Journal of Biomedical Engineering and Medical Devices
Open Access
ISSN: 2475-7586
ISSN: 2475-7586
Research Article - (2016) Volume 1, Issue 1
Keywords: Saccades; Double-step; Visual and auditory-visual bisensory stimuli; Oculomotor plant; Neural input; Parallel programming
Saccades are known as rapid eye movements made in order to locate a new target. Sensory inputs, including visual, auditory, and auditoryvisual combined stimuli, that can trigger voluntary saccades. Characteristics of such goal-oriented one-step (OS) saccades have been investigated extensively in previous studies [1-8]. Generally, saccades are extremely variable, but the variability is well coordinated for saccades of the same size. A typical visually-guided one-step (V-OS) saccade usually exhibits a peak velocity from 100 deg/s up to 500 deg/s, duration of 30-100 ms and a latency of 100-300 ms. Auditory (A-OS) saccades show lower peak velocity and longer duration, while bisensory (AV-OS) saccades have significant reduced latency [1].
Saccades operate in an open-loop mode and they are conjugate and ballistic. When a target is presented, the central nervous system determines whether to make a saccade. During latent period, the distance the eyeball needs to move is calculated, and then retinal or auditory error is transformed into transient muscle activity. To investigate how a saccade is modified by a second stimulus that is applied before the eye starts to move, the double-step paradigm has been developed by presenting subjects with two visual targets along a horizontal plane with a varying inter-stimulus interval (S1 in Figure 1) [9]. Our previous study indicates that the programming of a saccade to the second peripheral visual target can be initiated prior to the execution of a preceding saccade, but at the same time, the generation of the saccade to the first peripheral visual target may not be delayed when a second target is applied. These results are either a single-step (SS) response to the second target location directly or a double-step (DS) response [10]. Some other researchers have also discussed such “parallel programing” mechanism, which the visual information can be acquired continuously to modify saccade amplitude until about 70-100 ms prior to eye movement onset [11-17]. The analysis of response localization and latency under double-step conditions has also been presented to study saccade sequence preplanning [14,18-20].
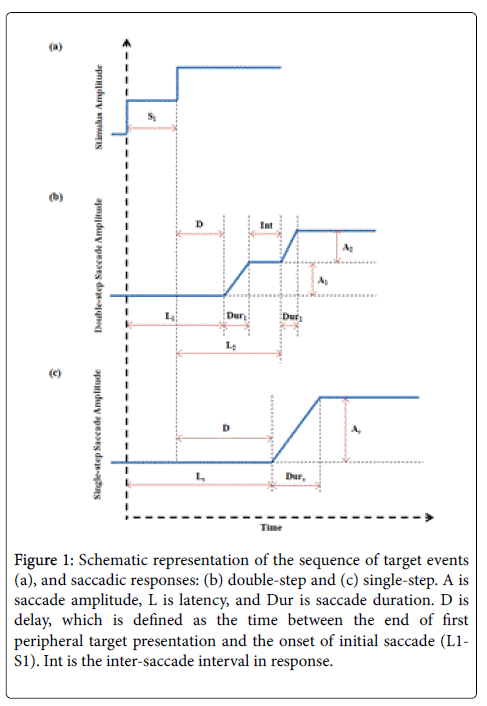
Figure 1: Schematic representation of the sequence of target events (a), and saccadic responses: (b) double-step and (c) single-step. A is saccade amplitude, L is latency, and Dur is saccade duration. D is delay, which is defined as the time between the end of first peripheral target presentation and the onset of initial saccade (L1- S1). Int is the inter-saccade interval in response.
