Drug Designing: Open Access
Open Access
ISSN: 2169-0138
ISSN: 2169-0138
Research Article - (2016) Volume 5, Issue 1
Dracunculiasis caused by the pathogen Dracunculus medinensis (a little dragon from Medina). The Cyclops is the intermediate host that ingests the larvae of parasite (D. medinensis), that later on ingested by the human from the contaminated stagnant unfiltered water from the water source. After an incubation period of a year these mature female worm comes towards the skin and start formation of a small round bulge on the skin by secreting an irritating chemical which causes severe pain. In this study we will summarize the potency of NADH dehydrogenase subunit 5 (mitochondrion) from Dracunculus medinensis with 527 amino acids. Antigenic peptide of NADHdehydrogenasesubunit5 protein is most suitable for subunit vaccine development because with single epitope, the immune response can be generated in large population. In this research, we used PSSM and SVM algorithms for the prediction of MHC class I and II binding peptide, antigenicity, Solvent accessibility, polar and nonpolar residue to analyse the regions that are likely exposed on the surface of proteins which are potentially antigenic that allows potential drug targets to identify active sites against infection as well as to design effective drug to treat it.
<Keywords: Dracunculiasis, Antigenic peptides; MHC-Binders; TapPred; PSSM; SVM; Nonamers; NADH; Dehydrogenase subunit 5
NADH dehydrogenase 5 (ND5) protein contribute to be a part of the large enzyme complex i.e Complex I, which is active in mitochondria. Complex I is an one of the essential enzyme complexes necessary for oxidative phosphorylation and thereafter, mitochondrial enzyme complexes execute chemical reactions that contribute to the ATP production. Complex I is responsible for the first step in the electron transport process. The NADH dehydrogenase complex (or mitochondrial respiratory complex I) plays an important role in the oxidation of NADH by ubiquinone by catalysis. This chemical reaction is linked to proton transfer across mitochondrial membranes. Study reveals that there are multiple humans disorders are consociated with mutations of the mitochondrial ND genes [1]. Mutations in mitochondrial DNA of ND5 have been found to be the most important cause of several disorder like hypertrophic cardiomyopathy(homoplasmic ND5 12338T>C variant) [2], Leber’s hereditary optic neuropathy (LHON) which is a maternally inherited eye disease(homoplasmic ND4 G11696A and ND5 T12338C mutation) [3], Light syndrome(ND5*13513 G to A mutation) [4], Leber’s hereditary optic neuropathy is associated with the T12338C mutation in mitochondrial ND5 gene in six Han Chinese families [5]. The heteroplasmic T>C transition at position 13271 in MTND5 affects a highly conserved base and segregates with the disease. Investigation suggest that, this is the 15th mutation affecting the MTND5 subunit of respiratory chain complex I and affirms its importance site for disease with phenotypes ranging from MELAS and infantile encephalopathies to isolated syndromes affecting a single tissue such as Leber hereditary optic neuropathy and now skeletal muscle [6]. Looking at its important we have used this protein to study the antigenicity of protein, its solvent accessibility, polar and nonpolar residue to analyse the regions that are likely exposed on the surface of proteins which could be the potentially antigenic that allows potential drug targets to identify active sites against infection as well as to design effective drug to treat Dracunculiasis. D.medinesis (a little dragon from Medina) is the causative agent and it is the only species from the 12 species of Dracunculus [7-10] which infects humans, commonly known as “Guinea Worm Disease (GWD)”. The other Dracunculus species generally resides in the internal tissues and body cavities of non-human mammals and reptiles (snake and turtles) [11]. This little dragon undergo a very unusual life cycle of six developmental stages with incubation period last for 1 to one an half years approximately (Figure 1) [12]. This is one of the most neglected tropicalparasites which bears clinical importance and needs to be eradicated after small pox [13]. After reaching to the maturation stage, these worms copulate and an adult female produces millions of eggs in its uterus whereas male dies. Later on, the female worm release the larvae which induces a painful blister (1-6 cm diameter ) on the skin of lower limbs (predominantly localized in the lower extremities(80-90%) in most of the reported cases) (Figure 1a). The infected person develops slight fever, local skin redness, swelling and severe pruritus around the blister. Other symptoms include: diarrhea, nausea, vomiting and dizziness. The blister burst within three days and female worms one or more slowly comes out from the wounds which causes an excoriating burning sensation and pain [14] (Figure 1b). Immersing or pouring water over the blister provides pain reliever. But this the moment that adult female is exposed to the external environment [15] During emergence of the limbs in open water sources it recognizes the temperature difference and releases the milky white liquid in the water which contains millions of immature larvae, when larvae released in water are ingested by copepods where they mount twice and become infective larvae within two weeks [16].
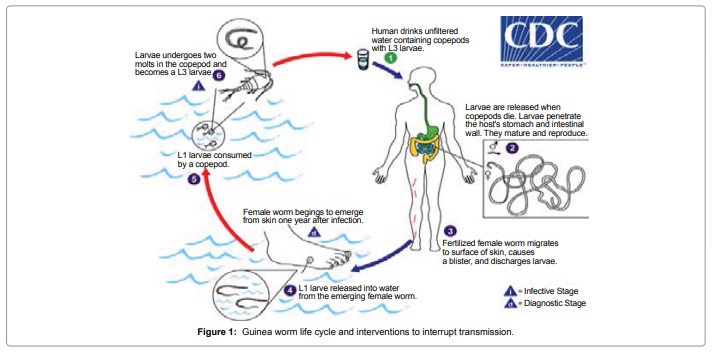
Figure 1: Guinea worm life cycle and interventions to interrupt transmission.
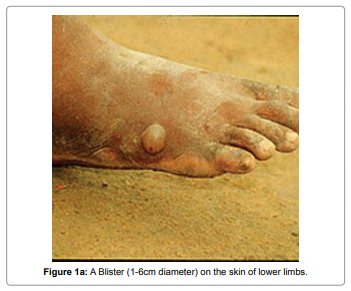
Figure 1a: A Blister (1-6cm diameter) on the skin of lower limbs.
