Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2015) Volume 0, Issue 0
Acorus calamus L. is an emergent aquatic macrophyte that is used in several medicines and food supplements. A. calamus ecotypes with low concentrations of β-asarone can be used for medicinal purposes. We investigated 50 populations of this species, representing the entire natural distribution range in Indian Himalayan Region (IHR), to understand natural variations in β-asarone concentration, ploidy level and genetic divergence. Our results showed that: 1) all four cytotypes (diploid, triploid, tetraploid and hexaploid) of A. calamus exist in natural populations in the IHR; 2) the β-asarone concentration is highly correlated with the ploidy level (diploid and hexaploid populations exhibit low β-asarone concentrations, whereas triploids and tetraploids show high β-asarone concentrations); 3) there is a significant correlation between the β-asarone concentration and its distribution along latitudinal and longitudinal ranges, but not ecological factors (mean annual temperature and mean annual precipitation); and 4) geographic boundaries play an important role in the population divergence of A. calamus. This study provides a foundation for understanding the genetic basis of natural variation in complex traits of A. calamus and for devising conservation strategies for this medically important plant.
Keywords: Acorus calamus; β-asarone; Ploidy; Chloroplast microsatellites; Geographical barriers; Phytochemistry
Acorus calamus L. is a medicinally important herbal and aromatic plant species. This species can grow in varying agroclimatic conditions in India, from tropical to temperate marshes with abundant distribution in Indian Himalayan Region (IHR) [1,2]. A. calamus consists of varying ploidy levels such as diploid (2x=24), triploid (3x=36), tetraploid (4x=48) and hexaploids (6x=72) [1]. The different ploidy levels cannot be morphologically distinguished in the field. This herb is often used as an ingredient in over 77 Indian ayurvedic preparations [3]. It is used as an aphrodisiac, laxative, emetic, expectorant, emmenagogue, and diuretic [4], and it is commonly used in the treatment of infectious diseases [5] and chronic diarrhea, insomnia, melancholia, neurosis, epilepsy, delirium, hysteria, dyspepsia and loss of memory [6,7]. A. calamus is also used as an antitumor and antidepressant agent [8].
Calamus oil is a brownish and yellow-colored essential oil obtained from different parts of A. calamus [9]. This oil contains several phytochemical constituents, but the major (up to 96%) active constituent is the phenolic ether ‘asarone’ [10]. There are three forms of asarone: α, β- and γ- [11]. β-asarone is considered the most characteristic component of the oil [10,11]. Its concentration generally depends on the geographical location, growth stage and ploidy level of the plant [1,12,13]. Although A. calamus has several medicinal, aromatic and industrial properties, the Scientific Committee on Food and Flavoring Substances and the CEFS (Council of European Committee of Experts on Flavoring Substances) has banned its use in food formulations and in other therapeutic preparations [3] due to the carcinogenic and toxic properties of the plants with high concentrations of β-asarone. It has been suggested that only varieties with low β-asarone levels should be used for medicinal purposes (<0.5% dry matter content) [14].
Many previous studies reported a large number of triploid and tetraploid variants with high β-asarone concentrations (i.e., >20% and 75-96%, respectively) in India [12,13,15,16] that cannot be used in medicines or food supplements due to high concentration of β-asarone. In contrast, few studies reported diploid variants with low concentration of β-asarone and no studies reported β-asarone levels in hexaploids [17-19]. Thus far, it is not clear if there are many diploid and hexaploid populations with low β-asarone concentration, which can be used in medicine. Therefore, the identification of low β-asarone containing populations and the conservation of their genetic diversity in their natural habitats are imperative.
The major natural geographic distribution of A. calamus in India occurs in the IHR. The IHR begins in the foothills of southern Siwaliks and extends up to the Tibetan Plateau in the north (Trans-Himalaya), and includes the Indian states of Jammu, Kashmir, Himachal Pradesh, Uttarakhand and Manipur. The IHR makes up a substantial component of the Indian subcontinent and contains diverse flora and fauna. Three of the world’s major rivers cross the entire Himalaya and separate it into distinct regions. Some of the highest mountains on earth are found in this region, and these barriers have influenced the genetic and phytochemical differentiation of many regional species by restricting gene flow and promoting habitat fragmentation. Several studies on various Himalayan species have revealed high levels of population differentiation in both genetic and phytochemical markers [20,21]. These studies have been restricted to the analysis of only intra- and inter-population diversity using genetic and phytochemical markers. However, the roles of geographical barriers and local adaptation on population differentiation have rarely been studied in depth.
To investigate both the genetic and phytochemical diversity of medicinal plants, a combination of DNA fingerprinting and phytochemical profiling is useful [22]. Phytochemical profiling is a comprehensive and quantifiable identification method [23] that establishes a characteristic chemical pattern for a plant species and its extracts [24]. Analytical techniques, such as gas chromatographyflame ionization detection (GC-FID), are widely used for quantitative and qualitative estimations of the secondary metabolites in herbs and for the analysis of the phytochemical variation among different plant populations [24].
