unicellular cyanobacteria [49]. Interestingly, the 16S rRNA gene sequence derived from majority of bands (13 number) were matched with previously uncultivated cyanobacteria sequence data available in NCBI. This happens because a majority of the cyanobacteria are still uncultivated in laboratory condition and hence its taxonomical positions are not assigned completely. This indicates the need for further studies in isolation and taxonomical classification of cyanobacteria in marine environments.
Effect of heavy metal pollution on community structure of cyanobacteria
The PCA analysis showed that the community structure of cyanobacteria in CE was influenced by the levels of heavy metals and nutrients (Figure 4). Ammonia and nitrate are reported as the preferable source of nitrogen for natural phytoplankton communities in estuaries and their influence on phytoplankton productivity is well documented from different estuaries across the world [54-57]. CE is reported as a nutrient rich estuary and the nutrients are always maintained at sufficient levels to support the growth of phytoplankton [32,58]. Therefore the variations in level of dissolved Zn may function as the selective pressure on community structure of cyanobacteria in CE. Previous studies also supported our hypothesis that availability of Zn in the water column influences the community structure of cyanobacteria in marine and estuarine environments [59,60]. Zn is required at the active site of carbonic anhydrase, as it catalyses the conversion of HCO3- to CO2, and hence plays a pivotal role in photosynthetic carbon synthesis [61]. The available concentration of Zn is very low in many parts of the oceanic environment compared to that of estuaries [59] and the Zn absorption mechanisms of oceanic cyanobacteria are well reviewed [15,60]. However, very little is known about the response of cyanobacteria to elevated Zn levels in the estuarine environment. It is possible that up to certain levels the Zn promotes the growth and activities of cyanobacteria, but different cyanobacteria will have varying tolerance limits. Further mesocosm studies are therefore required to understand the tolerance limit of different groups of cyanobacteria present in CE to elevated Zn. Mesocosm studies using Synechococcus PCC 7942 demonstrated the synthesis of a 56 aminoacid cysteine rich metal binding protein, SmtA, which binds and sequesters metal ions within the cells and thereby maintains the homeostasis between the metal concentration and cyanobacterial viability [61]. Beyond tolerable limits, the Zn may out compete less competitive metal ions such as Fe and Mn for their protein binding sites [18]. In a mesocosm study, it was reported that Zeaxanthin level in the water collected from Godavari River decreased with increasing Zn concentration from 1 × 10-7 to 2.5 × 10-7 M [62]. We observed maximum diversity of cyanobacteria in CE at S4 during wet season, when the levels of nitrate, silicate and Zn were high. Although the Zn level increased further at S4 during dry season, the diversity of cyanobacteria was low, which may be due to the comparatively low levels of nitrate and silicate at that time or due to the toxicity of Zn. High levels of primary productivity and chlorophyll contents were also reported from S4 during wet season (i.e., June to September), and was attributed to high enormous inputs of nutrients [63].
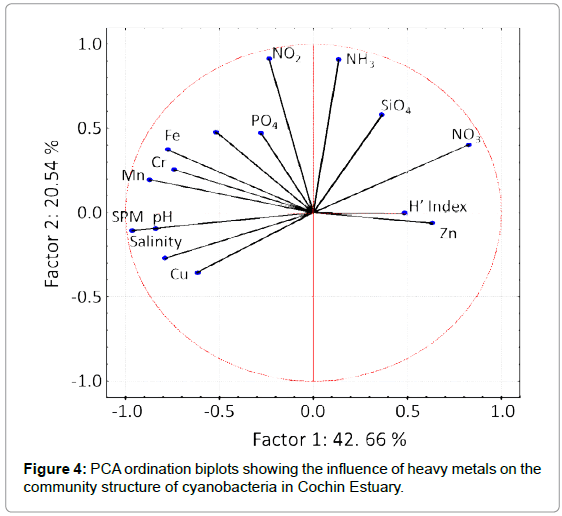
Figure 4: PCA ordination biplots showing the influence of heavy metals on the community structure of cyanobacteria in Cochin Estuary.
Although our results are specific with CE, the influence of Zn pollution on community structure of cyanobacteria and its implications regarding environmental impacts are of significant to tropical estuaries. In CE the Zn levels are high near the river end and is attributed to the anthropogenic input. Synechococcus sp was recorded as the dominant cyanobacteria in CE. The present study postulates that the anthropogenic inputs of Zn may impart critical control on the community structure of cyanobacteria in nutrient rich tropical estuaries like CE. In short, the present study points towards the importance of controlling Zn pollution in CE, as it may otherwise impinge the community structure of cyanobacteria, which may leads to unintended alterations in the functioning of estuarine ecosystem. Further studies to elucidate the tolerance limit of cyanobacteria to different heavy metals and the influence of metal pollution on photosynthetic efficiency of cyanobacteria will be appreciated.
Although our results are specific with CE, the influence of Zn pollution on community structure of cyanobacteria and its implications regarding environmental impacts are of significant to tropical estuaries. In CE the Zn levels are high near the river end and is attributed to the anthropogenic input. Synechococcus sp was recorded as the dominant cyanobacteria in CE. The present study postulates that the anthropogenic inputs of Zn may impart critical control on the community structure of cyanobacteria in nutrient rich tropical estuaries like CE. In short, the present study points towards the importance of controlling Zn pollution in CE, as it may otherwise impinge the community structure of cyanobacteria, which may leads to unintended alterations in the functioning of estuarine ecosystem. Further studies to elucidate the tolerance limit of cyanobacteria to different heavy metals and the influence of metal pollution on photosynthetic efficiency of cyanobacteria will be appreciated.
The authors thank the Director, CSIR-National Institute of Oceanography, Goa and the Scientist-in-Charge, CSIR-NIO Regional Centre Kochi, for extending all required support. The laboratory facility was extended by MMRF of CSIR-NIO, RC, Kochi, funded by Ministry of Earth Sciences, New Delhi. We thank Prof. Ramola Antao, consultant, Senior Cambridge English Examinations in India, for editing the manuscript. JC is a recipient of WoS-A fellowship of Department of Science and Technology, Govt. of India. AA is thankful to Department of Science and Technology, Govt of India, for the research grant (No. SR/FTP/ES-31/2011). This is NIO contribution No. xxxx.