The neural input to the oculomotor plant is responsible for movement of the eyes so that images are focused on the fovea. The work presented here is based on Enderle and his coworker’s whole muscle model [2,3] and muscle fiber muscle model [21], while the inputs to the muscle are agonist and antagonist active-state tensions that are described by pulse-slide-step waveforms with a post inhibitory rebound burst (PIRB) based on a time-optimal controller [2,3,22-25]. The neural input estimations provided in our previous studies support the responsive neuron populations in superior colliculus under different stimulus conditions [1], and indicate the double-step visual stimulus may affect the synchrony of neuron firing [10]. This paper is a follow-up of our visual double-step saccades (V-DS) study by adding the comparisons of saccade characteristics and neural control to auditory-visual bisensory double-step saccades (AV-DS) [10], further expands the saccade system investigation [1,10].
Detailed methodology has been presented in previous publications [1,10].
Subjects
Four healthy human volunteers participated the study. All subjects demonstrated normal or corrected-to-normal visions, and none disclosed any history of visual, auditory or vestibular disorders.
The subjects were trained on tasks of goal-oriented saccade experiments (one-step) under different stimulus conditions before performing the double-step saccades. The subjects were involved in both V-DS and AV-DS experiments.
Apparatus and experiment design
The visual targets consisted of white solid dots (4 mm diameter) on a gray background and were displayed on the screen which was 830 mm in front of the subjects. Seven target positions were presented at angles of 0, 5, 10 and 20 from the center to left or right in the horizontal plane. The auditory targets were tone signals at 1 kHz frequency with an average intensity around 60 dB SPL. To coordinate the sound stimuli locations with visual stimuli, 3D positional audio was implemented using H3D Binaural Spatializer plug-in (by Longcat Audio Technologies) inserted in audio editing software (Adobe Audition CS5.5), and the sounds were heard by a stereo noise-reduced headphone. Subjects were seated with head stabilized by a high speed eye tracking device (1250 Hz sample rate, by SensoMotoric Instruments (SMI)). Either visual stimuli (V-DS) or auditory-visual combined (synchronous) (AV-DS) targets were provided, and the subjects were instructed to maintain their eyes on the target by moving their eyes as fast as possible.
The experiments were computer-controlled using custom written software, while data were collected using the eye tracker implemented in the iView XTM system by SMI. All the tests were performed in an independent, quiet room with normal illumination. In either V-DS or AV-DS experiments, there were three test sessions with the first peripheral target’s duration (S1, shown in Figure 1 set as 70, 140 and 210 ms. The stimulus pattern followed a “stair case”, that is the target stepped twice centrifugally (toward the periphery) in the same direction. Each session included 32 trials: 16 0-5-10 double-step displacements and 16 010 20 double-step displacements, to right or left. The duration of any center or second peripheral stimulus in a trial was random from 3000 to 5000 ms.
Data processing and analysis
Raw data contained pupil and gaze information were generated automatically by the iView X™ system. Gaze data from the left eye were recorded in pixels, and then converted to degrees. The converted eye position data were first plotted for quality and response pattern examination. If two steps were obtained (Figure 1), they were separated for further analysis.
Acceptable saccades data were run in a FORTRAN program which was used to compute parameter estimates for the models of horizontal saccadic eye movements [2,3]. Final estimates of model parameters were found using a system identification technique. Saccade characteristics and neural inputs were also estimated, and post-saccade phenomena were analyzed.
378 trials of visual saccade data and 415 trials of auditory-visual saccade data that contained either double-step or single-step responses were successfully analyzed. Based on the data quality and initial condition requirements, approximately 12% of the collected data were discarded. Two subjects’ data are presented in this paper for discussion. Despite the variation between subjects, the results obtained were highly consistent with each other.
Saccade amplitude and latency
Single-step response occurrence rate is shown in Table 1. Under either visual or auditory-visual stimulus condition, subjects directly follow the second target without landing on the first peripheral stimulus (shown as a single-step response) when inter-stimulus interval S1 is small (70 ms), and a slight higher chance occurs under AV condition. More double-step responses are obtained with increasing S1. In contrast to that with small S1, AV condition facilities double-step response when S1 is 140 or 210 ms, providing lower single-step response occurrence rate. When compare between different trial patterns, single-step responses have a higher chance to appear in 0510 trials.
| 0°-5°-10° Pattern | 0°-10°-20° Pattern | |||
|---|---|---|---|---|
| V | AV | V | AV | |
| S1=70 ms | 54.70% | 56.30% | 25.00% | 34.80% |
| S1=140 ms | 12.50% | 8.90% | 4.70% | 4.10% |
| S1=210 ms | 4.70% | 0% | 4.70% | 0% |
Table 1: Single-step response occurrence rate.
