Journal of Cell Science & Therapy
Open Access
ISSN: 2157-7013
ISSN: 2157-7013
Review Article - (2015) Volume 6, Issue 5
METCAM/MUC18, an integral membrane cell adhesion molecule (CAM) in the Ig-like gene superfamily, is capable of performing typical functions of CAMs, such as cell-cell and cell-extracellular interactions, crosstalk with intracellular signaling pathways, and modulating social behaviors. METCAM/MUC18 is not expressed in >90% of the epithelial cells of normal prostate, or in 100% of benign prostatic hyperplasia (BPH), but is expressed in >80% of prostatic intracellular neoplasia (PIN), high grade prostate cancers, and metastatic lesions. Its expression is also correlated with the malignant progression of mouse prostate adenocarcinoma in a transgenic model, TRAMP. Overexpression of human METCAM/MUC18 increases epithelial-to-mesenchymal transition (in vitro motility and in vitro invasiveness) of prostate cancer cells and in vivo tumorigenesis and metastasis to multiple organs after orthotopic injection of human prostatic cancer LNCaP cells in male nude mice. From our preliminary studies, it appears to regulate these processes via increasing proliferation, up-regulating the AKT-signaling pathway, increasing aerobic glycolysis, and augmenting angiogenesis of prostate cancer cells, but has no effect on apoptosis. Furthermore, soluble METCAM/MUC18 could block angiogenesis of LNCaP tumors and specific shRNAs in a lentivirus vector block tumorigenesis of DU145 cells in an athymic nude mouse model. Taken together, METCAM/MUC18 may be a useful novel biomarker for early diagnosis of the malignant potential of prostate cancer, but also a metastatic progression gene to drive the malignant progression of prostate cancer in a pre-clinic mouse model. METCAM/ MUC18-specific siRNAs and its derived oligo-peptides may be useful as therapeutic agents to block the malignant progression of the cancer.
Keywords: METCAM/MUC18; Cell adhesion molecule; Immunoglobulin-like gene super family; biomarker and driver; siRNA blocking; Prostate cancer malignant progression; Animal models
Prostate cancer is the most commonly diagnosed cancer in American males and the second most common cause of cancer death among males in USA [1]. The majority of histologically localized prostate cancers remains subclinical and never requires treatment. However in some patients, prostate cancers rapidly metastasize and are highly fatal. The high lethality of the cancer in these patients is due to a rapid spread of the cancer cells outside of the gland and killing the victims within a year of initial diagnosis. Unfortunately, it is still impossible to predict when and which localized tumors will become cancer [2]. In addition, an accurate clinical prognosis of the carcinoma in any individual case is quite difficult because 85% of prostatic carcinomas are multifocal, the cancer seen in biopsies may not accurately reflect the entirely biological potential of the tumor [3]. Also, the standard diagnosis for the metastatic potential of prostate cancer, an elevated serum PSA level, does not always predict a pathological stage or the presence of a metastatic prostate cancer, because it is a prostate-specific marker, but not a prostate cancer-specific marker [4]. Many potential diagnostic markers for prostate cancer progression have been developed, however, few have been validated in large scale clinical trials [5]; most of them cannot accurately distinguish between indolent and aggressive cancers [6]. As such, there is still an urgent need to search for a better marker for the early detection of the metastatic potential of prostate carcinomas. Ideally, one would like to find diagnostic molecular markers that are also key players in converting an indolent cancer into an aggressive (metastatic) cancer. After being identified, these markers can also be used to design an effective therapy.
The complexity of tumor metastasis suggests that many genes are directly or indirectly involved in this process. Many metastatic genes and metastatic suppressor genes have been identified, suggesting that there are many alternative metastasis pathways [7]. Furthermore, the genome of metastatic tumor cells has greater instability than nonmetastatic tumor cells; thus metastatic tumor cells acquire multiple intrinsic genetic and epigenetic changes that are attributable to their metastatic phenotype [8], including genes encoding cell adhesion molecules (CAMs), such as E-cadherin [9], CD44 [10], integrin αVβ3 [11], integrins α2 & α6 [12], and METCAM/MUC18 [13-15]. Since cell-adhesion molecules govern the social behaviors of cells, the altered expressions of CAM genes are doomed to affect the motility and invasiveness of many tumor cells in vitro and their metastasis in vivo [16,17]. As documented in many carcinomas, a complex participation of many over- or under-expressed CAMs actually contributes to the metastatic potential of a tumor cell [18]. Furthermore, altered expressions of cell adhesion molecules (CAMs) in metastatic cells may increase their ability to interact with the extra-cellular matrix and establish secondary growth in a favorable soil (a favorable growth microenvironment or niche in the distant organs that had been conditioned by the tumor cells in the micrometastases) [19]. Studies of the altered expression of CAMs in prostate cancers have been limited to E-cadherin [9], CD44 [10], some integrins [11,20], and CEA-CAM 1 [21]. From our studies on the effect of the expression of cell adhesion molecules on the prostate cancer metastasis, we have found that huMETCAM/MUC18 is a bona fide metastatic gene and it very likely plays a role in converting an indolent prostate cancer to an aggressive one [13-15,22-27].