In addition to phytochemical profiling, molecular diversity also plays a potential role in the characterization, management and conservation of a plant species [25]. Understanding the pattern of genetic diversity in a geographic context entails the comprehensive knowledge of how landscape features shape population structure [26]. Features such as habitat fragmentation, geographical barriers and local adaptation strongly influence population differentiation at both genetic [27] and phytochemical levels. The fragmentation of natural populations into small isolated patches by geographical barriers, such as mountains, rivers, and valleys, is a major factor [28,29] that may increase population differentiation through random genetic drift, inbreeding and restricted gene flow [30,31]. Local adaptation may occur between populations located in close proximity but adapted to distinct niches. In populations that are adapted to distinct environments, the fitness of immigrants or hybrids may be reduced by natural selection [32], further facilitating genetic divergence [33] and reducing genetic exchange between populations. Our previous population structure study [34] using genomic RAPD and chloroplast SSR markers detected high levels of population differentiation. However, contemporary enhancements in newly developed statistical tools have led to the emergence of methods for evaluation of the landscape and environmental effects on the distribution of genetic and phytochemical variation in natural environments [26].
In this study, we investigated the β-asarone concentration, ploidy level and geographic boundaries in widely distributed natural populations of A. calamus in the IHR. Our intent was to address the following questions: 1) Will the vast collection of natural populations of A. calamus in the IHR provide an opportunity to identify low β-asarone-containing populations? 2) What is the relationship between the ploidy level and β-asarone concentration? 3) Are there significant correlations between the β-asarone concentration and geographic coordinates, as well as ecological factors? 4) What roles have the various landscape features and geographical barriers played in A. calamus population differentiation?
Population sampling
A total of 50 A. calamus L. populations (10 individuals per population, for a total of 500 individuals) were sampled throughout the natural geographic distribution in IHR. The distribution of these populations is illustrated in Figure 1. The name, location, sample size and geographical coordinates of each population are listed in Table 1. No specific permission was required for any locations/activities during the sample collection. The samples did not include any endangered or protected species.
| Population | Origin | Geographical Coordinates | Cytology | Phytochemical Estimation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Code | State | Locality | Latitude (N) | Longitude (E) | Altitude (m) | Chromosome no.; x =12 | Ploidy Level | Yield of essential oil (%) | β-asarone content (%) | β-asarone dry matter content (%) |