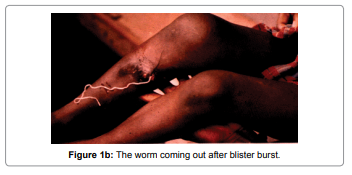
Figure 1b: The worm coming out after blister burst.
The D. medinensis antigen peptides can be most desirable segment for the subunit vaccine development because with the single epitope, the immune response can be generated in large population. This approach is usually based on the phenomenon of cross-protection, whereby infected with the mild strain and is protected against a more severe strain of the same. The phenotype of the resistant transgenic hosts includes fewer centers of initial infection, a delay in symptom development and low accumulation. The World Health Assembly called in the 1986 in order to dracunculiasis elimination. The global Guinea Worm Eradication Program, supported by The Carter Center, World Health Organization (WHO), UNICEF, CDC, and other partners, began assisting ministries of health of countries in which dracunculiasis is endemic in meeting this goal. At that time, an estimated 3.5 million cases occurred each year in 20 countries in Africa and Asia. Dracunculiasis remains endemic in four countries in 2014 (South Sudan, Chad, Mali, and Ethiopia), but the number of overall reported incidence is decreases in 2013 by 73% and in 2014 by 71% compared with 2012. Failures in surveillance and containment, lack of clean drinking water, insecurity in Mali and parts of South Sudan, and an unusual epidemiologic pattern in Chad are the main remaining challenges to dracunculiasis eradication [17]. A case of Onchocerca volvulus has been reported in the Cameroon which is mimicking Dracunculus medinensis [18]. More than two decades after the International Drinking Water Supply and Sanitation Decade (IDWSSD) implemented by the United Nations (1981-1990) [19], the disease still lingers, underscoring the daunting challenge of disease control, as has been the case of the failure of previous attempts to eradicate diseases like malaria, hookworm and yaws [20]. Till date there is no accurate and efficient curative drug of vaccine is available against dracunculiasis [21]. The investigation suggests that the immunity is not developed by the infected individual [22]. Dracunculus medinensis –specific antibodies (total, IgG1 and IgG4) during the time of patency, which were significantly higher than the levels measured in the same individuals eight months later, except for a few individuals who had developed a new patent infection [23]. It was observed that the mean level of specific IgG1 and IgG4 is higher during the month of potency of the infected individual whereas variation in the IgE value is relatively negotiable and constant before, during the infection and after the recovery. There is possibility that variation in antibody production is regulated by infected larvae (i.e by transmission ) and / or by adult worms (i.e by patency) is still need to be clear out. It is possibility that increased production of IgG1,IgG4 during the time of patency plays a role in blockingor protecting immune responses otherwise it could have killed ingested infected larvae [24]. The D. medinensis antigen peptides can be most desirable segment for the subunit vaccine development because with the single epitope, the immune response can be generated in large population. This approach is usually based on the phenomenon of cross-protection, whereby infected with the mild strain and is protected against a more severe strain of the same. The resistant transgenic host’s phenotype includes of fewer centers of initial infection, following a delay development in symptom with low accumulation. Antigenic peptides from D. medinens is most suitable for the development of peptide vaccine [25] because a single protein subunit can generate sufficient immune response. In this research work we have used the phenomenon of cross-protection, whereby an individual undertaken by a mild toxin can have immunity to survive against similar strong toxic effects. MHC molecules are cell surface protein that binds to the peptides derived from host or antigenic proteins and present them to cell surface for recognition by T-cells. T cell recognition is an important mechanism of the adaptive immune system by which the host identifies and responds to foreign antigens [25,26]. There are two types of MHC molecule and are extremely polymorphic. MHC class I molecules present peptides from proteins synthesized within the cell, whereas, MHC class II molecule present peptides derived from endocytosed extracellular proteins. MHC molecules have been well characterized due to their role in immune reactions and they take active part in host immune reactions and involvement of MHC class molecule in response to almost all antigens and it give impacts on specific sites. The involvement of MHC class-I molecule in response to almost all antigens make the study very interesting. They bind to some of the peptide fragments generated after proteolytic cleavage of antigen [27]. Identification of MHC-binding peptides and T-cell epitopes helps improve our understanding of specificity of immune responses [28]. Antigenic peptides are most suitable for peptide vaccine development because single epitope can generate large the immune response [29].
Database searching
The antigenic protein sequence of NADH dehydrogenase subunit5 from Dracunculus medinensis was retrieved from www.ncbi.nlm.nih. gov, UniProt databases are initially the most important [30-32].
Prediction of antigenicity
Prediction of antigenicity program predicts those segments from Antigen NADHdehydrogenasesubunit5 protein that are likely to be antigenic by eliciting an antibody response. In this research work antigenic epitopes of Dracunculus medinensis antigen NADHdehydrogenasesubunit5 are determined by using the Hopp and Woods, Welling, Parker, Bepipred ,Kolaskar and Tongaonkar antigenicity methods [33-38].
Prediction of MHC binding peptide
The major histocompatibility complex (MHC) peptide binding of Dracunculus medinensisis predicted using neural networks trained on C terminals of known epitopes. Rankpep predicts peptide binders to MHC-I ligands whose C-terminal end is likely to be the result of proteosomal cleavage using Position Specific Scoring Matrices (PSSMs). Peptides that bind to a given MHC molecule share sequence similarity. Traditionally, the sequence patterns used for the prediction of peptides binding to MHC molecules. Such sequence patterns, however, have proven to be too simple, as the complexity of the binding motif cannot be precisely represented by the few residues present in the pattern [39]. RANKPEP uses “Position Specific Scoring Matrices (PSSMs) or profiles” from set of aligned peptides known to bind to a given MHC molecule as the predictor of MHC-peptide binding and overcome the complexity of the binding motif limitation. RANKPEP web server is a variability masking feature to focus on the prediction of conserved epitopes, which could thus help to avoid immune evasion resulting from mutation. Support Vector Machine (SVM) based method for prediction of promiscuous MHC class II binding peptides from protein sequence; SVM has been trained on the binary input of single amino acid sequence [40-43].