An Ig-like cell adhesion molecule, human METCAM/MUC18, as a biomarker for the progression of human prostate cancer
Human METCAM/MUC18 (huMETCAM/MUC18) is an integral membrane CAM in the immunoglobulin (Ig) gene superfamily [15,28]. The name “MUC18” is often mistaken as a new mucin member. To avoid this unnecessary confusion and to reflect more precisely its biochemical properties and biological functions, we have recently changed its name to “METCAM” (metastasis cell adhesion molecule) [15]. The name “MELCAM”, which was coined to overly emphasize its importance in causing metastasis of melanoma cells [29-31], is not used because the same group has also proven its secondary importance in melanoma metastasis [30]. Other names, like CD146 [31], A32 [32], S-endo1 [33], are also discarded because they did not reflect its full biological functions and biochemical properties. MUC18 was retained here in addition to METCAM for the purpose of commemorating its original finding [28]. Human METCAM/MUC18 (huMETCAM/ MUC18) has 646 amino acids that include an N-terminal extracellular domain of 558 amino acids, which has a 28 amino acids characteristic of a signal peptide sequence at its N-terminus, a transmembrane domain of 24 amino acids (amino acid #559-583), and a cytoplasmic domain of 64 amino acids at the C-terminus. HuMETCAM/MUC18 has eight putative N-glycosylation sites (Asn-X-Ser/Thr) and is heavily glycosylated and sialylated resulting in an apparent molecular weight of 113,000-150,000. The extracellular domain of the protein comprises five immunoglobulin-like domains, V-V-C2-C2-C2 [15,28] and one X domain [15], which has four potential N-glycosylation sites. Its structure is depicted in Figure 1.
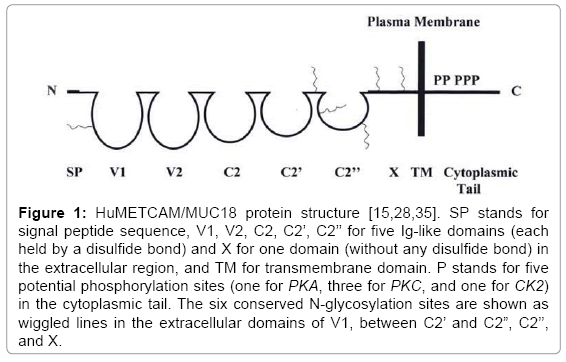
Figure 1: HuMETCAM/MUC18 protein structure [15,28,35]. SP stands for signal peptide sequence, V1, V2, C2, C2’, C2’’ for five Ig-like domains (each held by a disulfide bond) and X for one domain (without any disulfide bond) in the extracellular region, and TM for transmembrane domain. P stands for five potential phosphorylation sites (one for PKA, three for PKC, and one for CK2) in the cytoplasmic tail. The six conserved N-glycosylation sites are shown as wiggled lines in the extracellular domains of V1, between C2’ and C2”, C2’’, and X.
There is a limited literature concerning the contribution of these ecto-domains to the interaction of huMETCAM/MUC18 with its cognate heterophilic ligands, because they have not been identified [15]. The cytoplasmic tail contains peptide sequences potentially to be phosphorylated by PKC, PKA, and casein kinase II [15,28], which implicate its important roles in interactions with intra-cellular signaling pathways. Thus, we predict from the protein structure that METCAM/ MUC18 is necessary for the cells to interact with other cells and extracellular matrix, and with cytoskeleton, and cross talks with growth factor receptors and intra-cellular signaling pathways [15,18,34].
HuMETCAM/MUC18 is only expressed in a limited number of normal tissues/cells, such as endothelial cells, hair follicular cells, smooth muscle cells, basal cells in bronchial epithelium, the cerebellum, some activated T cells, intermediate trophoblasts [35], normal breast epithelium [36], normal ovarian epithelium [37], and nasopharyngeal epithelium [38]. HuMETCAM/MUC18 was first implied to play a role in the malignant progression of melanoma, because it is frequently and highly expressed in malignant melanoma, but less frequently and weakly expressed in dysplastic nevi and not expressed in melanocyte [28]. Its positive role in the progression of melanoma has been substantiated by the results of animal studies when the experimental metastasis test (i.v. injection of cells into the tail vein) was carried out: ectopic expression of huMETCAM/MUC18 in human melanoma cells increased the pulmonary metastasis of non-metastatic human melanoma cells in xenograft mice [39-40] and mouse METCAM/MUC18 increased the pulmonary metastasis of non-metastatic mouse melanoma cells in syngeneic mice [41]. However, the effect of increased METCAM/ MUC18 expression on causing melanoma metastasis has never been observed when the spontaneous metastasis test (orthotopic injection of cells subcutaneously) is used, suggesting it affects the late stages of melanoma metastasis [39-41]. Taken together, METCAM/MUC18 does not initiate the metastasis of melanoma cells, thus it may not convert an indolent melanoma to an aggressive cancer. This notion is strengthened by that FGF-2 initiates the metastasis of melanoma [30].