| A-44 | Himachal Pradesh | Khaziar lake | 32.32 | 76.03 | 1920 | 2x = 24 | Diploid | 2.50 | 19 | 0.47 |
| A-9 | Himachal Pradesh | Shilly, Solan | 30.92 | 77.12 | 1467 | 3x = 36 | Triploid | 3.87 | 96 | 3.71 |
| A-32 | Himachal Pradesh | Sirmaur, Kotla | 30.43 | 77.17 | 1190 | 3x = 36 | Triploid | 2.60 | 94 | 2.44 |
| A-33 | Himachal Pradesh | Khaltoo, Solan | 30.51 | 77.10 | 939 | 3x = 36 | Triploid | 2.98 | 92 | 2.74 |
| A-42 | Himachal Pradesh | Jatroon, Chamba | 32.25 | 75.59 | 916 | 3x = 36 | Triploid | 4.70 | 94 | 4.41 |
| A-43 | Himachal Pradesh | Mundi, Chamba | 32.21 | 76.00 | 1020 | 3x = 36 | Triploid | 4.15 | 94 | 3.90 |
| A-45 | Himachal Pradesh | Olie, Chamba | 32.32 | 76.07 | 1074 | 3x = 36 | Triploid | 6.25 | 80 | 5.00 |
| A-46 | Himachal Pradesh | Sultanpur, Chamba | 32.33 | 76.60 | 841 | 3x = 36 | Triploid | 1.44 | 96 | 1.38 |
| A-47 | Himachal Pradesh | Sihunta, Chamba | 32.18 | 76.50 | 874 | 3x = 36 | Triploid | 3.04 | 93 | 2.82 |
| A-48 | Himachal Pradesh | Rait, Kangra | 32.11 | 76.12 | 669 | 3x = 36 | Triploid | 5.27 | 96 | 5.05 |
| A-49 | Himachal Pradesh | Kachhred, Kangra | 32.05 | 76.21 | 794 | 3x = 36 | Triploid | 1.99 | 95 | 1.89 |
| A-50 | Himachal Pradesh | Dhanhadi, Una | 31.36 | 76.08 | 430 | 3x = 36 | Triploid | 3.82 | 94 | 3.59 |
| A-14 | Himachal Pradesh | Karsog, Mandi | 31.13 | 76.37 | 754 | 4x = 48 | Tetraploid | 3.54 | 97 | 3.43 |
| A-34 | Uttarakhand | Bhagwati, Almora | 29.51 | 79.18 | 878 | 4x = 48 | Tetraploid | 5.14 | 93 | 4.78 |
| A-35 | Uttarakhand | Chaukhutiya, Almora | 29.52 | 79.21 | 954 | 4x = 48 | Tetraploid | 3.48 | 92 | 3.20 |
| A-36 | Uttarakhand | Aagarchatti, Chamoli | 30.00 | 79.18 | 1321 | 4x = 48 | Tetraploid | 4.76 | 94 | 4.47 |
| A-37 | Uttarakhand | Dwarahat, Almora | 29.46 | 79.25 | 1399 | 4x = 48 | Tetraploid | 3.44 | 92 | 3.16 |
| A-38 | Uttarakhand | Soni, Almora | 29.37 | 79.21 | 1636 | 4x = 48 | Tetraploid | 3.13 | 93 | 2.91 |
| A-39 | Uttarakhand | Khargot, Pittoragarh | 29.33 | 80.11 | 1331 | 4x = 48 | Tetraploid | 4.93 | 93 | 4.58 |
| A-40 | Uttarakhand | Lohaghat, Champawat | 29.24 | 80.50 | 1565 | 4x = 48 | Tetraploid | 1.74 | 95 | 1.65 |
| A-41 | Uttarakhand | Champawat | 29.19 | 80.00 | 1577 | 4x = 48 | Tetraploid | 6.11 | 96 | 5.86 |
| A-53 | Uttarakhand | Jarmola, Uttarkashi | 34.56 | 78.05 | 1352 | 4x = 48 | Tetraploid | 3.42 | 96 | 3.28 |
| A-54 | Uttarakhand | Kateshwar, Almora | 29.37 | 79.51 | 1767 | 4x = 48 | Tetraploid | 2.60 | 94 | 2.44 |
| A-55 | Uttarakhand | Digoli, Almora | 29.38 | 79.43 | 1333 | 4x = 48 | Tetraploid | 3.52 | 96 | 3.37 |
| A-56 | Uttarakhand | Mori, Uttarkashi | 31.01 | 78.03 | 1236 | 4x = 48 | Tetraploid | 3.50 | 96 | 3.36 |
| A-1 | Uttarakhand | FRI Campus, Dehradun | 30.20 | 77.59 | 635 | 4x = 48 | Tetraploid | 4.77 | 96 | 4.57 |
| A-10 | Uttarakhand | Dhanualati | 30.26 | 78.14 | 635 | 4x = 48 | Tetraploid | 3.14 | 96 | 3.01 |
| A-13 | Uttarakhand | Gularghati | 30.18 | 78.01 | 635 | 4x = 48 | Tetraploid | 2.21 | 97 | 2.14 |
| A-16 | Uttarakhand | Almora | 29.36 | 79.30 | 1646 | 4x = 48 | Tetraploid | 3.55 | 96 | 3.40 |
| A-22 | Uttarakhand | Chamba, Chamni | 30.22 | 78.28 | 996 | 4x = 48 | Tetraploid | 3.11 | 95 | 2.95 |
