Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
![]() +44 1300 500008
+44 1300 500008
ISSN: 2167-0412
![]() +44 1300 500008
+44 1300 500008
Research Article - (2016) Volume 0, Issue 0
The aim of the present in vitro study was carried out to evaluate the anti-inflammatory activity of Hypericum triquetrifolium extracts (HT-extract) on lipopolyssacharide -stimulated human monocytic (THP-1) cells and human peripheral blood mononuclear cells (PBMNCs). The expression and production of pro-inflammatory cytokines tumor necrosis factor-α (TNF-α) and interleukin 6 (IL-6), as well as the anti-inflammatory cytokine interleukin 10 (IL-10) were evaluated by assessing the levels of proteins and mRNA’s of TNF-α, IL-6 and IL-10 in both cell types. Cells were exposed to 5 μg lipopolyssacharide (LPS) /ml in the absence and presence of increasing concentrations of 50% ethanol extracts from the aerial parts of Hypericum triquetrifolium. The anti-inflammatory efficacy experiments were performed with HT-extract concentrations up to 250 μg/ml that had no cytotoxic effects as assessed with MTT and LDH assays. HT-extract remarkably inhibited the expression and secretion of TNF-α and IL-6 at a concentration of 250 μg/ml. HT-extract remarkably elevated IL-10 secretion and mRNA levels at 125 μg/ml. Furthermore, HT-extract exhibited relatively high antioxidant activity (IC50 of 5 μg/ml) as measured with DPPH assay. These findings indicate that HT-extract probably exerts anti-inflammatory effects at both protein and gene expression levels of the anti-inflammatory as well of the pro-inflammatory cytokines in PBMNCs.
Keywords: Anti-inflammatory; Hypericum triquetrifolium; PBMNCs; Medicinal plants
The Inflammation is a physiological response that is essential for eradicating pathogens. It is mediated by pro-inflammatory mediators, such as interleukin-1 (IL-1), the tumor necrosis factor alpha (TNF-α), gamma-interferon (IFN-γ), IL-12, and IL-18, and is suppressed by anti-inflammatory mediators, such as IL-4, IL-10, and the transforming growth factor (TGF-β) [1-3]. Anti-inflammatory cytokines block inflammatory processes or at least suppress the intensity of the pro-inflammatory cytokines. Therefore, a “balance” between the production levels of the two types of cytokines is thought to determine the outcome of an inflammatory disease. Numerous in vitro studies showed that IL-10 inhibits the production of pro-inflammatory cytokines such as IL-1, IL-6, and TNF-α by LPS-activated macrophages [4,5]. Thus, inhibitors of the pro-inflammatory cytokines (e.g., IL-1, IL-6 and TNF-α) have been considered as potential candidates for the development of anti-inflammatory drugs [3-8].
Despite the great achievements in the conversional medicine, traditional medicine has always been utilized in the Mediterranean region. Greco-Arab and Islamic medicine had shown remarkable success in treating inflammatory conditions in general, and clearly distinguishing between several subtypes of inflammatory diseases and has offered large number of herbs for curing all types of inflammatory diseases [9-12]. Recently, there have been many in vitro and in vivo investigations concerning herbal-derived extracts as well as phytochemicals with anti-inflammatory effects including but not limited to Cinnamomum cassia, Peganum harmala,Punica granatum, Urtica dioica and Vitis vinifera [13-16]. Hypericum triquetrifolium based products have been used in traditional Greco-Arab medicine to cure various inflammatory conditions [9-12]. According to our previous ethano pharmacological study, Hypericum triquetrifolium is still used within the practitioner communities in the Galilee and in the West Bank [9-12]. Based on traditional Greco-Arab herbal medicine and on recent in vitro and in vivo reports in which HT-extract exhibited anti-inflammatory effects by reducing the expression of pro-inflammatory cytokines and nitric oxide production [15,17], this study explored the anti-inflammatory mechanism of Hypericum triquetrifolium. Therefore, the expression and production of pro-inflammatory cytokines tumor necrosis factor-α (TNF-α) and interleukine 6 (IL-6), the anti-inflammatory cytokine IL-10 were assessed in LPS-activated human monocytic (THP-1) cells as well as in in primary human PBMNCs.
