Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2013) Volume 3, Issue 1
Keywords: Pythium, Oomycetes, Paromomycin, Hemocytometer,MTT, Viability
Pythium myriotylum Drechsler, an economically important cosmopolitan oomycetous phytopathogen, is the causal agent of soft rot, a devastating soil-borne disease causing significant losses in the annual yield of a variety of crops [1]. The necrotrophic pathogen is capable of destroying whole crops within a single season, and contributing to severe epidemics. The pathogen’s infective potential, as well its spatial dissemination relies on rapid and copious production of asexual motile zoospores [2]. Adhesion of zoospores to plant’s root surface is an important early event in plant-oomycete interactions [3]. Zoospores lack cell wall and quickly differentiate to form cysts that germinate producing hyphae, and finally form mycelial mass that helps in host colonization [4,5]. Disease control is usually achieved by the use of chemical pesticides, especially metalaxyl/phosphonate treatment that are inadvertently associated with development of fungicide resistance [6-8]. Earlier studies have suggested the use of biocontrol agents as a good alternative to chemical fungicides [9], with Streptomyces and Bacillus suggested as potential BCAs against Pythium spp [10,11]. Paromomycin produced by Streptomyces spp. has been identified to exhibit modest activity against Pythium spp. Paromomycin is known to inhibit both bacterial and eukaryotic ribosomal protein synthesis, with eukaryotes being 10 to 15 times less sensitive than prokaryotes [12]. Oomycetes comprising Pythium spp. are eukaryotic organisms, closely related to heterkonts than fungi [13,14). An epidemic caused by P. myriotylum greatly impacts agricultural production and related economics, and also since the mechanistic basis for unusual sensitivity of Pythium spp. to paromomycin is relatively unknown, the rationale of the present study constituted examining the mechanistic aspect of the inhibitory effect of paromomycin on P. myriotylum, on both zoospores and hyphae.
P. myriotylum zoospore production
Zoospores of P. myriotylum were produced from seven to ten day old cultures grown on Potato Dextrose Agar (PDA) plates at 25°C by the methods of Rahimian et al. [15] and Pacumbaba et al. [16] with modifications. Mycelial discs (5 mm diameter) were transferred to petri dish containing sterile water (pH-7.0), and placed under fluorescent light at room temperature for 24 h to induce zoospore release. Zoospore concentrations were estimated with a Neubauer chamber under bright field microscope (Olympus BX51, Japan), at 40X magnification.
Microscopic evaluation to determine inhibitory effect of paromomycin on P. myriotylum zoospores
Different concentration of paromomycin (5, 10, 15, 20, 25, 50, 75, 100 μg/ml) was added to zoospores (107/ml), and incubated overnight at 25°C. Spores were counted on a haemocytometer under bright field microscope (Olympus BX51, Japan) at 40X, by taking mean value of counts measured from 15 hemacytometer squares per slide. Experiments were carried out in triplicate and data were expressed as mean ± SEM, of at least three independent experiments. One-way analysis of variance (ANOVA) and Tukey’s HSD test were carried out to test any significant differences among the means, with hypothesis testing at P ≤ 0.05.
Determination of biomass to assess P. myriotylum mycelial growth rate
Mycelial growth of P. myriotylum in liquid cultures, in presence of paromomycin, was assessed by inoculation of mycelial disc (5 mm) in Czepek Dox minimal media (30 g Sucrose, 2 g NaNO3, 1 g KH2PO4, 0.5 KCl, 0.5 g MgSO4.7H2O, 0.1 g FeSO4.7 H2O per liter), with trace element solution (1g ZnSO4, 0.5 g CuSO4.5H2O per liter). After incubation at 25°C for 5 days, mycelial biomass from triplicate samples containing varying concentrations of paromomycin was collected on pre-weighed filter papers (Whatman No. 1). Dry weight yield was determined after 8 h oven drying at 60°C, and percent loss in mycelial dry weight was calculated over untreated control.
Permeability changes in P. myriotylum hyphal membrane
Mycelial plug (5 mm diameter) taken from 7-day old culture of P. myriotylum grown in PDA, were inoculated in Czepek Dox broth (3 mL) and incubated at 25°C for 5 days. Mycelia were harvested and placed in 1X phosphate buffered saline (pH-7.2), containing different concentrations of paromomycin (5, 10, 25, 50, 75, 100 μg). After incubation at 25°C for different time periods ranging from 1-6 hours, the supernatant was filtered using 0.22 μm filter discs (Millipore) and UV absorbing materials, in each of the treated and control samples were measured at 260 nm, using UV-visible spectrophotometer (UV3000, LabIndia).