| A-23 | Uttarakhand | Maneri, Uttarkashi | 30.75 | 78.31 | 1537 | 4x = 48 | Tetraploid | 2.22 | 92 | 2.04 |
| A-24 | Uttarakhand | Gangori | 30.73 | 78.45 | 1588 | 4x = 48 | Tetraploid | 2.68 | 92 | 2.46 |
| A-25 | Uttarakhand | Srikot, Srinagar | 30.11 | 78.47 | 545 | 4x = 48 | Tetraploid | 2.03 | 94 | 1.90 |
| A-26 | Uttarakhand | Devaar, Chamoli | 30.19 | 79.11 | 1538 | 4x = 48 | Tetraploid | 2.47 | 91 | 2.24 |
| A-27 | Uttarakhand | Mandal, Chamoli | 30.26 | 79.16 | 1588 | 4x = 48 | Tetraploid | 2.11 | 93 | 1.96 |
| A-29 | Uttarakhand | Khunigad, Uttarkashi | 30.58 | 77.58 | 1135 | 4x = 48 | Tetraploid | 5.01 | 92 | 4.60 |
| A-30 | Uttarakhand | Naugaon, Uttarkashi | 30.47 | 78.07 | 1046 | 4x = 48 | Tetraploid | 4.74 | 93 | 4.40 |
| A-17 | Jammu | Parol, Kathua | 32.17 | 75.32 | 218 | 4x = 48 | Tetraploid | 2.68 | 92 | 2.46 |
| A-18 | Jammu | Jandi, Kathua | 32.27 | 75.16 | 307 | 4x = 48 | Tetraploid | 3.54 | 87 | 3.07 |
| A-19 | Jammu | Khokharchak, Sambha | 32.56 | 75.11 | 384 | 4x = 48 | Tetraploid | 4.74 | 79 | 3.74 |
| A-20 | Jammu | Baghajanha | 32.44 | 74.52 | 305 | 4x = 48 | Tetraploid | 2.03 | 91 | 1.84 |
| A-21 | Jammu | Beril, Kathua | 32.17 | 75.32 | 218 | 4x = 48 | Tetraploid | 0.76 | 82 | 0.62 |
| A-3 | Uttar Pradesh | NBRI, Lucknow | 26.55 | 80.59 | 123 | 4x = 48 | Tetraploid | 3.89 | 96 | 3.73 |
| A-4 | Harayana | Hissar | 29.10 | 75.43 | 211 | 4x = 48 | Tetraploid | 3.20 | 95 | 3.04 |
| A-28 | Arunachal Pradesh | Mao gate, Manipur | 23.83 | 93.03 | 790 | 4x = 48 | Tetraploid | 3.52 | 98 | 3.44 |
| A-51 | Kashmir Valley | Check-i-Kawoosa | 34.07 | 74.38 | 1604 | 6x = 72 | Hexaploid | 2.56 | 18 | 0.46 |
| A-52 | Kashmir Valley | Hokersar | 34.63 | 76.04 | 1570 | 6x = 72 | Hexaploid | 3.30 | 16 | 0.52 |
| A-57 | Kashmir Valley | Gohalkun | 34.14 | 74.30 | 1593 | 6x = 72 | Hexaploid | 2.95 | 20 | 0.59 |
| A-58 | Kashmir Valley | Bajwudar | 34.08 | 74.47 | 1585 | 6x = 72 | Hexaploid | 0.61 | 20 | 0.12 |
| A-59 | Kashmir Valley | Sangam | 33.80 | 75.07 | 1580 | 6x = 72 | Hexaploid | 1.36 | 18 | 0.24 |
Table 1: Geographic locations, ploidy level and concentration of β-asarone for the 50 studied populations of Acorus calamus L.
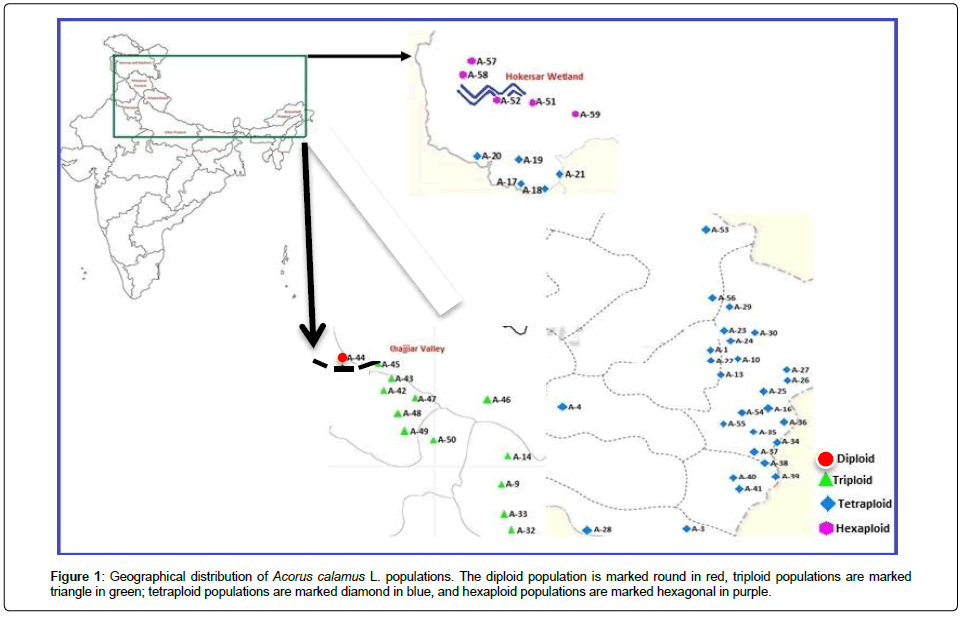
Figure 1: Geographical distribution of Acorus calamus L. populations. The diploid population is marked round in red, triploid populations are marked triangle in green; tetraploid populations are marked diamond in blue, and hexaploid populations are marked hexagonal in purple.
Phytochemical analysis
Rhizomes collected from the fifty different populations were washed thoroughly with tap water and shade dried at room temperature (25 °C) for 3-4 days. The shade-dried rhizomes (100 g) were chopped and hydrodistilled using a Clevenger apparatus (NSW-125 New Delhi, India) for five to six hours. The aqueous distillate obtained was extracted with diethyl ether (2 × 50 ml) in a separating funnel and dried over anhydrous Na2SO4. The solvent (diethyl ether) was removed using a gently heated water bath (NSW-125 New Delhi, India) at 30-35°C until it rendered a yellow colored essential oil with the characteristic sweet calamus odor.
