Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2014) Volume 2, Issue 3
Agricultural crops are an essential base component of all ecosystems and it is important to assess the impact of nanoparticles on them and the risks involved. In this study we have investigated the impact of the different concentrations (ppm) of citrate stabilized gold nanoparticles (GNPs) ranging from 100 to 400 on physiological and biochemical parameters of the seedlings of Brassica juncea as a model oil crop system. GNPs were synthesized and characterized through Uv-visible spectra and transmission electron microscopy. The qualitative accumulation of GNPs into seedlings of Brassica juncea was ascertained through cyclic voltammetry and Laser Induced Breakdown Spectra (LIBS). Exposure of GNPs caused a decrease in overall growth of Brassica juncea was attributed to increase in free radical stress, supported by a general increase in biomarkers such as antioxidative enzymes, proline and hydrogen peroxide due to the formation of reactive oxygen species.
Keywords: Gold nanoparticles; Brassica juncea; Physiological parameter; Antioxidative enzymes, Antioxidant potential
The investigation of risk effects of engineered nanoparticles over ecosystem has recently attracted much attention and it is often concluded that nanoparticles can exert cytotoxicity to animals and plant systems [1-3]. In this context, there are a few reports on the abiotic stress imposed by the impact of engineered nanoparticles leading to toxicity in plant cells [4-6]. The effect of alumina nanoparticles with or without phenanthrene coating reveals that uncoated alumina nanoparticles inhibits root elongation of corn, cucumber, soybean, cabbage and carrot [7]. Many other nanoparticles viz. multi-walled carbon nanotubes, aluminum, alumina, zinc, and zinc oxide nanoparticles also has demonstrated inhibitory effect in root growth of corn and has practically terminated root development in the radish, rape, ryegrass, lettuce, corn, and cucumber [4]. The culture medium containing copper nanoparticles has reported to induce negative effect in root elongation of Phaseolus radiates [8]. The seedling of Chilopsis linearis supplemented with mercuric acetate and chloroauric acid in hydroponics have shown reduction in root elongation by 25% and 55% than roots of control seedlings [9].
The biotic and or abiotic stress condition induced by metal and metal oxide nanoparticles over crop systems accelerates the formation of Reactive Oxygen Species (ROS), resulting in oxidative damage at cellular level. Thus, antioxidant enzymes such as Catalase (CAT), Ascorbate Peroxidase (APX), Guaiacol Peroxidase (GPX) and Glutathione Reductase (GR) functions to interrupt the cascades of uncontrolled oxidation in some organelles [6,10,11]. For example, increase in the production of ROS were found in Medicago sativa cells in presence of CdSe/ZnS quantum dots in suspension culture [6]. These antioxidative enzymes play role in eliminating H2O2 and are distributed in at least four distinct cell compartments [12-14]. Furthermore, imposition of stress conditions could give rise to excess concentrations of ROS, resulting in oxidative damage at cellular level. This leads to an increase in potential of antioxidant enzymes, which functions to interrupt the uncontrolled oxidation damage [15].
Recently, GNPs due to their inherent surface area to volume ratio are used as filler for polymer matrix precursor for development of drug delivery carrier for biomedical applications and as industrial Catalyst [16-20]. This tremendous industrial potential of GNPs has led to the hazard of disposal the related materials to environment, imposing biomagnifications and risk over ecosystem [11,20,21]. Recently, we have observed that GNPs up to 10 ppm concentration can improve the growth of Brassica juncea (B. juncea; leading to an increase in the net productivity in terms of seed yield [14]. The present study involves preparation, characterization and investigation of impact of GNPs on physiological and biochemical parameters of seedlings of B. juncea. The study reveals a general decrease in growth of the plant system when administered of over 10 ppm concentration of GNPs, due to increase in free radical stress. This has been supported by a general increase in biomarkers such as antioxidative enzymes, proline and hydrogen peroxide due to the formation of ROS [6,14,15].
Starting materials
All chemical agents including chloroauric acid (HAuCl4 x 4H2O) were obtained from Himedia Chemicals India. Trisodium citrate stabilized GNPs with size 50 nm were synthesized as per the procedure reported [14]. In brief, a solution of chloroauric acid (500 mL, 1.0 X10-3 M) was boiled and stirred vigorously in 1 L round- bottom flask. To this, sodium citrate (50 mL, 38.8 mM) was added. The solution was allowed to vortex until a change in color was observed. Further boiling was continued for another few minutes, thereafter heating was discontinued, but stirring continued for an additional 15- 20 min. The solution was allowed cool to room temperature to obtain the citrate stabilized GNPs.
Treatment of GNPs to plant seed
Seeds of B. juncea were rinsed with distilled water, followed by successive washings with Tween-20 over 20 minutes and cleaned under running deionized water, to remove all traces of detergent. Surface sterilization of seeds was done with 70% alcohol for 1 minute followed by treatment with 0.1% Mercuric chloride (HgCl2) for five minutes. The seeds were then thoroughly washed with autoclaved distilled water. Washed seeds were soaked in sterile deionized water for four hours. Twelve seeds per bottle were placed in minimal salt (MS) media supplemented with different concentration 0, 100, 200, 300.00, and 400 ppm of GNPs. MS medium was prepared by mixing the following macronutrients (in g/L at pH 6.2 ± 1): NH4NO3,33.0; KNO3,38.0; MgSO4.7H2O,7.4; KH2PO4,3.40; H3BO3,1.24; MnSO4.4H2O,4.46; CaCl2.2H2O,8.8 with micronutrients(in g/L at pH 6.2 ± 1):ZnSO4.7H2O,1.72; KI, 0.166l; Na2MoO4.2H2O,0.05; CuSO4.5H2O,0.005; CoCl2.6H2O,0.005; iron source: FeSO4.7H2O,5.56; Na2 EDTA and organic nutrients 7.46; nicotinic acid, 0.1; pyridoxine hydrochloride, 0.1; thymine hydrochloride, 0.1; glycine, 0.4; myoionisitol. The contents were mixed with agar-agar 0.70 g/L to prepare the MS basal medium. After inoculation, the bottles were kept in the incubation chamber at 26±10 ºC and humidity (70%). Illumination was provided with three florescent tubes (40 W, flux density 16 W m- 2) with a 16/8hrs day/night cycle. All the physiological and biochemical experiments were conducted under controlled aseptic conditions and the respective data was recorded after 10 days of treatment.