Over-expression of huMETCAM/MUC18 is not limited to melanoma, as originally thought [28-29]. Two groups have attempted to study possible huMETCAM/MUC18 expression in prostate cancer cell lines and prostate cancer tissues but failed because the monoclonal antibodies they used could not recognize the huMETCAM/MUC18 epitopes presented in prostate cancer cell lines and tissues [31,42]. For similar studies we prepared necessary reagents, such as cloning and characterizing of and expressing recombinant proteins from the huMETCAM/MUC18 and mouse METCAM/MUC18 (moMETCAM/ MUC18) cDNAs and preparing anti-huMETCAM/MUC18 [13,22] and anti-moMETCAM/MUC18 [43] polyclonal antibodies in chicken. By using our chicken polyclonal antibodies we successfully found positive immunological signals in both prostate cancer cell lines and tissues; these results were further confirmed by the RT-PCR analysis of huMETCAM/MUC18 mRNA [14,22-27]. HuMETCAM/MUC18 is not expressed in the epithelium of normal prostate and in benign prostatic hyperplasia (BPH), but is expressed in prostatic intraepithelial neoplasia (PIN), prostatic adenocarcinoma, and metastatic lesions [14]. Its expression appears to correlate with the initiation of the malignant progression of clinical prostate cancer [13-14,22]. TRAMP (transgenic adenocarcinoma mouse prostate) is an excellent mouse model for prostate cancer research [44]. Transgenic mice were generated by microinjection of a recombinant gene construct, which contains the prostate-specific rat probasin (PB) promoter to drive expression of the simian virus 40 large tumor antigen coding region (Tag), into the male pro-nucleus of a fertilized one cell zygote; the transgene was successfully integrated into a zygote chromosome to form a transgenic mouse line for further propagation. In this mouse model, expression of the PB-Tag transgene is spatially restricted to the dorsolateral and ventral lobes of the male TRAMP mice. Mice expressing high levels of the transgene display progressive forms of prostatic disease that histologically resemble human prostate cancer, ranging from mild intraepithelial hyperplasia to large multinodular malignant neoplasia. Prostate tumors have been detected specifically in the prostate as early as 10 weeks of age. This transgenic mouse model, which reproducibly develops prostate cancer, also provides an animal model system to study the molecular basis of transformation of normal prostatic cells and the factors influencing the progression to metastatic prostate cancer [44]. By taking advantage of this TRAMP, we collaborated with Norman Greenberg’s group to study if mouse METCAM/MUC18 expression is correlated with the progression of mouse prostate adenocarcinoma. We found that indeed METCAM/MUC18 expression also correlates with the progression of prostate cancer and that of mouse prostate adenocarcinoma in a transgenic mouse model, TRAMP [23-24]. Taken together, we formulate a hypothesis that huMETCAM/MUC18 may be used as a novel diagnostic biomarker for early detection of the metastatic potential of human prostate cancer. We further suggest that huMETCAM/MUC18 may directly play an important role in the development and malignant progression of prostate cancer. This notion has been further supported by the evidence, as described in the following section [13-15,22-27].
Human METCAM/MUC18 is a driver for the malignant progression of prostate cancer
We have pioneered in exploring the biology of METCAM/MUC18- mediated malignant progression of prostate cancer. We have shown that increased huMETCAM/MUC18 expression increases the tumorigenesis and initiates the metastasis of human prostate cancer cells [13-15,22-27], as summarized in this section. From the evidence, we strongly suggest that huMETCAM/MUC18 is a bona fide metastasis gene that initiates the metastasis of human prostate cancer cells in nude mice [13-15,22-27]. All the following animal studies were approved by the ethics committee of animal studies, IACUC of Emory University and that of Chung Yuan Christian University. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
HuMETCAM/MUC18 expression increases tumorigenicity of human prostate cancer cells [14,26-27]
To further study the effect of increased expression of huMETCAM/ MUC18 on the tumor-take and tumorigenesis of LNCaP cells, we subcutaneously co-injected the cells with Matrigel to nude mice and found that increased huMETCAM/MUC18 expression increased tumor-take two-fold (Figure 2A) [26], decreased the tumor onset time 18 days, increased in vivo tumorigenicity ten to twelve-fold (Figure 2B), and increased the tumor weight five-fold, as shown in Figure 2C [26].
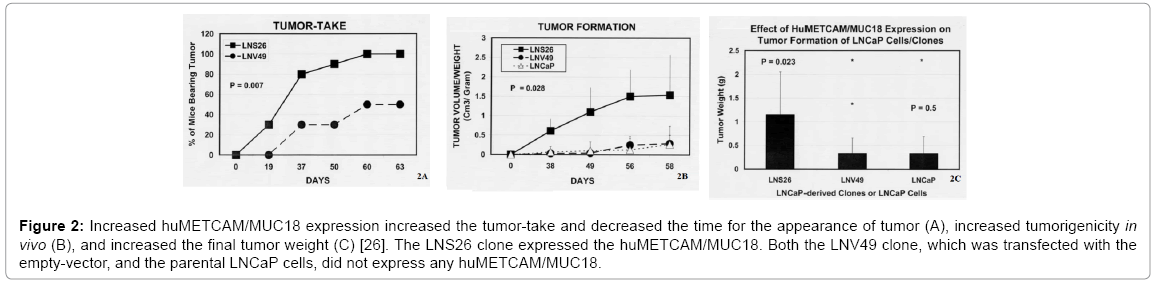
Figure 2: Increased huMETCAM/MUC18 expression increased the tumor-take and decreased the time for the appearance of tumor (A), increased tumorigenicity in vivo (B), and increased the final tumor weight (C) [26]. The LNS26 clone expressed the huMETCAM/MUC18. Both the LNV49 clone, which was transfected with the empty-vector, and the parental LNCaP cells, did not express any huMETCAM/MUC18.