The concentration of β-asarone was estimated using a gas chromatography-flame ionization detector (GC-FID). The GC-FID analysis of the essential oil was carried out on a CHEMITO GC 1000 equipped with a fused silica capillary column (BPX-5, 30 m × 0.22 mm, 0.25-μm film thickness). The chromatographic conditions were as follows: the injector temperature was set at 250°C; nitrogen was used as the carrier gas at a 1ml min-1 flow rate with a 36.4 cm sec-1 linear velocity; the programmed oven temperature was 60°C, using a 10°C ramp to 180°C at 5°C min-1, with a final hold at 220°C for 10 min; the detector temperature was 280°C; and the run time was 60 min. The identification of β-asarone was performed by comparing the percent authentic retention time with the standard. The phytochemical relationship based on the β-asarone concentrations and essential oil yields was analyzed with PCA in MINITAB (Figure 2).
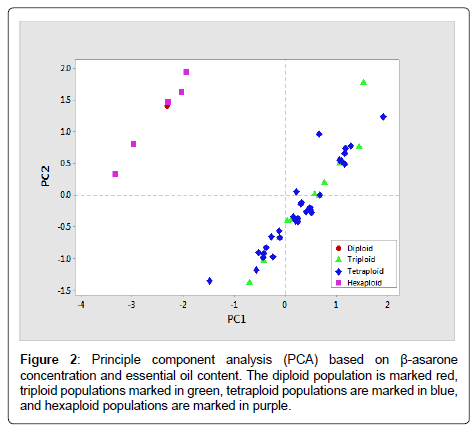
Figure 2: Principle component analysis (PCA) based on β-asarone concentration and essential oil content. The diploid population is marked red, triploid populations marked in green, tetraploid populations are marked in blue, and hexaploid populations are marked in purple.
Ploidy analysis
To determine the ploidy level of each population, young excised roots that were approximately 1-2 cm from the rhizomes were fixed in freshly prepared Carnoy’s fixative (absolute alcohol and acetic acid, 3:1 v/v) for 24 h at room temperature (25°C). After fixation, the roots were transferred to 70% alcohol (v/v) and stored at 4°C. To prepare a squash for the mitotic study, a few roots were placed in a watch glass containing a 9:1 solution of 2% aceto-orcein stain and 1 N HCl. The roots were then warmed gently for 5-7 minutes and left covered for 15 minutes. Subsequently, the roots were placed on a microscope slide and their root tips were removed. The root tips were squashed in a drop of 45% acetic acid by gently pressing with a needle. Finally, the preparation was mounted in a drop of 45% acetic acid, sealed with vaseline and observed under a digital compound microscope (Olympus BX41). A total of 10 cells per individual and 10 individuals per population were examined and analyzed during the metaphase stage to estimate the total number of chromosomes. The micrograph for all the cytotypes was shown in Supplemental Figure S1.
Correlation analyses
To test the correlation between β-asarone and the two main environmental factors (mean annual temperature and mean annual precipitation), the environmental data from WorldClim website (http://www.worldclim.org/ ) for all 50 of the populations were downloaded using the program ArcGIS map 10.2. Due to non-normality of the data, nonparametric spearman correlation analyses were carried out between β-asarone content and environmental factors, as well as between β-asarone and geographical coordinates (i.e., latitude and longitude), and elevation using MINITAB v.14.
Genetic analyses
Haplotype analysis: We reanalyzed the previously published cpSSR data from Ginwal (34). In each individual, the cpDNA alleles for all the loci were combined into a 9-locus chloroplast haplotype (chlorotype). The 9-locus chloroplast haplotype data were analyzed for the total number of haplotypes (nh), haplotypic richness (Rh), mean haplotypic distance between individuals (D2SH) within each population and haplotype diversity (He) using the program HAPLOTYPE analysis v. 1.05 [35].
Genetic diversity and spatial analyses: The genetic polymorphism within populations (HS), [36] total gene diversity (HT), Shannon’s information index (I) [37], Nei’s gene diversity (h), and a measure of population differentiation, FST, were calculated using PopGene v. 1.31 [38] and Arlequin version 3.11 [39]. To partition the variation within populations and between populations, AMOVA was analyzed using GenAlEx v. 6.5 [40]. To analyze the correlation between the geographical distances and Nei’s genetic distances among the 50 populations, the Mantel test was conducted using GenAlEx v. 6.5 [40]. The spatial autocorrelation method has been widely used to assess the association between the genetic relatedness of pairs of individuals and their geographic distance in a smaller geographic scale [41]. Thus, a spatial correlogram was used to evaluate the behavior of the autocorrelation as a function of distance [26]. The autocorrelation coefficient r, bounded by -1 and +1, is related to Moran’s I [42,43]. In this study, autocorrelation analysis was conducted with GenAlEx v. 6.5 [40]. For our data, the maximum geographic distance was divided into ten distance classes of equal size in km.