The initial/primary saccade amplitude (A1 and As) verses delay D (Figure 1) under two different stimulus conditions is shown in Figure 2. V and AV responses exhibit similar patterns while the delay in AV saccades can be as small as -100 ms.
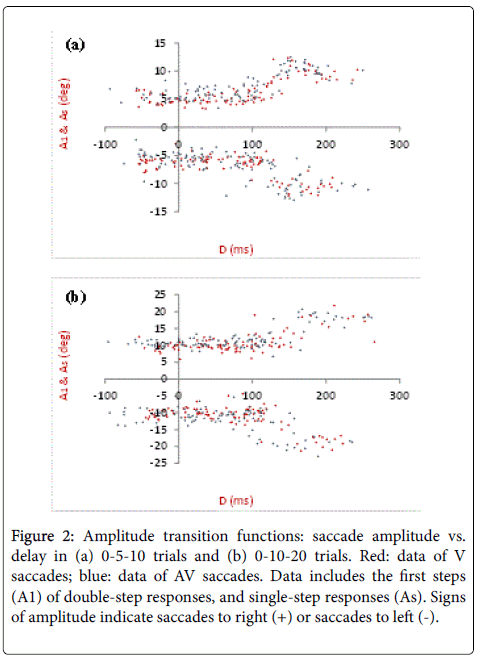
Figure 2: Amplitude transition functions: saccade amplitude vs. delay in (a) 0-5-10 trials and (b) 0-10-20 trials. Red: data of V saccades; blue: data of AV saccades. Data includes the first steps (A1) of double-step responses, and single-step responses (As). Signs of amplitude indicate saccades to right (+) or saccades to left (-).
One subject’s second step saccade latency L2 (defined in Figure 1) is plotted against delay as shown in Figure 3. Average L2 in response to AV stimulus is slight lower than that to V stimulus. When delays fall in the range from 40 to 150 ms, L2 shows a linear increase, where the approximate slope is 1. In general, the latency data shows smaller variance under AV condition.
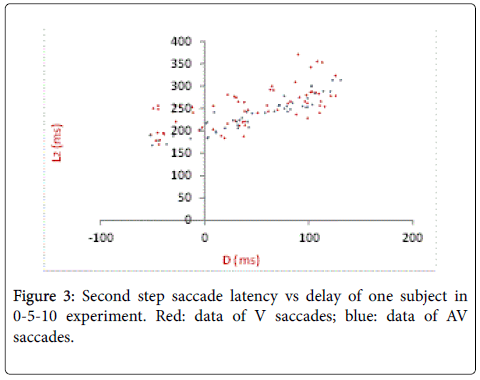
Figure 3: Second step saccade latency vs delay of one subject in 0-5-10 experiment. Red: data of V saccades; blue: data of AV saccades.
Agonist pulse magnitude
Parameter estimations were calculated by the system identification technique (for details, refer to Ref. [2,3]) for a double-step response. Figure 4 shows the estimated agonist and antagonist neural inputs and active-state tensions of a V-DS with the first and second steps analyzed separately. The model predictions match the data very well.