Hyphal morphology analysis
To study morphological alterations induced by paromomycin, P. myriotylum mycelial disc (5 mm diameter) grown on Czepek Dox minimal media were harvested and treated with paromomycin (25 μg/ml) for 2 h at 25°C. Observations were done under Light microscope (Olympus BS60, Japan) at 40X magnification. For SEM measurements, thin layers (1 mm) of agar blocks containing actively growing mycelium were cut from growing edges of PDA plates. Mycelial discs were treated with paromomycin at sub-inhibitory concentration (25 μg/ml). Treated and untreated mycelia were adhered to polylysine coated glass cover slips, and fixed with glutaraldehyde (2.5%) in 0.1M phosphate buffer (pH-7.5) for 3 hours at 25°C. After fixation, they were washed in the same buffer and dehydrated in a graded ethanol series (30%, 50%, 70%, 90%, 95%, and two times at 100%), for a period of 10 min in each series [17]. The fixed material was mounted on stubs using double- sided carbon tape, and coated with gold in a sputter coater system (E-1010-ion sputter, Hitachi) for 20 seconds at 15 mA. The SEM observations were examined and digital images captured using a S06600SEM (Hitachi), at an accelerating voltage of 5 kV.
The infective potential of P. myriotylum, as well its spatial dissemination, relies on asexual zoospore production. The asexual zoospores of Pythium spp. have been reported to initiate infection after attachment to the host surface [18], and mediate epidemic development of soft rot disease. Thus, understanding the mode of action of agents that reduce the spore viability/germination constitutes key elements for effective pathogen control. Paromomycin, an aminoglycoside antiobiotic isolated from Streptomyces rimosus ssp. paromomycinus [19] has previously been demonstrated to be effective against Phytophthora and Pythium species [20]. Present study provides a mechanistic basis for understanding the anti-Pythium activity of paromomycin, in view of the limited information available on the mode of inhibitory action. Microscopic examination of P. myriotylum zoospores using haemocytometer revealed a concentration-dependent reduction in zoospore number, with increasing paromomycin concentration (Table 1). The mode of action of paromomycin and various other prokaryotic inhibitory aminoglycoside antibiotics against eukaryotic phytopathogens like Phytophthora and Pythium species is still not well-defined [20]. Like macrolide antibiotics [21], mechanism of action of paromomycin on spores of P. myriotylum could be through compromising membrane permeability, allowing easy access of the antibiotic to sensitive intracellular sites, that includes the mitochondrial and cytoplasmic protein synthetic systems [21].
| Paromomycin concentration | Zoospore lysisa (in %) |
|---|---|
| Hemocytometer count | |
| 5 µg | 44.33 ± 2.08 |
| 10 µg | 36.26 ± 1.07 |
| 15 µg | 31.66 ± 1.52 |
| 20 µg | 23.83 ± 2.30 |
| 25 µg | 13.83 ± 2.30 |
| 50 µg | 9.75 ± 1.08 |
| 75 µg | 7.73 ± 0.25 |
| 100 µg | 6.55 ± 0.18 |
a: % of zoospore lysis ± SE, calculated by dividing number of zoospores counted after treatment with paromomycin to the number of zoospores in control sample, and expressed as percentage.
Table 1: Lytic activity of paromomycin on P. myriotylum zoospores. Results expressed as mean ± standard deviation.
Inhibition of hyphal growth by paromomycin accounts for the observed dose-dependent decline in mycelial biomass (Figure 1). If the observed reduction in mycelial biomass by paromomycin involves compromising the integrity of hyphal membrane, then it will result in leakage of cytosolic elements into the extracellular milieu [22]. This was further monitored by measuring the release of UV absorbing cellular materials, after exposure to paromomycin. We observed leakage of cellular contents in a time and dose-dependent manner (Figure 2). Further evaluation of morphological damages to the hyphae under light microscope (Figure 3a and b), and subsequently with SEM (Figure 3c and d) revealed several morphological alterations, such as hyphal shrinkage and cytoplasmic coagulation, within 2 h of treatment with paromomycin (Figure 3b), resulting in severely collapsed and squashed hyphae (Figure 3d), whereas healthy and continuous hyphae were observed in control sample (Figure 3a and c). Similar effects of essential oils on the hyphal morphology of the plant pathogenic fungi have also been reported by other authors [23,24]. Collectively, our findings suggest that the anti-Pythium effects of paromomycin, which is a lipophilic aminoglycoside, are due to damage to hyphal membrane by altering the fluidity and permeability of cell membranes.
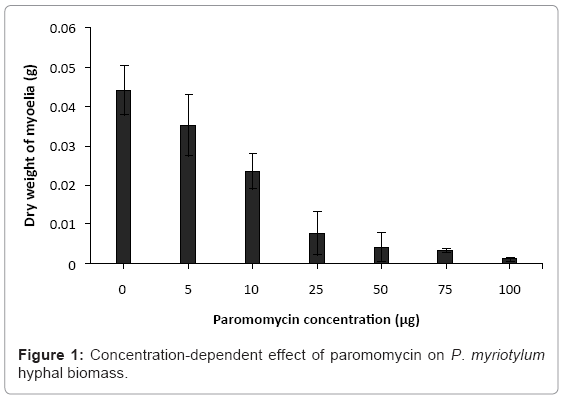
Figure 1: Concentration-dependent effect of paromomycin on P. myriotylum hyphal biomass.