Identification of genetic boundaries: The detection of genetic discontinuities and the correlation of these discontinuities with landscape features, such as barriers (e.g., mountains, rivers, hills, lakes, and gradient of humidity), are the key to identifying the reasons underlying restricted gene flow among populations. Genetic barriers were identified based on Monmorier’s algorithm, and estimated using Barrier v. 2.2 [43].
Variations in the β-asarone concentrations and ploidy levels
All 50 populations were analyzed for β-asarone concentrations and ploidy level (Table 1). Twenty-four populations collected from different parts of Uttarakhand State were characterized as tetraploids with high β-asarone content (91 to 97%). Out of the thirteen populations collected from different locations in Himachal Pradesh, twelve were characterized as having a high β-asarone content. This included eight triploid populations with 80-95% concentration and four tetraploid populations with a 96-97% concentration. Only one population (A-44) showed a low concentration of β-asarone (19%) and it was found to be diploid (Table 1). All five of the populations from the Jammu region were tetraploid and contained high concentrations of β-asarone (79- 92%), whereas all five of the populations grown in the Kashmir Valley were hexaploids and possessed low concentrations of β-asarone (16- 20%). One population each from Haryana, Uttar Pradesh and Manipur was analyzed and found to be tetraploid containing high β-asarone concentrations (95, 96 and 98%, respectively) (Table 1).
PCA analysis based on the β-asarone concentrations and essential oil yields revealed two major groups. One group includes populations that are either diploids or hexaploids (Figure 2) with lower concentrations of β-asarone (<20%). The other group contains populations that are either triploid or tetraploid (Figure 2) with high β-asarone concentration (79-98%).
Correlation analysis
Correlation analyses were carried out for the β-asarone content, geographical coordinates, elevation, and two ecological factors, mean annual temperature and mean annual precipitation. β-asarone content was found to exhibit a significant positive correlation with longitude (r=0.500, P<0.001 but a negative correlation with altitude (r= -0.511, P<0.001), even after the Bonferroni correction. Interestingly, the β-asarone content did not show any significant correlations with the two ecological factors (Table 2). There is no significant correlation between β-asarone and elevation (r= -0.264, P=0.064). The nonparametric t-test of elevations between the high β-asarone populations and low β-asarone showed significant difference (P=0.001).
| Variables | Latitude | Longitude | Altitude | MAT | MAP | β-Asarone |
|---|---|---|---|---|---|---|
| Latitude | ||||||
| Longitude | -0.846 (0.000) | |||||
| Altitude | 0.016 (0.913) | 0.215 (0.133) | ||||
| MAT | -0.201 (0.162) | -0.125 (0.386) | -0.656 (0.000) | |||
| MAP | -0.356 (0.011) | 0.397 (0.004) | 0.169 (0.241) | -0.004(0.977) | ||
| β-Asarone | -0.511 (0.000) | 0.500 (0.000) | -0.264 (0.064) | 0.190 (0.187) | 0.202 (0.160) |
Table 2: Correlation coefficients (r) between geographical coordinates, β-asarone content, and ecological factors. The numbers marked bold indicate significant correlation.
Genetic diversity and differentiation
A. calamus L. shows high level of diversity at the species level: Nei’s genetic diversity, HT= 0.530 ± 0.005, and the mean haplotypic diversity, He =0.610. This species also shows a high level of population differentiation: I=0.864 and PhiPT (an analog of FST) = 0.73, with a low level of inter-population gene flow (Nm =0.184). Within populations, the mean Nei’s gene diversity (h) and Shannon’s information index (I) were found to be 0.137 and 0.149, respectively (Table S1). Nei’s gene diversity (h) was found to be highest in population A-45 (0.435) and lowest in populations A-4, A-9, A-10, A-16, A-23, A-24, A-25, A-27, A-29 and A-38 (0.001). Shannon’s information index followed a similar trend (Table S1). A total of 137 chloroplast haplotypes were found, with the maximum number in populations A-30, A-32, A-42, and A-48. The mean haplotype diversity among the populations was 0.610, and the haplotype diversity within populations ranged from 0.000 to 0.889 (Table S2).