HuMETCAM/MUC18 promotes epithelial-mesenchymal transition (EMT) of three human prostate cancer cell lines [22]
Epithelial-to-mesenchymal transition (EMT) is a biological process by which carcinoma cells (cancer cells derived from normal epithelial cells) detach from the surrounding tissue and acquire characteristics of mesenchymal cells, which are unique motile and spindle-shaped cells with end-to-end polarity [45]. Cells that have undergone EMT can migrate out of their epithelial layers to distant points and either remain mesenchymal or re-differentiate into epithelial cells by a process known as mesenchymal-to-epithelial transition (MET). Thus EMT may be a process pre-required for tumor progression. In addition to increased motility, carcinoma cells via EMT may become stem cell-like, protected from senescence, apoptosis and immune surveillance, and resistant to conventional and targeted therapies [45]. The degree of EMT in cells usually can be determined by the extent of increased motility and invasiveness of the cells in vitro. Since the three prostate cancer cell lines, DU145, PC-3, and TSU-PR1 (which was originally identified as a prostate cancer cell line, but later validated to be contaminated with bladder cancer cells), endogenously expressed METCAM/MUC18 at a level of 64%, 53%, and 1%, respectively, of that in SK-Mel-28 (endogenous METCAM/MUC18 as 100%) [22], one way to determine if METCAM/MUC18 directly plays a role in increasing motility and invasiveness is by reducing these two behaviors after treatment of these cell lines with an anti-METCAM/MUC18 antibody, which decreased the endogenous METCAM/MUC18 level. As shown in Figure 3, an anti-huMETCAM/MUC18 antibody could decrease the in vitro motility (Figure 3A) and invasiveness (Figure 3B) of these three human prostate cancer cell lines [22]. From the results of Figure 3, we suggested that huMETCAM/MUC18 may be a mediator for EMT of the two prostate cancer cell lines, DU145 and PC-3, and a bladder cancer cell line, TSU-Pr1, which endogenously express huMETCAM/ MUC18, but not LNCaP cell line, because it does not express the gene.
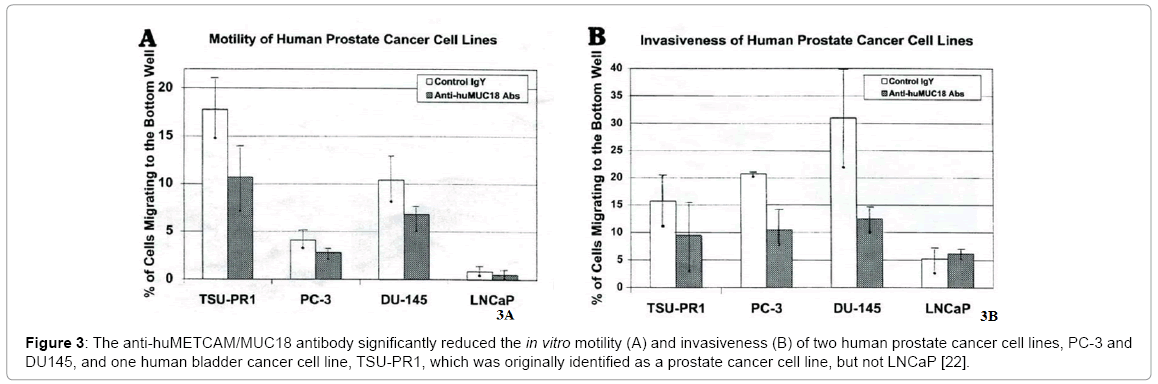
Figure 3: The anti-huMETCAM/MUC18 antibody significantly reduced the in vitro motility (A) and invasiveness (B) of two human prostate cancer cell lines, PC-3 and DU145, and one human bladder cancer cell line, TSU-PR1, which was originally identified as a prostate cancer cell line, but not LNCaP [22].
HuMETCAM/MUC18 also promotes EMT of the human prostate cancer cell line LNCaP [25]
Since the LNCaP cell line does not endogenously express any METCAM/MUC18 [22], to determine if METCAM/MUC18 plays a role in mediating EMT in this cell line, we ectopically increase the METCAM/MUC18 expression in this cell line and determine its in vitro motility and invasiveness. If METCAM/MUC18 plays a role in mediating EMT in this cell line, we should observe an increased motility and invasiveness of the stable LNCaP clones that overly express METCAM/MUC18. As shown in Figure 4, over-expression of huMETCAM/MUC18 in human prostate cancer cell line LNCaP huMETCAM/MUC18 indeed increased its in vitro motility (Figure 4A) and invasiveness (Figure 4B) [25]. From the results of Figure 4, we have strongly suggested that huMETCAM/MUC18 causes the EMT of LNCaP cells. As also shown in the Figure 4, the increased motility and invasiveness of LNCaP clones/cells was significantly reduced by the anti-huMETCAM/MUC18 antibody, suggesting that huMETCAM/ MUC18 directly causes the EMT of LNCaP cells.