Our results indicate that Hypericum triquetrifolium could modulate the regulatory mechanism of pro-inflammatory cytokines (TNF-α and IL-6) as well as the anti-inflammatory cytokine (IL-10) in the LPS-activated THP-1 cells and in primary human PBMNCs. Hypericum triquetrifolium inhibited the production and expression of IL-6 and TNF-α, and increased the secretion and expression of IL-10. In addition, HT-extract exhibited relatively high antioxidant activity as measured with DPPH assay.
Preparation of Hypericum triquetrifolium extracts (HT-extract)
The ethanolic extract of HT was prepared as follows: 100 grams of air-dried areal parts of HT were added to 1000 ml of 50% ethanol in water and boiled for 10 minutes. The boiled extracts were filtered through filter paper and the filtrate was freeze-dried in a lyophilizer. The freeze-dried crude extracts were kept at -70°C [14].
DPPH scavenging assay
DPPH radical scavenging activity assay was carried out as described in Ref. [18] with slight modifications. Briefly, 825 μL of 100 µM ethanol solution of 1,2-diphenyl-1-picrylhydrazyl (DPPH) solution was added to 25 μl of a dilution series of the plants extracts ranging from (0.3-40 mg/mL) and mixed vigorously, then incubated in dark for 1 hr. The absorbance was estimated in 1 ml cuvettes at 517 nm using a Perkin Elmer Lambda 40 UV/VIS spectrophotometer against ethanol blank where DPPH and extract were absent. The experiments were performed in triplicates and average absorption was noted for each concentration. Butyl hydroxyl toluene (BHT) was used as a standard. The percent inhibitions of the DPPH radical by the samples was calculated based on the below formula:
(The absorbance of negative control–The absorbance of the sample)/The absorbance of negative control × 100
The standard graph was plotted against sample concentration taking the concentration of (BHT) as a positive control on the x-axis and percentage scavenging activity on the y-axis. Based on this standard graph, IC50 was determined (concentration of sample able to scavenge 50% of DPPH free radical).
Cell culture
The human monocytic cell line THP-1 (ATCC 202-TIB) was purchased from American Type Culture Collection (Manassas, VA, USA). These cells are known to express various monocytes receptors and have been widely used as a model system for macrophage research. Cells were maintained in DMEM (Dulbecco’s modified Eagle’s medium) supplemented with 10% vol/vol inactivated fetal calf serum (FCS), 1% nonessential amino acids, 1% glutamine, 100 U/mL penicillin, 10 µg/ml streptomycin and kept in a humidified atmosphere of 5% CO2 at 37°C.
Isolation of Peripheral Blood Mononuclear Cells (PBMNCs)
Blood samples were taken from 16 healthy volunteer students from the Arab America University (Jenin, Palestine), aged 19-21 years (7 males and 9 females).
Blood samples were withdrawn in heparin tube after filling consent and a written questionnaire. Venous blood (15 mL) was processed immediately after collection. Peripheral blood mononuclear cells (PBMNCs) were isolated using gradient centrifugation in Histopaque-1077 solution (Sigma-Aldrich). Separated PBMNCs were incubated in RPMI-1640 media supplemented with sodium bicarbonate, L-glutamine-penicillin-streptomycin solution (200 mM L-glutamine, 10,000 U penicillin and 10 mg streptomycin/mL in 0.9% NaCl) and 10% vol/vol FCS. Isolated PBMNCs were seeded at a cell density of 1 × 106 cells/ml in 24-well plates and exposed to HT-extract (125 µg/mL and 250 µg/ml) in a fresh serum-free medium in the absence and presence of from Escherichia coli serotype O127:B8 (5 µg/mL). Cells were maintained at 37°C for 4, 6 and 20 hours at 5% CO2 and the levels of secreted IL-6, TNF-α and IL-10 were determined as described below.