The results from the AMOVA analysis show that amongpopulation divergence contributes the most to the total variation (73%), while within population variation contributes a small portion (27%). The Mantel test did not detect a significant correlation between genetic distance and geographic distance (r=0.017, P=0.32). Genetic substructure was only detected in the distance class 6640 km by spatial autocorrelation analysis (Figure 3). As per Monomier’s algorithm (44), a total of nine genetic barriers (a, b, c, d, e, f, g, h, and i) were identified, which separate populations into isolated patches. The thick red lines in Figure 4 represent barriers among the Uttarakhand, Himachal Pradesh and Jammu & Kashmir. The genetic barrier separates population A-44 (diploid) from all of the other populations in Himachal Pradesh (Figure 4a). Genetic boundaries b, c, d, f, and g separate Kashmir populations (i.e., A-51, A-52, A-57, A-58 and A-59, hexaploid) from the other populations (Figure 4b). Even within Kashmir regions, genetic boundaries (b, c, f and g) were detected (Figure 4b).
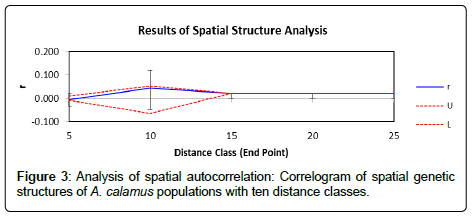
Figure 3: Analysis of spatial autocorrelation: Correlogram of spatial genetic structures of A. calamus populations with ten distance classes.
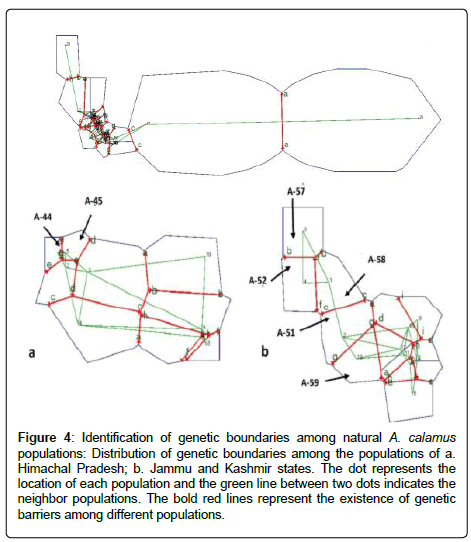
Figure 4: Identification of genetic boundaries among natural A. calamus populations: Distribution of genetic boundaries among the populations of a. Himachal Pradesh; b. Jammu and Kashmir states. The dot represents the location of each population and the green line between two dots indicates the neighbor populations. The bold red lines represent the existence of genetic barriers among different populations.
Natural A. calamus populations containing all four cytotypes, including diploids and hexaploids, exist in India
An A. calamus plant can be used for medicinal purposes only if it exhibits a low level of β-asarone and it is carcinogenic with a high level of β-asarone. Two strategies are used to obtain low levels of β-asarone. One is to convert high levels of β-asarone to other forms of asarone, such as the α- and γ- forms [44]. However, this process is time consuming, labor-intensive and expensive. The other strategy is to identify low β-asarone-containing populations in their natural habitats. Thus far, however, researchers have only found triploids and tetraploids that contain high concentrations of the carcinogenic β-asarone compound in India [45]. Few studies reported diploids in India but with small sample size and geographically limited to the southern India where usually considered no natural populations [46]. One study reported hexaploids in the Kashmir region of India but did not measure β-asarone content [18]. It is therefore not surprising that there is a common misconception that only triploids and tetraploids containing high β-asarone levels exist in the natural A. calamus populations in IHR. For the first time, we sampled 50 populations across the species’ natural distribution range in India (IHR) and, besides triploid and tetraploid plants, we also identified diploid and hexaploid individuals (Table1 and Figure1).
Among the 50 sampled populations, the majority of the populations were triploid and tetraploid; only one diploid and five hexaploid populations were identified. The latter are mainly found in small isolated patches (Figure 4) and probably have experienced local adaptation. This is easy to imagine given the geographic barriers (valleys, rivers, mountains) in the natural range of this species. Another possibility is that the diploid/hexaploid populations might have been collected aggressively by local people and thus reduced in frequency. If so, it is important to identify these populations and to protect them from over-collection.
The β-asarone concentration has high correlation with the ploidy level
We found a clear relationship between β-asarone and ploidy level: diploids and hexaploids have lower β-asarone concentrations, while triploids and tetraploids have higher β-asarone concentrations (Table 1). Several previous studies with samples from other continents have reported a similar relationship between β-asarone and ploidy level (Table S3) [13,47]. Even though the small number of diploid and hexaploid populations found in our 50 sampled populations limits the statistical power, this correlation is convincing due to the compatible results found in other studies [Supplemental Table S3]. Additional studies with more samples from diverse habitats should further corroborate this general relationship.
Different levels of β-asarone concentration in diverse locations might be affected by stress from competition with surrounding vegetation. It is possible that the low concentration of β-asarone in hexaploids from the Kashmir valley might be because stress in this region is relatively lower than in the other regions. If so, however, this would not explain the low content in the diploid population that grows along with triploid populations in the Himachal Pradesh region with similar competing species. Moreover, diploid populations from different continents with different competition stresses all show lower levels of β–asarone content [11,12,14,19,47-50]. Thus, it is unlikely that competition stress is a viable explanation for the differences in the concentration of β-asarone among the populations.