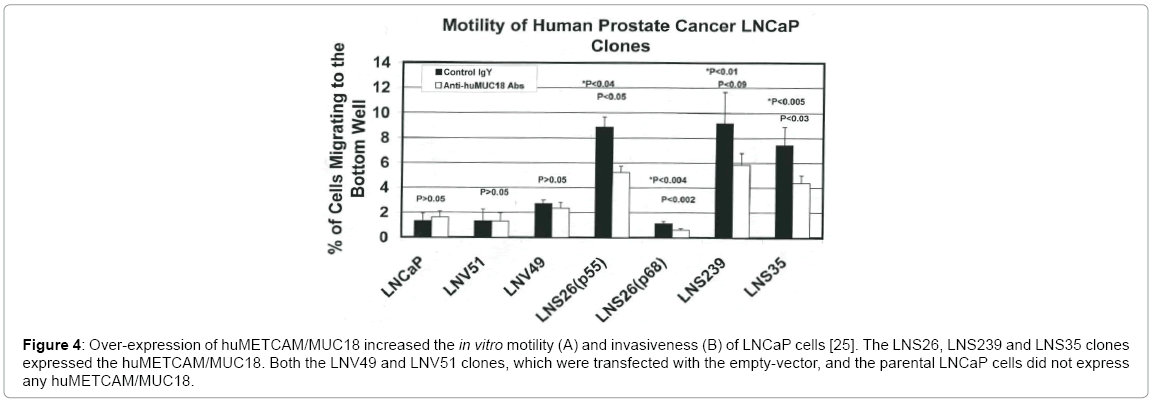
Figure 4: Over-expression of huMETCAM/MUC18 increased the in vitro motility (A) and invasiveness (B) of LNCaP cells [25]. The LNS26, LNS239 and LNS35 clones expressed the huMETCAM/MUC18. Both the LNV49 and LNV51 clones, which were transfected with the empty-vector, and the parental LNCaP cells did not express any huMETCAM/MUC18.
Ectopic expression of huMETCAM/MUC18 initiates the metastasis of prostate cancer cells to multiple organs [25]
We injected huMETCAM/MUC18-expressing LNCaP clones and control clones into the dorsolateral lobes of mouse prostates of male nude mice to study the effect of over-expression of huMETCAM/ MUC18 on the progression of LNCaP cells. We have found that overexpression of huMETCAM/MUC18 promotes the metastasis of LNCaP cells to periaortic lymph nodes and also to multiple urogenital organs, such as seminal vesicles, urethers, and kidneys (Table 1 and Figure 5) [25]. Since the vector control clones and the parental cells did not have any metastatic lesions, though they can induce prostatic tumors, we suggest that over-expression of huMETCAM/MUC18 initiates the metastasis of LNCaP cells and it profoundly affects many steps of metastasis of LNCaP cells.
| Cell line | Level of human METCAM/MUC18 expression |
Tumor take, Number of mice with tumor (%)* |
Tumor size (g), mean value (±standard deviation) | Number of mice with metastatic lesions to (target organs) |
|---|---|---|---|---|
| LNCaP (p40) | none | 2/6 (33%) | 1.85 (±1.77) | 0/6 (none) |
| LN-2-6 (p49) | ++++ | 3/9 (33%) | 1.5 (±1.32) | 3/9 (peri-aortic lymph nodes, ureter, kidney, seminal vesicles) |
| LN-2-6 (p64) | + | 1/5 (20%) | 3 | 1/5 (peri-aortic lymph nodes, ureter, kidney, seminal vesicles) |
| LN-2-39 (p57) | ++++ | 3/5 (60%) | 2.57 (±2.45) | 3/5 (peri-aortic lymph nodes, ureter, kidney, seminal vesicles) |
| LN-3-5 (p58) | ++++ | 3/4 (75%) | 1.8 (±2.7) | 3/4 (peri-aortic lymph nodes, ureter, kidney, seminal vesicles) |
| LN-4-1 (p55) | none | 1/5 (20%) | 0.7 | 0/5 (none) |
| LN-5-1 (p57) | none | 1/5 (20%) | 8.6 | 0/5 (none) |
*1 × 106 cells were injected orthotopically (to the dorsolateral lobe) of male nude mice.
Table 1: Metastasis of huMETCAM/MUC18-expressing LNCaP clones to multiple organs [25].