MTT assay
MTT (Tetrazolium dye) is widely used to measure the viability and/or the metabolic state of cultured cells [19]. Twenty-four hours after cell seeding, cells were treated with varying concentrations of HT-extracts for 24 hours at 37°C. Cells were then washed in phosphate buffered saline, incubated in serum-free RPMI to which MTT (500 µg/mL) was added to each well (100 µL), and incubated for a further four hours. After removal of medium, the cells were incubated for 15 minutes with 100 µl of acidic isopropanol (0.08 N HCl) to dissolve the formazan crystals. The absorbance of the dissolved MTT formazan was measured at 570 nm in an Elisa reader. Viability was defined as the ratio (expressed as a percentage) of absorbance of treated cells (HT-extracts up to 500 µg/mL) to untreated control cells.
Lactate dehydrogenase
In the Lactate dehydrogenase(LDH) assay the release of this exclusively cytosolic enzyme in the cell culture medium is a widely used indicator of cell membrane damage [20].
For the LDH assay, THP1 cells were seeded in well of 96-microtiter plates at a cell density of 5 × 103/100 µL. Twenty-four hour after cell seeding, cells were treated with HT-extracts at concentrations of 0-250 µg/mL. The supernatants were collected 24 h after treatment and cell monolayers were then lysed with a cell lysis solution for 30 minutes at room temperature. LDH activity was determined in both the supernatants and the cell lysate fractions by using CytoTox 96, a non-radioactive cytotoxicity assay kit (Promega, WI, USA) in accordance with the manufacturer’s instruction. The absorbance was measured at 490 nm with 96-well plate ELISA reader. The LDH release from the cells was determined using the formula
For the LDH assay, THP1 cells were seeded in well of 96-microtiter plates at a cell density of 5 × 103/100 µL. Twenty-four hour after cell seeding, cells were treated with HT-extracts at concentrations of 0-250 µg/mL. The supernatants were collected 24 h after treatment and cell monolayers were then lysed with a cell lysis solution for 30 minutes at room temperature. LDH activity was determined in both the supernatants and the cell lysate fractions by using CytoTox 96, a non-radioactive cytotoxicity assay kit (Promega, WI, USA) in accordance with the manufacturer’s instruction. The absorbance was measured at 490 nm with 96-well plate ELISA reader. The LDH release from the cells was determined using the formula
Immunoassay for cytokines
The amounts of secreted TNF-α, IL-6, and IL-10 were measured using a commercial ELISA kits (R&D Systems, Minneapolis, MN, USA). The absorbance at 450 nm was read by a microplate reader (Model 680; Bio-Rad Laboratories, Mississauga, ON, Canada) with the wavelength correction set at 550 nm. The amounts of TNF-α, IL-6, and IL-10 were calculated with the help of a standard curve, which was constructed using serial dilutions of cytokine standards provided with the kit.
RT-PCR analysis
For the determination of cytokine mRNA expression, cells were seeded at a density of 2 × 105 cell/mL in 24-well plates. Cells were activated with PMA (100 ng/mL) and Vitamin D3 (0.1 µM). Twenty-four hours after cell activation, the obtained THP-1-derived macrophages were treated with various concentrations of the HT- extracts in a fresh serum-free culture medium in the absence and presence of LPS (5 µg/mL).
Total RNA was obtained from cells using the Rneasy Plus Mini Kit (QIAGEN) according to manufacturer's instructions, and immediately frozen at -80°C until use. DNase-treated RNAs were used to synthesize cDNA with the Transcriptor First Strand cDNA Synthesis Kit using random hexamers as specified by the manufacturer (Maxima First Strand cDNA Synthesis Kit for RT-qPCR by Thermo). All RNA samples from a single experimental setup were reverse transcribed simultaneously and in duplicates, to minimize variation in the reverse transcription reaction.