Prediction of antigenic peptides by Cascade SVM based TAPPred method
In the present study, we predict cascade SVM based several TAP binders which was based on the sequence and the features of amino acids [44]. We found the MHCI binding regions (Table 1), the binding affinity of Dracunculus medinensis.
| MHC-I Allele | RANK | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|---|
| 8mer_H2_Db | 1 | 262 | STL | SQIGFCFF | GLG | 930.1 | 20.659 | 39.35% |
| 8mer_H2_Db | 7 | 63 | GSV | VVYSGFYM | MED | 947.12 | 13.569 | 25.85% |
| 8mer_H2_Db | 8 | 306 | LCG | GQQDSRGY | VGV | 891.9 | 12.539 | 23.89% |
| 8mer_H2_Db | 9 | 512 | FSG | KEINFLFL | CLF | 1005.23 | 12.34 | 23.51% |
| 8mer_H2_Db | 10 | 419 | LWW | LNYNSFVI | SSS | 951.09 | 12.251 | 23.34% |
| 9mer_H2_Db | 1 | 418 | FLW | WLNYNSFVI | SSS | 1114.3 | 22.359 | 44.39% |
| 9mer_H2_Db | 2 | 71 | FYM | MEDYNFNYF | CVV | 1224.32 | 20.942 | 41.58% |
| 9mer_H2_Db | 3 | 95 | GVV | FSNNCISML | IFW | 1010.19 | 19.364 | 38.45% |
| 9mer_H2_Db | 4 | 332 | VSL | MCLCGLFFL | GGS | 1028.36 | 18.362 | 36.46% |
| 9mer_H2_Db | 6 | 101 | NCI | SMLIFWDLL | GVS | 1096.39 | 16.749 | 33.26% |
| 10mer_H2_Db | 1 | 179 | FTK | SAQYP FSSWL | PKA | 1144.3 | 29.252 | 49.70% |
| 10mer_H2_Db | 4 | 109 | WDL | LGVSSYFLVL | YYG | 1079.31 | 12.086 | 20.53% |
| 10mer_H2_Db | 6 | 305 | HLC | GGQQDSRGYV | GVG | 1048.08 | 9.329 | 15.85% |
| 10mer_H2_Db | 8 | 395 | LFY | SGGSLVFSFV | SLV | 981.12 | 8.591 | 14.60% |
| 10mer_H2_Db | 9 | 433 | LLY | SDYYSLFYLF | ILG | 1299.47 | 8.398 | 14.27% |
| 11mer_H2_Db | 3 | 263 | TLS | QIGFCFFGLGL | GLV | 1183.44 | 16.546 | 20.81% |
| 11mer_H2_Db | 5 | 61 | VVG | SVVVYSGFYMM | EDY | 1264.52 | 15.559 | 19.57% |
| 11mer_H2_Db | 6 | 368 | LFF | FSILLTYLYCY | RLM | 1380.68 | 13.326 | 16.76% |
| 11mer_H2_Db | 7 | 261 | LST | LSQIGFCFFGL | GLG | 1213.47 | 12.853 | 16.17% |
| 11mer_H2_Db | 8 | 93 | MVG | VVFSNNCISML | IFW | 1208.45 | 12.594 | 15.84% |
Table 1: Promiscuous MHC ligands, having C-terminal ends are proteosomal cleavage sites of Dracunculus medinensis. The antigenic peptide to the MHC-1 Allele i.e. 8mer_H2_Db(The binding thresholds:33.04, optimal score:52.494), 9mer_H2_Db(Optimal Score: 50.365, Binding Threshold: 17.96), 10mer_H2_Db(The Optimal Score: 58.858, BindingThreshold: 41.32),11mer_H2_Db(Optimal Score: 79.495, Binding Threshold: 56.96). (All rows highlighted in red represent predicted binders and A peptide highlighted in violet has a C-teminus predicted by the cleavage model used).
Solvent accessible regions
We also analyzed the solvent accessible regions of proteins having highest probability that a given protein region lies on the surface of a protein Surface Accessibility, backbone or chain flexibility by Emini et al. [45] and Karplus and Schulz [46]. By using different scale we predict the hydrophobic and hydrophilic characteristics of amino acids that are rich in charged and polar residues [47-56].
The Dracunculus medinensis Antigen NADH dehydrogenase subunit 5, contain a long residue of 527 amino acids with 483 nonamers.
MDVYIYWFGVILLCFLLLFVFFYDDFVFSLDFSSLELLQFQFRLDWFSFSFFCLLVMVVGSVVVYSGFYM
MEDYNFNYFCVVLSIFVFSMVGVVFSNNCISMLIFWDLLGVSSYFLVLYYGNWDSCSGSMNTVMMNRVGDVCVFLVFCGLFFMGIDFISLELVVSVALFFFILSTFTKSAQYPFSSWLPKAMSAP
TPVSALVHSSTLVTAGLFLGMCFSEVMFLDFVLDFMFFVGLFTMFSSGLMAYFEFDIKKLVALSTLSQIGFCFFGLGLGLVYFSFIHMLSHAVFKSCLFMQMGYIIHLCGGQQDSRGYVGVGGLS SVVYIQTFVSLMCLCGLFFLGGSVSKEILLEHYFFCNWSLFLVFLFFFSILLTYLYCYRLMKGFYYYCSSSLFYSGGSLVFSFVSLVLVVFSIVFLWWLN
YNSFVISSSLLYSDYYSLFYLFILGLVLCVVFFKFGSFDVKYKFYGDLLPKVIIRGNYVVKWSDCMVDYSIMKFGDFSFYVSKIFVMGFSGKEINFLFLCLFLLLMI
Prediction of antigenic peptides
In this study, we found the antigenic determinants by finding the area of greatest local hydrophilicity. The Hopp-Woods scale Hydrophilicity Prediction Result Data found high in position between300-320 in a protein, assuming that the antigenic determinants would be exposed on the surface of the protein and thus would be located in hydrophilic regions (Figure 2). Welling antigenicity plot gives value as the log of the quotient between percentage in a sample of known antigenic regions and percentage in average proteins and Prediction Result Data found high in position 250-270 (Figure 3). We also study Hydrophobicity plot of HPLC / Parker Hydrophilicity Prediction Result Data found between 300-320 (Maximum Score-6.3) i.e., the maximum predicted residues at the position 308(Residue is Q) is 305- GGQQDSR -311and 309 (Residue D) is 306- GQQDSRG-312 (Figure 4), BepiPred predicts the location of linear B-cell epitopes Result found that between 300-310 is 301-MQMGYIIHLC-310 and the maximum score (1.369) is found at the position 308,309 (Figure 5), Kolaskar and Tongaonkar antigenicity methods (Figure 6) Predicted peptides result found i.e.,
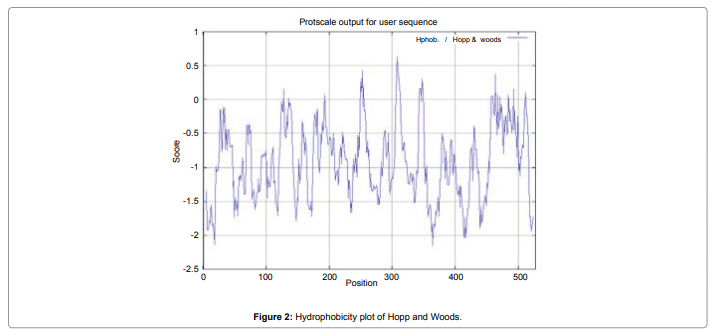
Figure 2: Hydrophobicity plot of Hopp and Woods.