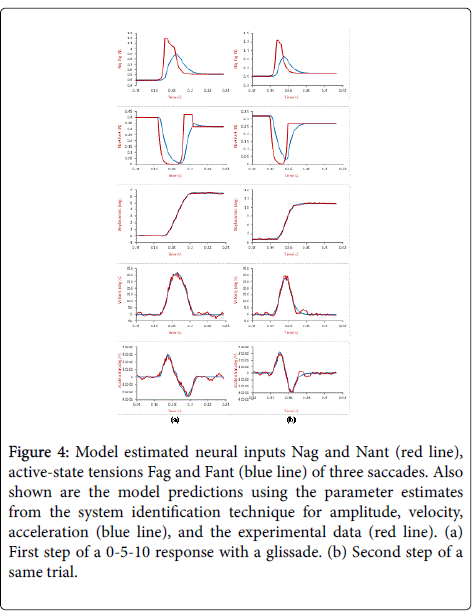
Figure 4: Model estimated neural inputs Nag and Nant (red line), active-state tensions Fag and Fant (blue line) of three saccades. Also shown are the model predictions using the parameter estimates from the system identification technique for amplitude, velocity, acceleration (blue line), and the experimental data (red line). (a) First step of a 0-5-10 response with a glissade. (b) Second step of a same trial.
Agonist pulse magnitude is defined as the peak magnitude of the agonist neural input, as shown in Figure 4 first row (red). The results of the model estimation are shown in Figures 5 and 6, including the pulse magnitude of single-step and/or the two separate steps in double-step responses for two subjects. In Figure 5, the results of a same step (first or second) are compared under different stimulus conditions. For this subject, V-DS shows a slight higher pulse magnitude for saccades around 10 degrees, this is more obvious in the first steps (Figure 5). No significant difference is observed between the V-DS and AV-DS for small saccades. In Figure 6, the pulse magnitude of two steps in AV-DS experiments are plotted together for comparison (for V-DS results [10]). In Figure 6, saccades have the pulse magnitude of secondary saccades slightly larger than that of the primary/initial saccades of the same size, which are less than 7 degrees.
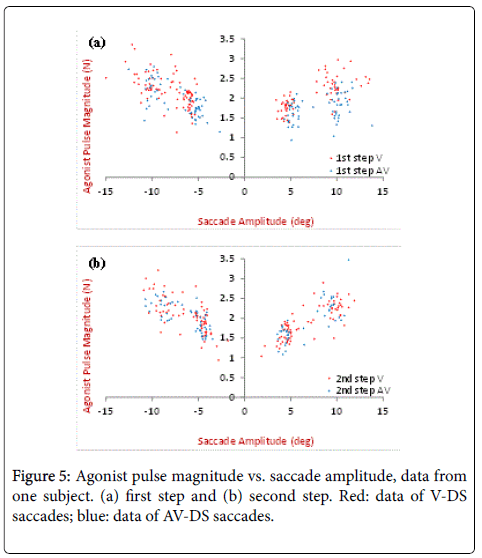
Figure 5: Agonist pulse magnitude vs. saccade amplitude, data from one subject. (a) first step and (b) second step. Red: data of V-DS saccades; blue: data of AV-DS saccades.
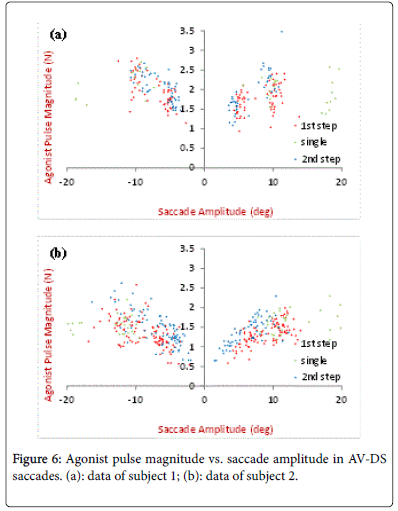
Figure 6: Agonist pulse magnitude vs. saccade amplitude in AV-DS saccades. (a): data of subject 1; (b): data of subject 2.