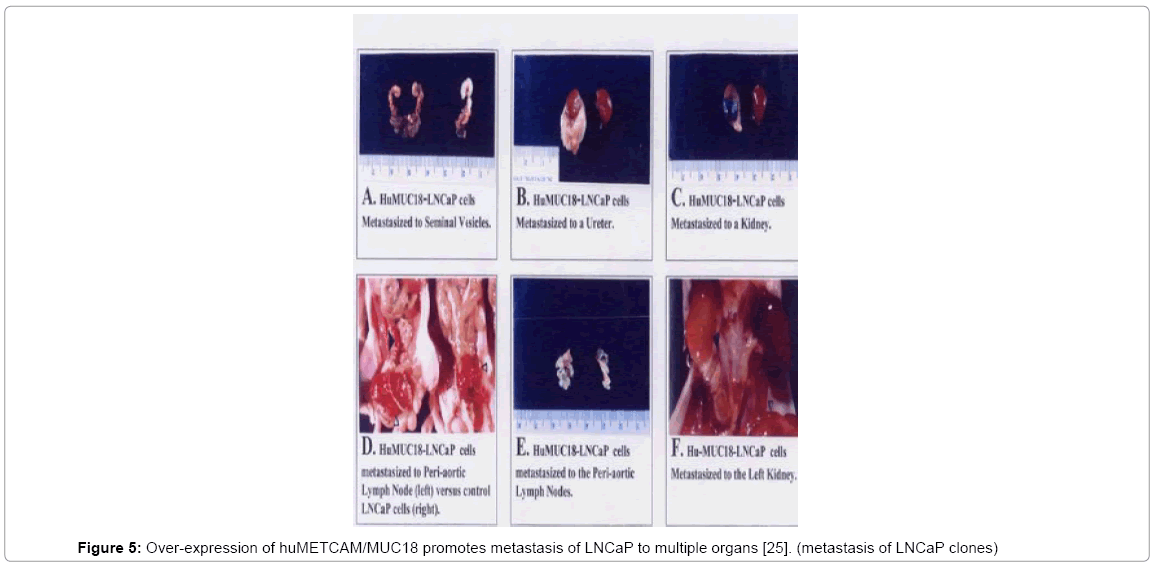
Figure 5: Over-expression of huMETCAM/MUC18 promotes metastasis of LNCaP to multiple organs [25]. (metastasis of LNCaP clones)
Over-expression of huMETCAM/MUC18 promotes the growth of human prostate cancer LNCaP cells in tibia bone marrow
Prostate cancer patients have a high incidence of bone metastasis [46], but the role of huMETCAM/MUC18 in this process has not been investigated. We suggest that huMETCAM/MUC18 very likely plays a role in the bone metastasis of prostate cancer cells based on three evidence: (a) Putz et al. [42] found that all the cell lines established from bone micrometastases expressed huMETCAM/MUC18, suggesting that huMETCAM/MUC18 plays an important role in establishing bone colony formation, (b) huMETCAM/MUC18 is expressed in the endothelial cells lining of bone marrow [47,48], and (c) our findings that enforced expression of huMETCAM/MUC18 promotes LNCaP clones/ cells to metastasize to multiple organs [25]. To test this hypothesis, we have carried out intra-tibial injection [49] of both METCAM/MUC18- expressing LNCaP and C4-2B clones/cells (as shown in Figure 6) to investigate if over-expression of huMETCAM/MUC18 in these clones/ cells can increase their abilities to establish bone colonies. Figure 7 and Table 2 show that the mice intra-tibially injected with cells from both clone LNCaP#25 (LNLC-MC) and clone C4-2B#31(C4-2B-MC), which had been transfected with huMETCAM/MUC18 cDNA and hence had an increased huMETCAM/MUC18 expression (Figure 6), had an increased PSA level in the serum when compared to the mice injected with the cells of both LNCaP-vector (LNLC-V) and C4-2B-vector (C4-2B-V), which were transfected with the empty vector and hence no METCAM/MUC18 expression. The increased serum PSA level indicated growth of those cells in the tibia bone marrow of the injected mice [50]. Both Figure 7 and Table 2 also showed that the serum PSA
| Clones | METCAM/MUC18 Expression | Serum PSA level (fold) | Number of mice positive (%) |
|---|---|---|---|
| LNCaP#25 (LNLC-MC) | Yes | 1.24 ± 0.36 ng/ml (2.88) | 7/7 (100) |
| LNCaP-Vector (LNLC-V) | No | 0.43 ± 0.49 ng/ml (1) | 2/5 (40) |
| C4-2B#31 (C4-2B-MC) |
Yes | 6.2 ± 9.2 ng/ml (14.4) | 6/10 (60) |
| C4-2B-Vector (C4-2B-V) | No | 0.43 ± 0.32 ng/ml (1) | 2/6 (30) |
Table 2: Bone colony formation of huMETCAM/MUC18-expressing LNCaP and C4-2B clones and vector control clones after intra-tibial injection [50].

Figure 6: Expression of METCAM/MUC18 in LNCaP and C4-2B clones [50]. LNCaP and C4-2B cell lines were transfected with the huMETCAM/MUC18 cDNA, stable G418-resisitant clones were isolated, and METCAM/MUC18 expression was determined by Western blot analysis. Lanes 6 and 8 show the expression levels of METCAM/MUC18 in two typical high expression clones (LNCaP#25 and C4-2B3#31) derived from LNCaP and C4-2B cell lines, respectively. Lanes 5 and 7 show no expression of METCAM/MUC18 in the vector-transfected LNCaP and C4-2B3 clones. Lanes 1-4 show variable expression level of METCAM/MUC18 in SK-Mel-28, DU145, PC-3, and LNCaP cell lines.