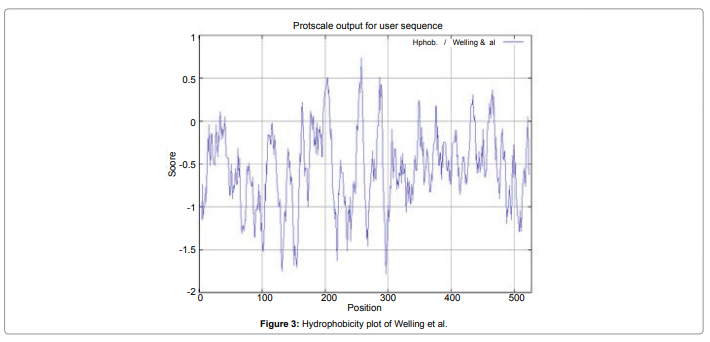
Figure 3: Hydrophobicity plot of Welling et al.
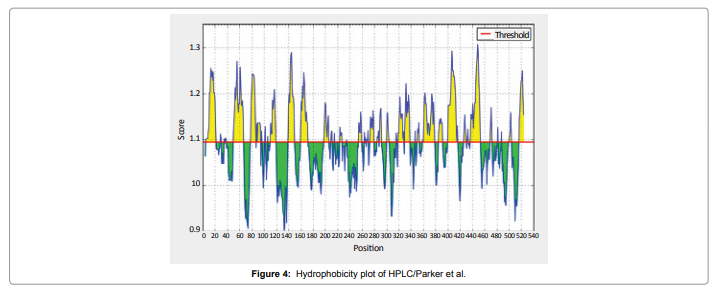
Figure 4: Hydrophobicity plot of HPLC/Parker et al.
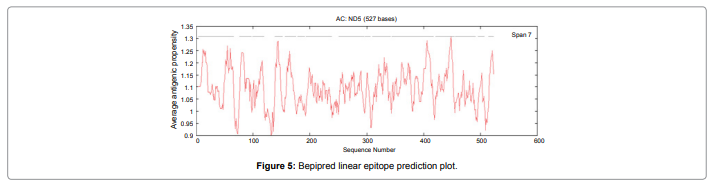
Figure 5: Bepipred linear epitope prediction plot.
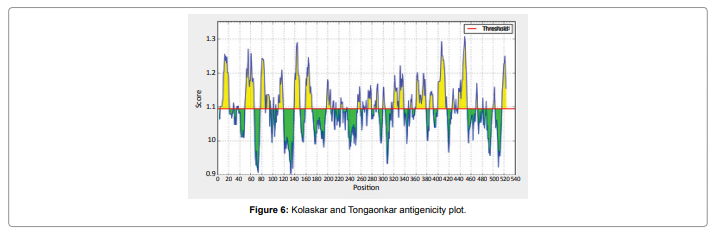
Figure 6: Kolaskar and Tongaonkar antigenicity plot.
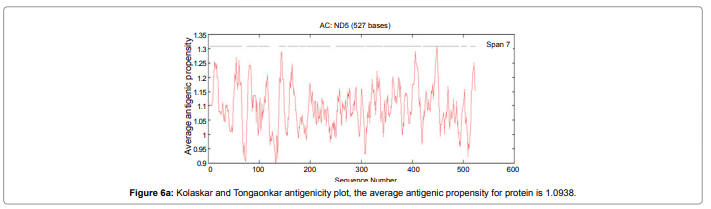
Figure 6a: Kolaskar and Tongaonkar antigenicity plot, the average antigenic propensity for protein is 1.0938.
4-YIYWFGVILLCFLLLFVFFYDDFVFSLDFSSLELLQFQFRLDWFSFSFFCLLVMVVGSVVVYS- 66
76-FNYFCVVLSIFVFSMVGVVFSN-97
99-CISMLIFWDLLGVSSYFLVLY-119
138-VGDVCVFLVFCGLFF-152
156-DFISLELVVSVALFFFILST-175
178-KSAQYPFSSWLPKA-191
193-SAPTPVSALVHSSTLVTAGLFLGMCFSEVMFLDFVLDFMFFVGLFT- 238
251-DIKKLVALSTLSQIGFCFFGLGLGLVYFSFIHMLSHAVFKSCLFM- 295
297-MGYIIHLCGG-306
310-SRGYVGVGGLSSVVYIQTFVSLMCLCGLFFLGG-342
344-VSKEILLEHYFFCNWSLFLVFLFFFSILLTYLYCYRLMKGFYYYCSSSLFYSGGSLVFSFVSLVLVVFSIVFLW- 417
421-YNSFVISSSLLYSDYYSLFYLFILGLVLCVVFFK-454
456-GSFDVKYKFYGDLLPKVIIRGNYVVKWSDCMVDYSI-491
496-DFSFYVSKIFVM-507
514-INFLFLCLFL-523 and the predicted antigenic fragments can bind to MHC molecule is the first bottlenecks in vaccine design.