PCR amplifications and Advanced relative quantification analysis were achieved using a Light Cycler 480 instrument (Roche Applied Science) with software version LCS480 1.5.0.39. All reactions were performed in duplicates with the Light Cycler Fast Start DNA Master SYBR Green I kit (Roche Applied Science) in a final 20 µl volume with 2.5mM MgCl2, 0.2 µM of each primer and 2 µL cDNA. Total RNA (0.1 μg) were used for a single reaction. Nucleotide sequences of oligonucleotide primers for the housekeeping glyceraldehyde-3-phosphate dehydrogenase (GAPDH) plus the IL-6, IL-10, or TNF-α primers pairs were used for RT-PCR, which were described elsewhere respectively [21]. Amplification conditions consisted of an initial pre-incubation at 95°C for 10 min (polymerase activation) followed by amplification of the target cDNA for 45 cycles (95°C for 15 s, 60°C for 20 s and extension time at 72°C for 30 s).
Statistical analysis
Error bars plotted in the figures represent simple standard deviations of the mean. When comparing different samples, results were considered to be statistically different when P<0.05 (Student’s t test for unpaired samples).
The cytotoxic activities of HT-extract on THP-1 cells and PBMNCs
The anti-inflammatory properties of HT-extracts were assessed in the present in vitro study using cells from the human monocyte cell line, which were differentiated to macrophages with the help of PMA (100 ng/mL) and Vitamin D3 (0.1 µM). The cell viability tests MTT and LDH assays were used to determine the non-toxic concentrations of the HT-extracts.
MTT test measures the activity of the mitochondrial enzyme succinate dehydrogenase. This colorimetric assay is widely used to evaluate the toxicity of herbal extracts. We used this test in order to evaluate the none-toxic concentrations of HT-extracts. THP-1-derived macrophages were treated with increasing concentrations (1-500 µg/mL of culture medium) of HT-extracts for 24 h. No significant reduction in cell viability was seen after treatment with HT-extracts up to 250 µg/mL (Figure 1). Concentrations higher than 250 µg/mL caused a significant reduction in the cell viability.
Lactate dehydrogenase activity(LDH-release test) is a widely used enzymatic assay to evaluatethe effects of medicinal plants onmembrane integrity (Cell viability). Lactate dehydrogenase, an enzyme located in the cytoplasm, catalyses the conversion of lactate and pyruvate. There are two possible causes that can lead to an elevation in the concentrations of cytoplasmatic lactate dehydrogenase; the first is cellular death and the second is a 'leak' in a plasma membrane. Results obtained indicate no significant increase in LDH amounts in the culture media after treatment with HT-extracts up to 125 µg/mL. A slight, but not significant elevation of LDH was seen after treatment with concentration of 125 and 250 µg/mL (Figure 1).
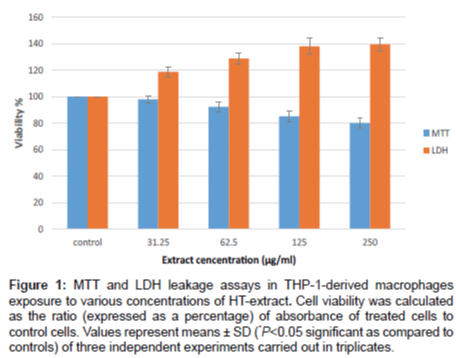
Figure 1: MTT and LDH leakage assays in THP-1-derived macrophages exposure to various concentrations of HT-extract. Cell viability was calculated as the ratio (expressed as a percentage) of absorbance of treated cells to control cells. Values represent means ± SD (*P<0.05 significant as compared to controls) of three independent experiments carried out in triplicates.
Based on these results, HT-extract concentrations of 500 mg/mL were excluded and concentrations below 250 µg/mL were used in the following experiments.
DPPH radical scavenging activity
The radical scavenging activity of three HT-extract (Methanol, Hexan and 50% Ethanol) was determined in order to evaluate the antioxidant activity of the HT-extracts. The radical scavenging activities are shown in Figure 2. As it is known, the lower the IC50 value the higher the antioxidant activity of the antioxidants. HT-extract exhibited strong antioxidant activity (IC50 of 5 µg/mL for 50% ethanol extract and methanol, and 7.5 µg/ml for hexane extract), which is equal to that of vitamin C (5.1 µg/mL).