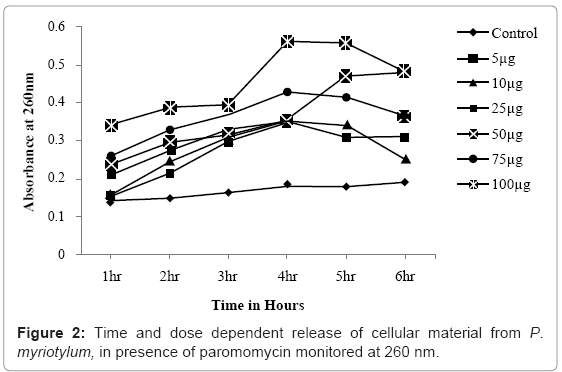
Figure 2: Time and dose dependent release of cellular material from P. myriotylum, in presence of paromomycin monitored at 260 nm.
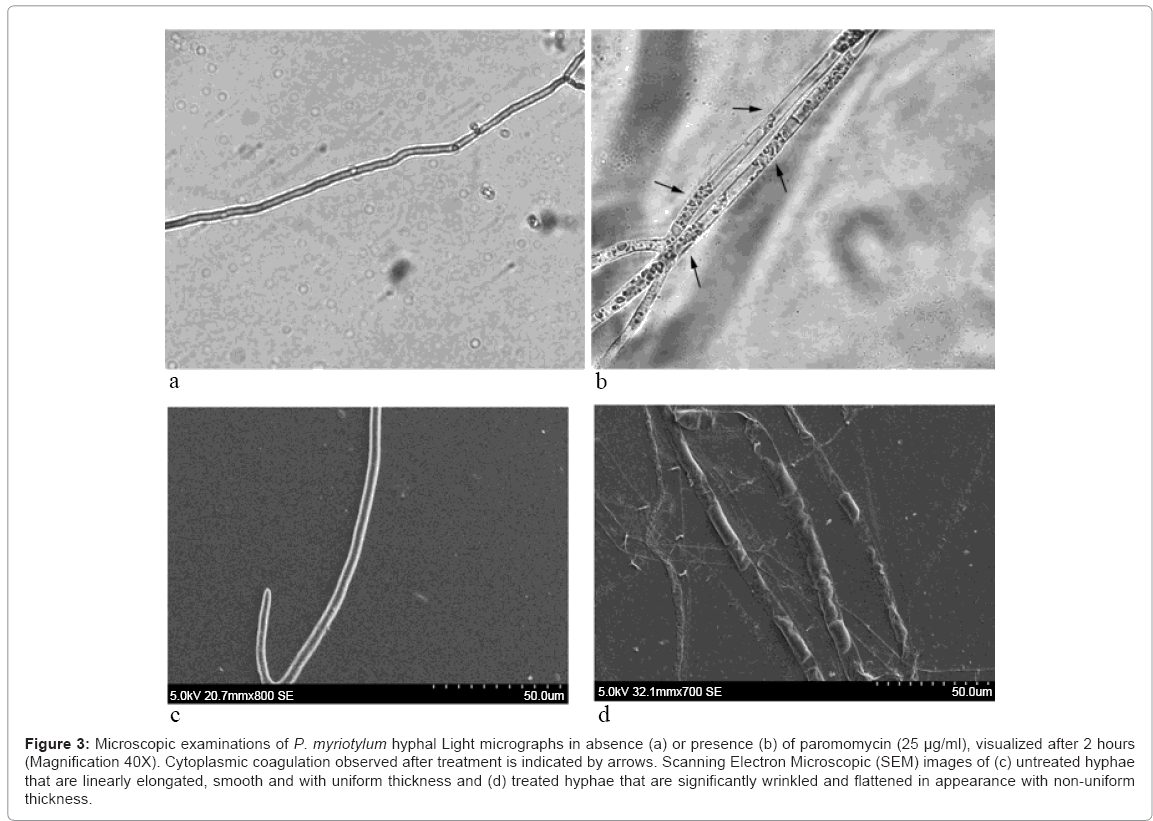
Figure 3: Microscopic examinations of P. myriotylum hyphal Light micrographs in absence (a) or presence (b) of paromomycin (25 μg/ml), visualized after 2 hours (Magnification 40X). Cytoplasmic coagulation observed after treatment is indicated by arrows. Scanning Electron Microscopic (SEM) images of (c) untreated hyphae that are linearly elongated, smooth and with uniform thickness and (d) treated hyphae that are significantly wrinkled and flattened in appearance with non-uniform thickness.
We gratefully acknowledge financial support provided under Faculty Research Grant (FRG) scheme (No. NITC/Dean (C&SR)/ FRG10/0112) of National Institute of Technology Calicut.