Solvent accessible regions
We also predict solvent accessible regions in proteins; different measurement was performed for the prediction of antigenic activity, surface region of peptides. Emini et al. (Figure 7) predicts the highest probability i.e. found306-GQQDSR-311(maximum score 7.194), 307-QQDSRG-312(maximum score 7.194),308- QDSRGY- 313(score-6.508),432- YSDYYS-437(6.379),178- KSAQYP-183(score 6.281), that a given protein region lies on the surface of a protein and are used to identify antigenic determinants on the surface of proteins. Karplus and Schulz (Figure 8) High score is found i.e. found 1.122 maximum in 305- GGQQDSR-311. Predict backbone or chain flexibility on the basis of the known temperature B factors of the a-carbons. The hydrophobicity and hydrophilic characteristics of amino acids is determined by using different scales that are rich in charged and polar residues i.e. Sweet et al. hydrophobicity prediction Result Data found high in position 350-370 (Figure 9), Kyte and Doolittle result high in position 10-20 (Figure 10), Abraham and Leo result high in position 350-370(Figure 11), Bull and Breese result high in position 300-320 (Figure 12), Miyazawa result high in position 350-370 (Figure 13),Guy result high in position 300-320 (Figure 14), Wolfenden result high in position 440-460 (Figure 15), Roseman result high in position 350-380 (Figure 16),Wilson et 350-380 (Figure 17), Cowan 350-380 (Figure 18), Chothia 400-430 (Figure 19).
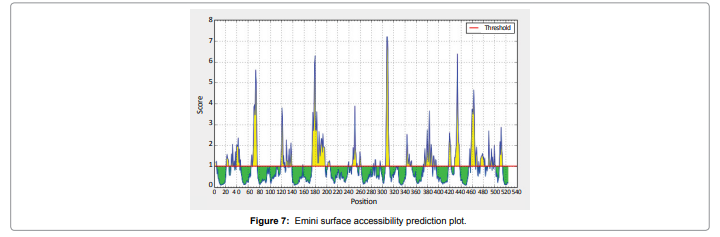
Figure 7: Emini surface accessibility prediction plot.
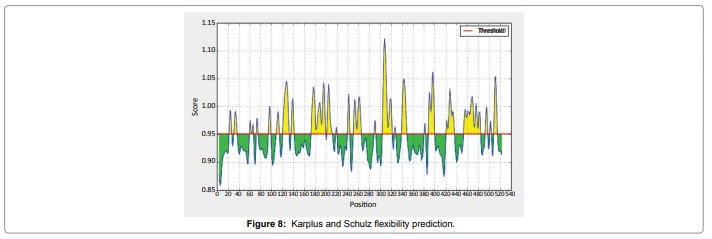
Figure 8: Karplus and Schulz flexibility prediction.
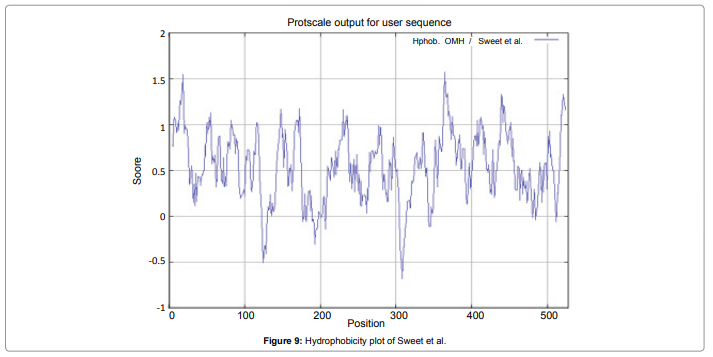
Figure 9: Hydrophobicity plot of Sweet et al.
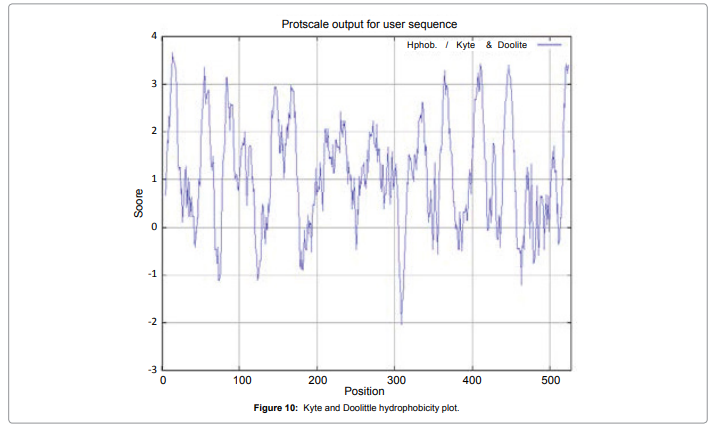
Figure 10: Kyte and Doolittle hydrophobicity plot.
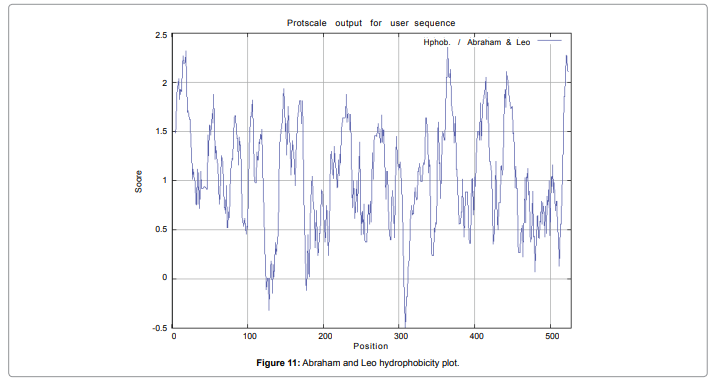
Figure 11: Abraham and Leo hydrophobicity plot.