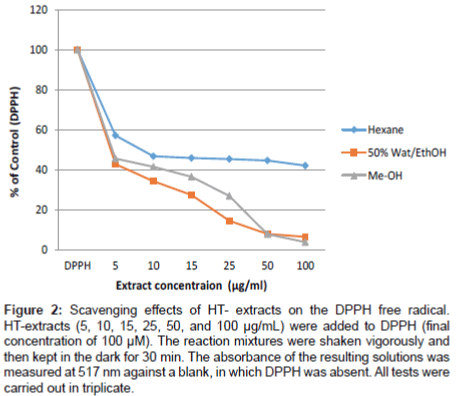
Figure 2: Scavenging effects of HT- extracts on the DPPH free radical. HT-extracts (5, 10, 15, 25, 50, and 100 μg/mL) were added to DPPH (final concentration of 100 μM). The reaction mixtures were shaken vigorously and then kept in the dark for 30 min. The absorbance of the resulting solutions was measured at 517 nm against a blank, in which DPPH was absent. All tests were carried out in triplicate.
The anti-inflammatory effects of HT-extract
The concentrations of IL-6, IL-10 and TNF-α in culture media of freshly isolated PBMNCs were measured using commercial enzyme-linked immunosorbent assay (ELISA) kits. Treatment of PBMNCs with HT-extract alone did not affect the levels of any of the studied cytokines (data not shown). Figure 3 shows the TNF-α and IL-6 secretion into the culture supernatant of untreated and LPS treated PBMNCs cells, respectively. HT-extract inhibited the secretion of the two pro-inflammatory cytokines in a dose dependent manner. LPS treatment of PBMNCs cells increased TNF-α and IL-6 secretion by 3.79 folds (484 ± 68 pg/mL) and 2.58 folds (517 ± 40 pg/mL), respectively. However, when those stimulated cells were treated with HT-extract at 125 µg/ml and 250 µg/mL, TNF-α and IL-6 secretions were decreased by 40% (268 ± 40 pg/mL) and 60% (180 ± 20 pg/mL), respectively. No significant cytotoxic effects were evident at these extract concentrations (Figure 1). HT-extract increased the secretion of the anti- inflammatory cytokine, IL-10 LPS-stimulated PBMNCs reaching by 121 ± 12 pg/mL and 79 ± 15 pg/mL at none cytotoxic concentration of 125 µg/mL and 250 µg/mL (Figure 4), respectively.
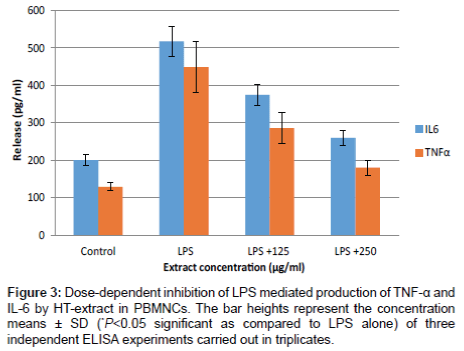
Figure 3: Dose-dependent inhibition of LPS mediated production of TNF-α and IL-6 by HT-extract in PBMNCs. The bar heights represent the concentration means ± SD (*P<0.05 significant as compared to LPS alone) of three independent ELISA experiments carried out in triplicates.
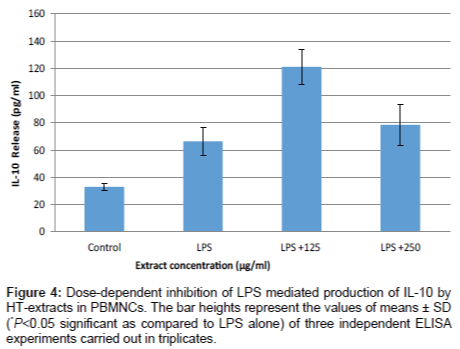
Figure 4: Dose-dependent inhibition of LPS mediated production of IL-10 by HT-extracts in PBMNCs. The bar heights represent the values of means ± SD (*P<0.05 significant as compared to LPS alone) of three independent ELISA experiments carried out in triplicates.