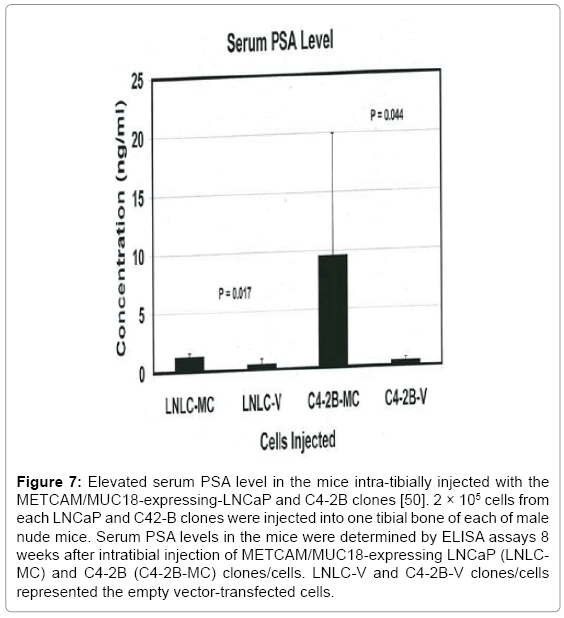
Figure 7: Elevated serum PSA level in the mice intra-tibially injected with the METCAM/MUC18-expressing-LNCaP and C4-2B clones [50]. 2 × 105 cells from each LNCaP and C42-B clones were injected into one tibial bone of each of male nude mice. Serum PSA levels in the mice were determined by ELISA assays 8 weeks after intratibial injection of METCAM/MUC18-expressing LNCaP (LNLCMC) and C4-2B (C4-2B-MC) clones/cells. LNLC-V and C4-2B-V clones/cells represented the empty vector-transfected cells.
level was increased more dramatically in C4-2B cells (with a 14.4-fold increase) by the enforced expression of METCAM/MUC18, but less dramatically in the parental LNCaP cells (with a 3-fold increase). Since C4-2B cells, which were established from bone metastasis of LNCaP cells in androgen-deprived male nude mice [51], are LNCaP-derived bonehoming cells, this indicates that enforced expression of huMETCAM/ MUC18 in LNCaP cells may increase their bone metastasis and even more dramatically for the bone-homing C4-2B cells.
METCAM/MUC18-specific shRNAs reduced the tumorigenesis of human prostate cancer cell line DU145 in an Athymic nude mouse model [52]
To investigate if METCAM/MUC18 also plays a positive role in the progression of other prostate cancer cell lines besides LNCaP, we determined knock-down effects of shRNAs, which reduced endogenous METCAM/MUC18 expression in DU145 cell line, on in vivo tumorigenesis in male nude mice. After transfecting DU145 cells with a lentivirus vector pGIPZ (Thermo Scientific), which carried one of three METCAM/MUC18-specific shRNAs (#72, 24, and 27) or a negative control shRNA (#46) (as shown in Table 3), four pooled puromycin-resistant DU145 clones/cells were obtained and subjected to western blot analysis. We found shRNAs, #24 and 27, more efficiently reduced the endogenous METCAM/MUC18 expression than shRNA #72 (Figure 8) [52]. Effect of decreased METCAM/MUC18 expression on tumorigenicity of DU145 cells was determined by weekly monitoring tumor volumes after SC injection of these pooled clones into male nude mice. As shown in Figure 9. Two pooled clones derived from shRNAs, #24 and #27, supported tumorigenesis of DU145 cells much less efficiently than the control clone #46 and the clone #72. Comparing to the control clone #46, final tumor weights of clones #72, #24, and #27 were reduced to 53%, 23%, and 9.1%, respectively (Figure 10). Taken together, we concluded that METCAM/MUC18 also promotes tumorigenesis of prostate cancer DU145 cell line, similar to it in the LNCaP cell line [52].
| Short Name | Catalogue number of Thermo Sci | Sequence of sense strand | Location in METCAM cDNA |
|---|---|---|---|
| 46 | RHS4346 (non-silence control) | 5’-ATCTCGCTTGGGCGAGAGTAAG-3’ | none |
| 72 | RHS4430-200292372 (shRNA 392652) | 5’-TGGTCTTGTTCACTTGCCG-3’ | 1435-1453 |
| 24 | RHS4430-200288324 (shRNA 392649) | 5’-TCAACTACAAGTTCGCTCT-3’ | 1843-1861 |
| 27 | RHS4430-200295227 (shRNA 382648) | 5’-TTGTCTTCTTTAACCAGCT-3’ | 661-679 |
Table 3: METCAM/MUC18-specifc shRNA inserts in the lentivirus vector pGIPZ [52].

Figure 8: METCAM/MUC18 expression in METCAM/MUC18-specific shRNAs transfected DU145 clones [52].
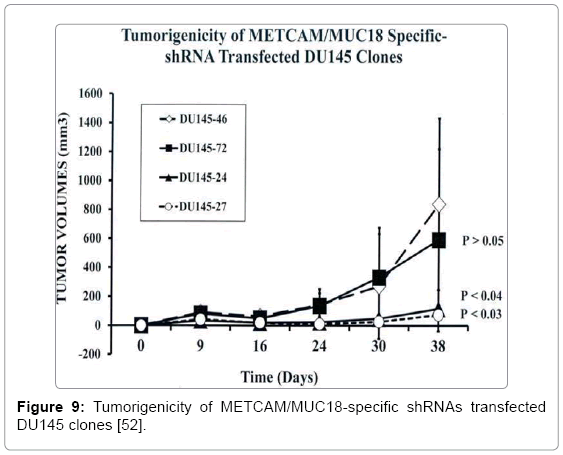
Figure 9: Tumorigenicity of METCAM/MUC18-specific shRNAs transfected DU145 clones [52].