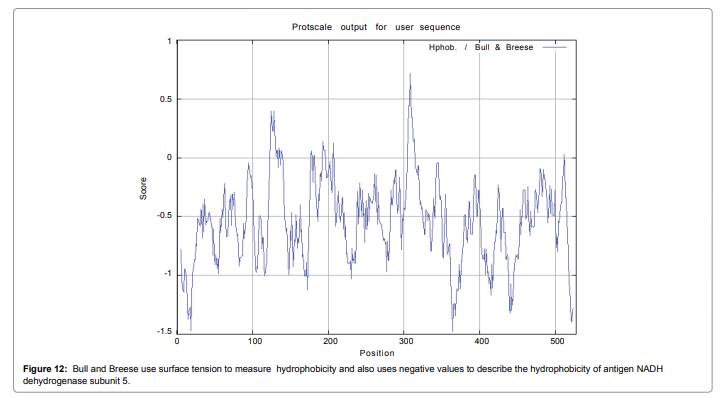
Figure 12: Bull and Breese use surface tension to measure hydrophobicity and also uses negative values to describe the hydrophobicity of antigen NADH dehydrogenase subunit 5.
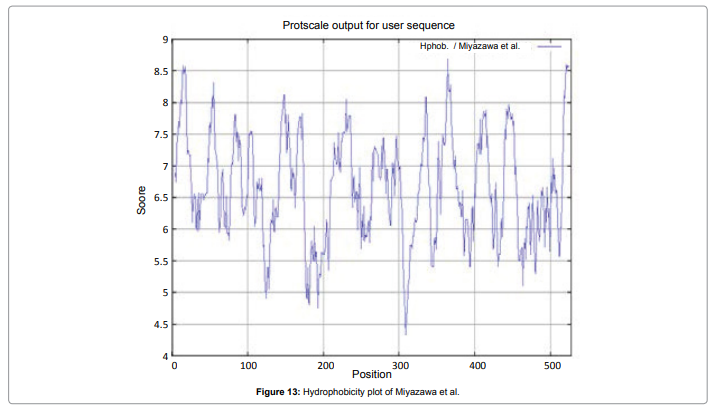
Figure 13: Hydrophobicity plot of Miyazawa et al.
Figure 14: Hydrophobicity plot of Guy.
Figure 15: Hydrophobicity plot of Wolfenden et al.
Figure 16: Hydrophobicity plot of Roseman.
Figure 17: Hydrophobicity/HPLC plot of Wilson.
Figure 18: Hydrophobicity/HPLC pH 3.4/ plot of Cowan.
Figure 19: Hydrophobicity plot of Chothia.
Prediction of MHC binding peptide
We found the binding of peptides to a number of different alleles using Position Specific Scoring Matrix. NADH dehydrogenase subunit 5 of Dracunculus medinensis antigen, with sequence 527 amino acid residues long, is having 519 nonamers. MHC molecules are cell surface proteins, which actively participate in host immune reactions and involvement of MHC-I and MHC-II in response to almost all antigens. We have predicted MHC-I peptide binders of NADH dehydrogenase subunit 5 from Dracunculus medinensis was tested with on a set of 4 different alleles i.e. H2-Db (mouse) 8mer, H2-Db (mouse) 9mer, H2- Db (mouse) 10mer and H2-Db (mouse) 11mer (Table 2) and MHC-II peptide binders for I_Ab.p, I_Ad.p,I_Ag7.p alleles highlighted in red represent predicted binders (Table 3). Here RANKPEP report PSSMspecific binding threshold and is obtained by scoring all the antigenic peptide sequences included in the alignment from which a profile is derived, and is defined as the score value that includes 85% of the peptides within the set. Peptides whose score is above the binding threshold will appear highlighted in red and peptides produced by the cleavage prediction model are highlighted in violet. We also use a cascade SVM based TAPPred method which found 80 High affinity TAP Transporter peptide regions which represents predicted TAP binders residues which occur at N and C termini from Dracunculus medinensis (NADHdehydrogenasesubunit5) (Table 3).
| Allele | RANK | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|---|
| MHC-II I_Ab | 1 | 385 | KGF | YYYCSSSLF | YSG | 1114.26 | 12.219 | 34.29% |
| MHC-II I_Ab | 2 | 484 | KWS | DCMVDYSIM | KFG | 1058.25 | 12.095 | 33.94% |
| MHC-II I_Ab | 3 | 384 | MKG | FYYYCSSSL | FYS | 1114.26 | 11.744 | 32.96% |
| MHC-II I_Ab | 4 | 324 | SVV | YIQTFVSLM | CLC | 1083.31 | 11.388 | 31.96% |
| MHC-II I_Ab | 5 | 192 | PKA | MSAPTPVSA | LVH | 841.98 | 11.24 | 31.54% |
| MHC-II I_Ad | 1 | 89 | FVF | SMVGVVFSN | NCI | 921.07 | 16.144 | 30.38% |
| MHC-II I_Ad | 2 | 282 | SFI | HMLSHAVFK | SCL | 1051.27 | 11.598 | 21.82% |
| MHC-II I_Ad | 3 | 77 | YNF | NYFCVVLSI | FVF | 1039.26 | 7.574 | 14.25% |
| MHC-II I_Ad | 4 | 387 | FYY | YCSSSLFYS | GGS | 1038.16 | 7.329 | 13.79% |
| MHC-II I_Ag7 | 1 | 332 | VSL | MCLCGLFFL | GGS | 1028.36 | 10.512 | 25.72% |
| MHC-II I_Ag7 | 2 | 33 | LDF | SSLELLQFQ | FRL | 1046.2 | 9.939 | 24.32% |
| MHC-II I_Ag7 | 3 | 373 | ILL | TYLYCYRLM | KGF | 1207.48 | 9.4 | 23.00% |
| MHC-II I_Ag7 | 4 | 457 | KFG | SFDVKYKFY | GDL | 1178.36 | 9.342 | 22.86% |
| MHC-II I_Ag7 | 5 | 71 | FYM | MEDYNFNYF | CVV | 1224.32 | 8.271 | 20.24% |
Table 2: Prediction of MHCII ligands all rows highlighted in red represent predicted binders to the MHC-II Allele i.e. MHC-II I_Ab, MHC-II I_Ad,MHC-II I_Ag7.(All rows highlighted in red represent predicted binders).