Saccade characteristics
The aim of this study is to compare the saccade characteristics and neural input properties between goal-oriented saccades respond to one-step or double-step visual and auditory-visual targets. With such purpose, our experiment design focuses on horizontal saccades with smaller stimulus steps (5 and 10) appear centrifugally, while some other double-step studies in literatures use large amplitude targets (12, 15, 45, etc.) in horizontal plane [11,12,14], or in 2-dimentional coordinates [13,17]. Saccade amplitude transition function (ATF) was first introduced by Becker and Jürgens [11], where the primary/initial response amplitude is plotted verses delay D. Under both visual and bisensory conditions, double-step responses are obtained when the delay is smaller than 100 ms, while single-step responses occur when the delay falls between 170 and 250 ms. Since the delay is defined as the first/primary saccade latency minus the duration of the first peripheral target, the smaller D toward negative 100 ms observed in AV-DS indicates lower AV saccade latency, which is consistent with our previous studies [1]. For a one-step target triggered saccade, average latency under V and AV conditions are reported as approximately 190 and 150 ms, respectively [1]. For one of our subjects, the average saccade latency of single-step responses (Ls in Figure 1) in 0-5-10 experiments (meaning As˜10) is 252 ms under V condition and 224 ms under AV condition, when S1 is 70 ms. This means the retinal error is immediately updated when a second target is applied; the programming of the preceding saccade ends and the programming of saccade to the new target starts at once resulting is a single-step response with relative long delay. As the inter-stimulus interval increases, the preceding saccade programming cannot be modified, but the programming of two sequential saccades may be overlapped.
Neural input estimation
The time-optimal controller implemented in our saccade model states that the overall neural input increases as the amplitude of saccades increases up to 7 due to the increasing number of firing neurons, with all neurons firing maximally. In large saccades, all neurons fire maximally, which results in a relatively constant pulse magnitude [2,3,21].
When compared the first steps under different stimulus conditions, visual response shows a larger agonist pulse magnitude for saccades around 10 degrees (Figure 5). This further supports the results in our previous study [1], indicating a smaller population of neurons fire in the superior colliculus during AV-saccades than V-saccades, while all responsible neurons fire maximally.
When we looked at the AV-DS only (Figure 6), for both subjects, the pulse magnitude appears similar for primary/initial and secondary saccades for large saccade size, same results have been shown in V-DS as well [10]. This is explained that only the retinal error determines the active neuron population. Of note for subject 2, the pulse magnitude of small first step response is slightly lower than that of small second step response (Figure 6). Since smaller saccades have fewer agonist neurons firing than larger saccades, the firing synchrony of individual neurons has a greater impact on the overall neural input for small saccades [2,3,21]. Any lack of synchrony causes the overall agonist input to be smaller. This indicates that the double-step stimulus may affect the synchrony of neuron firing. It can be obtained in both V-DS and AVDS [10].
Auditory double-step saccades
Auditory double-step saccade experiments were also conducted with similar design. Auditory targets were displayed alone without any visual cues. Due to the difficulty in accurately locating such centrifugally sound steps, the results showed great variability and they are not presented here. Generally, auditory double-steps saccades exhibited a much longer latent period and inter-saccade interval. Moreover, the saccade amplitude of the first steps was larger than the desired value, meaning the eye landed in the location between the first and second peripheral sound target. Similar results are shown in AVDS responses (Figure 6), where the average saccade amplitude of the first steps are slight higher than that of the second steps.
The work presented in this paper provides a more comprehensive analysis of horizontal human saccadic eye movements. Saccades responded to auditory and/or visual stimulus under double-step paradigm were recorded by a high sample rate eye tracking system, and then processed to produce parameter estimates using system identification technique for a 3rd-order linear horizontal saccade model. Saccade characteristics, including amplitude and latency, indicated that visual information can be acquired continuously to modify saccade amplitude until about 70-100 ms prior to eye movement onset, and the CNS appears able to overlap some presaccadic planning. The estimated neural inputs supported the responsive neuron populations in the superior colliculus under different stimulus conditions, and indicated the double-step visual or auditory-visual stimulus may affect the synchrony of neuron firing.