TNF-α and IL-6 mRNA levels were elevated upon LPS treatment by 4.2 and 4.5, respectively. When the PBMNCs cells were treated with 62 µg/ml and 125 µg/mLHT-extract, the cytokine expression was dramatically reduced. IL-6 and TNF-α were reduced about 2.2 folds. In contrast, Il-10 was increased by 65% after treatment of LPS-stimulated PBMNCs (Figure 5).
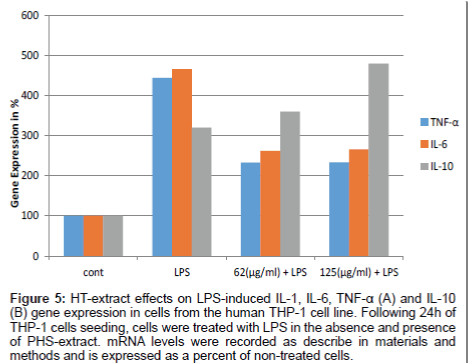
Figure 5: HT-extract effects on LPS-induced IL-1, IL-6, TNF-α (A) and IL-10 (B) gene expression in cells from the human THP-1 cell line. Following 24h of THP-1 cells seeding, cells were treated with LPS in the absence and presence of PHS-extract. mRNA levels were recorded as describe in materials and methods and is expressed as a percent of non-treated cells.
Hypericum triquetrifolium-derived products have been used in traditional Greco-Arab herbal medicine in the treatment various inflammatory diseases. However, only limited numbers of studies have been conducted to elucidate the action mechanisms of these productson inflammatory diseases [15,17]. LPS-activated human peripheral blood mononuclear cells and LPS-activated THP-1-derived macrophages, which represent appropriate model systems to study anti-inflammatory effects of potential natural remedies, were utilized here to examine the role of pro-inflammatory as well as anti-inflammatory cytokines in the observed anti-inflammatory effects of HT-extracts. Lipopolysaccharide (LPS)-activated macrophages finds a widespread use in the evaluation of anti-inflammatory properties of potential anti-inflammatory herbal-based extracts and compounds. The endotoxin LPS is a principle lippopolysaccharide of the outer membrane of Gram-negative bacteria that stimulates the production of pro-inflammatory cytokines such as IL-1, IL-6, TNF-α, leukotrienes, and nitric oxide (NO) [22-24]. The anti-inflammatory effects of HT-extracts were assessed here by measuring the expression and release of pro-inflammatory cytokines TNF-α and IL-6, and the anti-inflammatory cytokine IL-10.
Inflammation diseases are mediated through the action of pro-inflammatory cytokines (TNF-α, IL-1 and IL-6) and by eicosanoid such as PGE2. TNF-α and IL-1 are main contributors to chronic inflammatory diseases, including rheumatoid arthritis [24]. Receptor antagonists of the pro-inflammatory cytokines have been clinically successful to improve the symptoms of rheumatoid arthritis patients. In recent years, various herbal-derived extracts and phytochemicals have been found to modulate the production of pro-inflammatory mediators. Thus, herbal-derived inhibitors of these mediators are highly appreciated as potential candidates for development of new anti-inflammatory drugs with reduced side effects. For instance, several flavonoids, such as amoradicin and genistein were found to block TNF-α production from LPS-treated cells from the macrophage cell line (RAW 264.7) [25]. Baicalin was found to inhibit the production of IL-1, IL-6, TNF-α, IFN-γ, monocyte chemotactic protein-1, macrophage inflammatory protein (MIP)-1, and MIP-1 at protein as well as at RNA levels in LPS-activated human blood monocytes [26]. Results obtained here show that HT-extracts inhibit the LPS-induced TNF-α and IL-6 production by down-regulating the gene expression of TNF-β and IL-6. Similar results were seen with various medicinal plants. For instance, feverfew extracts were found to inhibit the LPS-induced CCL2 (MCP-1) and TNF-α production by cells from the THP-1 line [27]. Moutan Cortex extracts were found to exhibit anti-inflammatory properties via the reduction of iNOS and COX-2 expression as well as through the suppression of the phosphorylation of I-kBa and the activation of NF-kB. Uncaria tomentosa extracts inhibited the MAP kinase-signaling pathway and altered cytokine expression in THP-1-derived macrophages cell line [28]. In another study, HT-extracts were found to inhibit the production of LPS-induced TNF-α production by down regulating the transcription of the TNF-α gene in LPS-activated THP-1-derived macrophages [15].