Different levels of β–asarone concentration among diverse populations can also be attributed to seasonal variation. For example, Venskutonis et al. [48] and Ogra et al. [1] reported that β–asarone concentrations depend on the different plant tissues and seasonal variations. However, the β-asarone concentration within the diploid cytotypes was always lower with the range of 0-5% compared to the triploids (60-80%) and tetraploids (80-90%) [49] no matter which tissues were analyzed and which season the materials were collected.
Correlation between β-asarone content with geographic coordinates and ecological factors
We found that the β-asarone concentration was correlated with both latitude and longitude (Table 2). Low β-asarone-containing populations can be found at higher longitudes and lower latitudes. This can be explained by the fact that in India, A. calamus grows in varying agroclimatic conditions, and its oil composition and β-asarone content were found to be greatly affected by its geographic location [12,13,50]. There was no significant correlation between β-asarone and elevation. This might be due to the smaller number of samples in low β-asarone populations. However, after the nonparametric t-test in elevations between two groups (low vs high β-asarone content), significant difference between the two groups was found (P=0.001). The low β-asarone content populations can be found at high elevation, while high β-asarone content populations are typically found at low elevation. This pattern was also found in Ogra et al. [1].
Environmental conditions also have been suggested as having substantial effects on the concentration of the β-asarone compound in the calamus oil [1]. To test this hypothesis, association analyses were conducted between β-asarone content and two main environmental factors, mean annual temperature and mean annual precipitation. However, no significant correlation was found between β-asarone content and these two major ecological factors. The significant correlation between β–asarone and geographical coordinates, rather than with specific ecological factors, suggests that low concentration of β–asarone might be affected by other environmental factors or a complex suite of environmental factors.
Spatial genetic structure
The cpSSR analysis revealed high population differentiation (I=0.864; PhiPT=0.73). However, the Mantel test (r=0.02; P=0.32) did not show a significant correlation between geographic distances and genetic distance among all sampled populations. This indicates that the geographic distances are not key factors that have restricted gene flow among the populations as would be indicated by the presence of isolation by distance (26). Instead, geographically close populations presumably show high differentiation due to adaptation and limited gene flow brought about by geographic barriers. For example, A-44 (diploid) and triploid populations are geographically very close (Figure 1) but these populations show pronounced genetic and phytochemical differences (Table 1).
Spatial autocorrelation analysis did not detect substructure in most distance classes (Figure 3), probably because of the complex landscape and geographic features in this region. A number of studies in the past two decades have suggested that climate changes and geographic features are key factors influencing formation of glacial lakes, deep valleys, and mountainous environments in this region [51]. The continuous emergence of these geographic boundaries is the primary reason underlying habitat fragmentation and restricted gene flow in a number of species [51]. Our investigation of A. calamus populations in the IHR helps us understand whether high geographic and landscape variability has influenced the genetic and phytochemical diversity in this species. The identification of several genetic boundaries suggested that the high genetic divergence among A. calamus populations is likely attributed to the restricted gene flow due to geographic boundaries that separated populations into small isolated patches [31,52-56]. For example, genetic barrier g separates the diploid population (A-44) from the other neighbor populations (Figure 4a), which might be caused by the geographic barrier, Khajjiar Valley (Figure 1). Also, genetic barriers (b, c, f and g) separate two hexaploid populations (A-57 and A-58) from the other hexaploid ones (A-51, A-52 and A-59) (Figure 4b). The geographic barrier Hokersar Wetland might have played important role in the genetic differentiation (Figure 1).
Conservation implications
A. calamus has been categorized as a medicinal plant with immense potential to cure various diseases. However, the medicinally important cytotypes of this species are threatened in India [57]. The wild populations of diploids and hexaploids are sharply declining in India. Therefore, viable conservation strategies are needed to preserve the dwindling genetic diversity of this species. The results indicated that the phytochemical and genetic variability observed in A. calamus primarily reside at the inter-population level, with low intra-population variation due to restricted gene flow, drift, and selection. Based on these results, it is necessary to protect populations occurring in as many distinctive ecological sites as possible, with a special emphasis on identifying low β-asarone-containing populations. These populations deserve the attention of genetic conservation programs for mass-scale production. The conservation of various genotypes and the mass-scale multiplication of identified populations will help to protect the various genotypes of the species and also create a greater availability of low β-asarone-containing varieties/populations for industrial utilization. This availability will also enhance the medicinal and economical value of these highly valued, but vulnerable, medicinally important cytotypes in India.
We thank to Dr. Larry Leamy, Daniella Freese and Cameryn Bevill for helpful comments and proof reading.