| Peptide Rank | Start Position | Sequence | Score | Predicted Affinity |
|---|---|---|---|---|
| 1 | 489 | YSIMKFGDF | 8.651 | High |
| 2 | 483 | SDCMVDYSI | 8.648 | High |
| 3 | 161 | ELVVSVALF | 8.643 | High |
| 4 | 96 | SNNCISMLI | 8.64 | High |
| 5 | 337 | LFFLGGSVS | 8.639 | High |
| 6 | 305 | GGQQDSRGY | 8.636 | High |
| 7 | 262 | SQIGFCFFG | 8.626 | High |
| 8 | 502 | SKIFVMGFS | 8.62 | High |
| 9 | 417 | WWLNYNSFV | 8.616 | High |
| 10 | 217 | CFSEVMFLD | 8.613 | High |
Table 3: Cascade SVM based high affinity tap binders of Dracunculus medinensis.
In this study, we found the antigenic determinants by finding the area of greatest local hydrophilicity. Hopp and Woods hydrophobicity scale is used to identify of potentially antigenic sites in proteins by analyzing amino acid sequences in order to find the point of greatest hydrophilic. Hydrophilicity Prediction result data found high in sequence position at 300-320 in a protein this scale is basically a hydrophilic index where apolar residues have been assigned negative values. The Window size of 5-7 is good for finding hydrophilic regions, greater than 0 values are consider as hydrophilic which is consider as antigenic. Welling used information on the relative occurrence of amino acids in antigenic regions to make a scale which is useful for prediction of antigenic regions and the predicted result data found high in sequence position 250-270. Welling antigenicity plot gives value as the log of the quotient between percentage in a sample of known antigenic regions and percentage in average proteins. We also study Hydrophobicity plot of HPLC / Parker Hydrophilicity Prediction Result Data found between 300-320 (Maximum Score-6.3) i.e., the maximum predicted residues at the position 308(Residue is Q) is 305- GGQQDSR -311 and 309(Residue D) is 306- GQQDSRG-312. BepiPred predicts the location of linear B-cell epitopes Result found that between 300-310 is 301-MQMGYIIHLC-310 and the maximum score (1.369) is found at the position 308,309. There are 15 antigenic determinant sequences is found by Kolaskar and Tongaonkar antigenicity scales (Figures 6a and Table 4) the results show highest pick at position.
| Start Position | Sequence | End Position | ||
|---|---|---|---|---|
| 1 | 4 | YIYWFGVILLCFLLLFVFFYDDFVFSLDFS SLELLQFQFRLDWFSFSFFCLLVMVVGS VVVYS | 66 | |
| 2 | 76 | FNYFCVVLSIFVFSMVGVVFSN | 97 | |
| 3 | 99 | CISMLIFWDLLGVSSYFLVLY | 119 | |
| 4 | 138 | VGDVCVFLVFCGLFF | 152 | |
| 5 | 156 | DFISLELVVSVALFFFILST | 175 | |
| 6 | 178 | KSAQYPFSSWLPKA | 191 | |
| 7 | 193 | SAPTPVSALVHSSTLVTAGLFLGMCFSE VM FLDFVLDFM FFVGLFT | 238 | |
| 8 | 251 | DIKKLVALSTLSQIGFCFFGLGLGLVYFSF IHMLSHAVFKSCLFM | 295 | |
| 9 | 297 | MGYIIHLCGG | 306 | |
| 10 | 310 | SRGYVGVGGLSSWYIQTFVSLMCLCGL FFLGG | 342 | |
| 11 | 344 | VSKEILLEHYFFCNWSLFLVFLFFFSILLT YLYCYRLMKGFYYYCSSSLFYSGGSLV FSFVSLVLVVFSIVFLW | 417 | |
| 12 | 421 | YNSFVISSSLLYSDYYSLFYLFILGLVLCV VFFK | 454 | |
| 13 | 456 | GSFDVKYKFYGDLLPKVIIRGNYVVKWS DCMVDYSI | 491 | |
| 14 | 496 | DFSFYVSKIFVM | 507 | |
| 15 | 514 | INFLFLCLFL | 523 | |
Table 4: The 15antigenic determinants of NADH dehydrogenasesubunit5.
4-YIYWFGVILLCFLLLFVFFYDDFVFSLDFSSLELLQFQFRLDWFSFSFFCLLVMVVGSVVVYS- 66, 76- FNYFCVVLSIFVFSMVGVVFSN- 97, 99-CISMLIFWDLLGVSSYFLVLY-119, 138-VGDVCVFLVFCGLFF- 152,156-DFISLELVVSVALFFFILST-175,178- KSAQYPFSSWLPKA-191,193-SAPTPVSALVHSSTLVTAGLFLGMCFSEVMFLDFVLDFMFFVGLFT- 238,251-DIKKLVALSTLSQIGFCFFGLGLGLVYFSFIHMLSHAVFKSCLFM- 295,297-MGYIIHLCGG- 306,310-SRGYVGVGGLSSVVYIQTFVSLMCLCGLFFLGG-342,344- SKEILLEHYFFCNWSLFLVFLFFFSILLTYLYCYRLMKGFYYYCSSSLFYSGGSLVFSFVSLVLVVFSIVFLW- 417,421-YNSFVISSSLLYSDYYSLFYLFILGLVLCVVFFK- 454,456-GSFDVKYKFYGDLLPKVIIRGNYVVKWSDCMVDYSI- 491, 496-DFSFYVSKIFVM-507, 514-INFLFLCLFL- 523.