Treatment of PBMNCs with HT-extract remarkably increased levels of IL-10 protein and mRNA expression. These effects were evident at a significantly lower extract concentrations (125 µg/ml) compared to maximal inhibitory concentrations of IL-6 and TNF-α (250 µg/ml). IL-10 is a potent immune mediator, the primary physiological role is to reduce and terminate the ongoing inflammatory process [29,30]. LPS treatment of PBMNCs cells induced the secretion of a low level of IL-10 (65 pg/ml). Treatment with HT-extract increased IL-10 by two folds. Previous reports have clearly shown the antagonist effect of IL-10 on the secretion of pro-inflammatory cytokines [31,32], suggesting that HT-extract-mediated inhibition of the LPS-induced secretion and mRNA expression of IL-6 and TNF-α may pass through the induction of IL-10 production. Several inflammatory diseases share the dual characteristic of a very low blood level of IL-10 and a high blood level of TNF-α. Furthermore, injection of the recombinant form of IL-10 decreased the blood concentrations of TNF-α that has proven beneficial for such diseases [31-33]. The ability of HT-extract to modulate both, the pro-inflammatory and anti-inflammatory cytokines in LPS-activated PBMNCs represents an additional argument for the suggestion that it is an alternative or a complement that may help in the treatment and/or prevention of inflammatory diseases. The finding that the expressions of pro-inflammatory and the anti-inflammatory cytokines are affected at different HT-extract concentrations implies further detailed investigations.
HT-extract exhibited strong antioxidant activity as measured with the DPPH radical scavenging activity assay (IC 50 of 5 µg/ml), which is equal to that of vitamin C (5.1 µg/ml). Typically, inflamed tissues are associated with an elevated level of reactive species (reactive oxygen species (ROS)/reactive nitrogen species (RNS). These ROS/RNS are generated during the respiratory burst of immune cells and are important factors in defense against invading pathogens. It has been suggested that herbal-based antioxidants would be beneficial to human health and a lot of interest is focused on the determination of antioxidant capacity of natural products. Indeed, HT-extracts were found to inhibit the production of NO at the transcription level of iNOS gene [15], which was totally inhibited at non-toxic concentration of 250 µg HT-extract/ml. Similar results were found with other herbal-derived factors (eg.: flavonoids). Using LPS/cytokine-treated primary isolated macrophages or cell lines, varieties of flavonoids including apigenin, luteolin, and quercetin were shown to inhibit NO production. NO is a free radical synthetized from L-arginine with the help of nitric oxide synthases (NOSs). It is also an essential second messenger in large number of signal transduction of physiological and pathological processes [34]. At adequate levels, NO can generate or modify intracellular signals, thereby affecting the function of immune cells, tumor cells, and resident cells of different tissues and organs. However, its uncontrolled release can cause inflammatory damage of target tissue. A significantly increased concentartions of NO synthesized by the inducible form of nitric oxide synthase (iNOS) activate inflammatory process and acts synergistically with other inflammatory cytokines. Inhibition of iNOS activity or down-regulation of iNOS expression may be beneficial to reduce the inflammatory response. Therefore, it is meaningful to assess the role of NOS in the observed anti-inflammatory effects of Hypericum triquetrifolium [31].
These results indicate that HT-extract seems to exert anti-inflammatory properties through increasing the production of IL-10 at protein and mRNA levels as well as by suppressing the production of IL-6 and TNF-α at protein and gene expression levels in PBMNCs.
The authors would like to acknowledge Al-Qasemi Research Foundation and the Arab American University Research Foundation for providing their financial support.