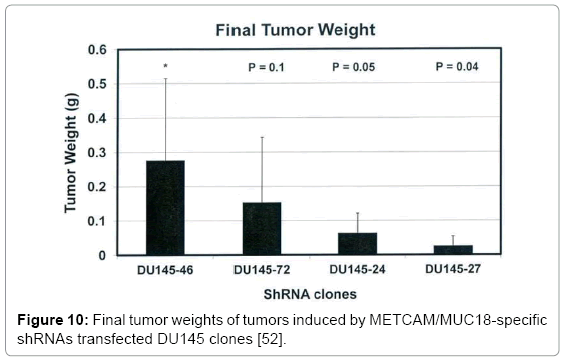
Figure 10: Final tumor weights of tumors induced by METCAM/MUC18-specific shRNAs transfected DU145 clones [52].
Possible mechanisms for the METCAM/MUC18-mediated malignant progression of prostate cancer cells
The mechanisms of which huMETCAM/MUC18 expression affects tumorigenesis and metastasis of prostate cancer cells are poorly understood. By deducing knowledge learned from other tumors [7- 8,18-19] and functions of huMETCAM/MUC18 in endothelial cells and melanoma [27,33-34], huMETCAM/MUC18 expression may increase tumorigenesis by cross-talk with many signaling pathways that affect survival and proliferation and angiogenesis of tumor cells [15,27]. We therefore predicted that enforced expression of huMETCAM/MUC18 may increase tumorigenesis by affecting its key downstream effectors, such as decreasing apoptosis and increasing survival and proliferation of the cells. For this purpose, expression levels of apoptotic indexes, proliferative indexes, the survival index, and angiogenesis indexes in subcutaneous (S.C.) tumors were determined and compared to those in prostatic tumors obtained previously. We found that enforced expression of human METCAM/MUC18 appeared to render the cells with increased levels of proliferation indexes, Ki67 and PCNA, a survival index, phospho-AKT, and angiogenesis indexes, VEGF, VEGFR2, and CD31; however, it did not significantly render the cells with altered levels of various apoptosis indexes [26]. Taken together, we suggest that METCAM/MUC18 mediates tumorigenesis of human prostate cancer cells via increasing their proliferating potential, upregulating the AKT-signaling pathways, and augmenting angiogenesis. METCAM/MUC18 may mediate metastasis of human prostate cancer cells via increasing expression of MMP 2. However, many other detailed mechanisms mediated by this Ig-like CAM [53,54], which drives the malignant progression of prostate cancer cells, require further studies in the near future.
Perspectives and possible clinical applications
Over-expression of METCAM/MUC18 is correlated with the malignant progression of clinical prostate cancer and mouse prostate adenocarcinoma in the transgenic model, TRAMP suggesting that it may be a useful novel biomarker for the diagnosis of the malignant potential of prostate cancer at early stages. Thus it potentially can replace the PSA test in the diagnosis of the malignant potential of prostate cancers after further simplification of the detection method. Since it can promote tumorigenesis and initiate metastasis of human prostate cancer cells in a preclinical mouse model, we suggest that it is also a driver for the malignant progression of prostate cancer cells. Preliminary studies show that METCAM/MUC18 mediate tumorigenesis and metastasis of human prostate cancer cells via increasing their proliferating potential, up-regulating the AKT-signaling pathways, and augmenting angiogenesis. This mouse model can also be used for a real-time observation of the dynamic process of prostate cancer progression by using a non-intrusive and high photo-penetrating imaging system to further understand the progression of the cancer [55]. The knowledge gained should also be useful for designing effective means to decrease the metastatic potential, or even to block the metastatic process of the cancer cells. Along these lines, a preclinical trial of using doxazosin, a 1-adrenergic antagonist that has been used to treat the BPH patients, has been shown to be able to suppress prostate cancer metastasis in the TRAMP mouse model [24]. Furthermore, preclinical trials using a fully humanized anti-METCAM antibody against melanoma growth and metastasis [56] and using a mouse anti-METCAM monoclonal antibody against angiogenesis and tumor growth of hepatocarcinoma, leiomyosarcoma, and pancreatic cancer [57] have been successfully demonstrated, these antibodies may be used to block the progression of prostate cancer. Alternatively, small soluble peptides derived from METCAM may also be useful for blocking the tumor formation and tumor angiogenesis of the cancer [58]. The attachment of these reagents to nanoparticles may be another alternative for diagnostic and therapeutic use [59]. Recently, we also found that METCAM/ MUC18-specific RNAs in a lentivirus vector could efficiently reduce the endogenously expressed METCAM/MUC18 and decreased the tumorigenesis of another human prostate cancer cell line DU145 in an athymic nude mouse model [52]. The success of reducing tumor formation of human prostate cancer cell lines in a preclinical mouse model may extent the use of METCAM/MUC18-specific siRNAs for clinical trial to block the progression of clinical prostate cancer [60].
I thank financial supports from Emory University School of Medicine (USA), Chung Yuan Christian University, and grants from NSC (NSC-101-2320-B-033-001 and -003), Taiwan (GJW).