Result of determined antigenic sites on proteins has revealed that the hydrophobic residues if they occur on the surface of a protein are more likely to be a part of antigenic sites. This method can predict antigenic determinants with about 75% accuracy and also gives the information of surface accessibility and flexibility. Further this region form beta sheet which show high antigenic response than helical region of this peptide and shows highly antigenicity. The Structure of the Dracunculus medinensis antigen-NADHdehydrogenasesubunit5 is predicted by SWISS-MODEL (automated protein structure homologymodelling server) DeepView (Swiss Pdb-Viewer) program (Figure 20). The purpose of this server is to make Protein Modelling accessible to all biochemists and molecular biologists worldwide. The target structure will also serve as a detailed model for determining the structure of peptide within that protein structure. We predict Solvent accessibility by using Emini et al. the result found the highest probability i.e. found 306-GQQDSR-311(maximum score 7.194), 307-QQDSRG- 312(maximum score 7.194), 308- QDSRGY-313(score-6.508),432- YSDYYS-437(6.379),178- KSAQYP-183(score 6.281), that a given protein region lies on the surface of a protein and are used to identify antigenic determinants on the surface of proteins. This algorithm also used to identify the antigenic determinants on the surface of proteins and Karplus and Schulz predict backbone or chain flexibility on the basis of the known temperature B factors of the a-carbons here we found the result with High score 1.122 maximum in 305- GGQQDSR-311. We predict Solvent accessibility of Dracunculus medinensis antigen NADH dehydrogenase subunit 5 for delineating hydrophobic and hydrophilic characteristics of amino acids. Solvent accessibility used to identify active site of functionally important residues in membrane proteins. Solvent-accessible surface areas and backbone angles are continuously varying because proteins can move freely in a three-dimensional space. The mobility of protein segments which are located on the surface of a protein due to an entropic energy potential and which seem to correlate well with known antigenic determinants. We also found the i.e. Sweet et al. hydrophobicity prediction result data found high in position 350-370, Kyte and Doolittle result high in position 10-20, Abraham and Leo result high in position 350-370, Bull and Breese result high in position 300-320, Guy result high in position 300-320, Miyazawa result high in position 350-370, Roseman result high in position 350-380,Wolfenden result high in position 440-460,Wilson et 350-380, Cowan 350-380,Chothia 400-430. These scales are a hydrophilic with a polar residues assigned negative value. Because the N- and C- terminal regions of proteins are usually solvent accessible and unstructured, antibodies against those regions recognize the antigenic protein. In this study, we found predicted MHC-I peptide binders of toxin protein for 8mer_H2_Db alleles with the consensus sequence QNWNCCTI that yields the maximum score i.e. 52.494, 9mer_H2_Db with, the consensus sequence FCIHNCDYM that yields the maximum score i.e. 50.365, 10mer_H2_Db with, the consensus sequence SGYYNFFWCL that yields the maximum score i.e. 58.858, 11mer_H2_Db with, the consensus sequence CGVYNFYYCCY that yields the maximum score i.e. 79.495 and I_Abwith the consensus sequence YYAPWCNNA that yields the maximum score i.e. 35.632,I_Ad with the consensus sequence QMVHAAHAE that yields the maximum score i.e. 53.145, MHC-II I_Ag7 with the consensus sequence WYAHAFKYV that yields the maximum score i.e. 40.873 for MHC II allele was tasted. We also use a cascade SVM based TAPPred method which found 160 High affinity TAP Transporter peptide regions which represents predicted TAP binders residues which occur at N and C termini from Dracunculus medinensis antigen NADHdehydrogenasesubunit5. TAP is an important transporter that transports antigenic peptides from cytosol to ER. TAP binds and translocate selective antigenic peptides for binding to specific MHC molecules. The efficiency of TAP-mediated translocation of antigenic peptides is directly proportional to its TAP binding affinity. Thus, by understanding the nature of peptides, that bind to TAP with high affinity, is important steps in endogenous antigen processing. The correlation coefficient of 0.88 was obtained by using jackknife validation test. In this test, we found the MHCI and MHCII binding regions. T cell immune responses are derived by antigenic epitopes hence their identification is important for design synthetic peptide vaccine. T cell epitopes are recognized by MHCI molecules producing a strong defensive immune response against antigen NADHdehydrogenasesubunit5. Therefore, the prediction of peptide binding to MHCI molecules by appropriate processing of antigen peptides occurs by their binding to the relevant MHC molecules. Because, the C-terminus of MHCI-restricted epitopes results from cleavage by the proteasome and thus, proteasome specifity is important for determing T-cell epitopes. Consequently, RANKPEP also focus on the prediction of conserved epitopes. C-terminus of MHCI-restricted peptides is generated by the proteasome, and thus RANKPEP also determines whether the C-terminus of the predicted MHCI-peptide binders is the result of proteasomal cleavage. Moreover, these sequences are highlighted in purple in the output results. Proteasomal cleavage predictions are carried out using three optional models obtained applying statistical language models to a set of known epitopes restricted by human MHCI molecules as indicated here.
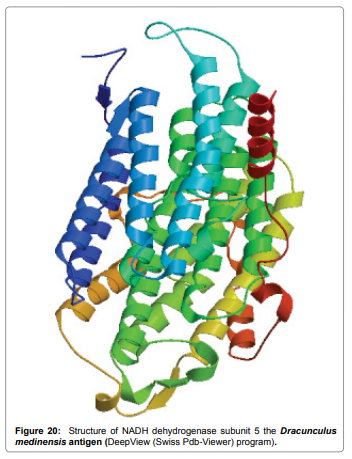
Figure 20: Structure of NADH dehydrogenase subunit 5 the Dracunculus medinensis antigen (DeepView (Swiss Pdb-Viewer) program).
From the above result and discussion it is concluded that the ability of RANKPEP to predict MHC binding peptides, and thereby potential T-cell epitopes, antigenic peptide that binds to MHC molecule are antigenic that means hydrophilic in nature. This means the increase in affinity of MHC binding peptides may result in enhancement of immunogenicity of Dracunculus medinensis antigen NADH dehydrogenase subunit5 and are helpful in the designing of synthetic peptide vaccine. This approach can help reduce the time and cost of experimentation for determining functional properties of Dracunculus medinensis antigen NADHdehydrogenasesubunit5. Overall, the results are encouraging, both the ‘sites of action’ and ‘physiological functions’ can be predicted with very high accuracies helping minimize the number of validation experiments.