Spectral characterizations
Uv-Visible spectra was recorded over Genesis-10 Thermospectronic spectrophotometer. Plant seedlings collected were dried at 60°C and LIBS of their dried powdered samples were recorded over LIBS 2000þ spectrometer (Ocean Optics, USA) equipped with CCD by using a Q-switched Nd: YAG laser at 1064 mm, with 28 ns pulse duration.
In the present method, a plasma plume was produced above the surface by ablation of a small quantity of plant sample using intense laser pulse. The ablated material was vaporized then partly atomized, excited and ionized. The excited atoms and ions in the plasma emitted a secondary light, which was then analyzed by a light detector. The emission spectrum from the plasma plume was detected and used for identification of the elements present in the ablated plant material [22].
Cyclic voltammetry
For cyclic voltammetry based quantification of GNPs the plant samples were homogenized with PBS (200 mM, pH 7.0) and centrifuged (10000 rpm). The extract was then diluted with PBS to 0.1ppm and scanned over cyclic voltammeter. Cyclic voltammograms (CV) were recorded over IVIUM Potentiostat Galvanostat at scan rate of 0.05 V/s at -1.1V using a three-electrode cell equipped with Ag/AgCl (3M NaCl) as reference, platinum as counter and glassy carbon as working electrodes.
Transmission Electron Microscopy
The sample preparation for TEM analysis of root samples of seedlings treated with GNPs were carried out by the treatment of the root sample with Karnovsky solution (4% formaldehyde + 5% glutaraldehyde at room temperature followed by incubation in a buffer (0.025 M cacodilate buffer, pH 6.9) solution at 4°C. The treated plant samples were embedded into Epon, then approx. 1-mm-thick, hand-cut cross-sections were made and dehydrated through an ethanol series (30 to 100%) and were then finally treated with propylene oxide and infiltrated with Epon. Sample was left for polymerization overnight at 60°C. Ultra-thin sections (70 to 90 nm) were cut from the blocks and observed under transmission electron microscope over JEOL 1011 (Tokyo, Japan) with primary beam voltage of 80 kV [23].
Growth analysis and biochemical assay
Ten days old seedlings were taken out from the tissue culture bottles then roots and shoots were separated. The physiological parameters of the seedlings were analyzed in terms of root and shoot length with the help of standard scale. Twelve seedlings per treatment were used for determination of physiological parameters. Antioxidant potential was investigated through monitoring the activity (μM/min/ mg protein) of four different antioxidative enzymes viz. Ascorbate Peroxidase (APX), catalase (CAT), Guaiacol Peroxidase (GPX) and Glutathione Reductase (GR) with concentration of GNPs. For this purpose, leaf of seedlings (2.0 g) was homogenized in extraction buffer (10 mL). The homogenate was filtered and centrifuged (13000) for 30 min at 4ºC. All Enzyme activities were expressed as units per mg of protein. Protein concentration of the enzyme extract was estimated [24]. For APX (EC 1.11.1.11) assay, leaves were homogenized in sodium phosphate buffer (100 mM, pH 7.0) containing ascorbate (5 mM), glycerol (10%), and ethylene diammine tetra acetic acid (EDTA, 1 mM). The activity was determined in a reaction mixture (1 mL) containing phosphate buffer solution (PBS, 50 mM, pH 7.0), ascorbate (0.1 mM), H2O2 (0.3 mM), and enzyme extract (50 μL). The fall in absorbance of the reaction mixture was recorded at 290 nm with extinction coefficient of 2.8 mM-1 cm-1 [25]. For CAT (EC 1.11.1.6) activity, leaves were homogenized in sodium phosphate buffer (100 mM, pH 7.0) containing EDTA (1 mM). The assay mixture consisted of enzyme extract (50 μL), PBS (100 mM, pH 7.0), EDTA (0.1 μM), and H2O2 (20 mM) in a total volume 1.5 mL. The decrease in absorbance of the reaction mixture was recorded at 240 nm using extinction coefficient of 40 mM-1 cm-1 [26]. For GPX (EC 1.11.1.7) activity, 3.0 mL of mixture contained PBS (50 mM, pH 7.8), guaiacol (5 mM), H2O2 (12.3) mM and enzyme extract (100 μL). The GPX activity was determined using the extinction coefficient as 26.6 mM cm-1 at 436nm [27]. GR (EC 1.8. 1.7) activity was determined by oxidation of NADPH at 340 nm with the extinction coefficient of 6.2 mM cm-1 [28]. The reaction mixture consisted of PBS (100 mM, pH 7.8), EDTA (2 mM), NADPH (0.2 mM) and glutathione 0.5 mM (oxidized form, GSSG) with enzyme extract in a 1 mL volume. For the determination of H2O2 levels, plant material (0.8g) was homogenized with trichloroacetic acid (5 mL, 0.1% w/v) followed by centrifuge (10000 g) of homogenate for 30 min at 4°C. Supernatant (0.5 mL) was added to PBS (0.5 mL, 10 mM, pH 7.0) and KI (1 mL ,1 M). The absorbance of the supernatant was measured at 390 nm. The H2O2 content in micro mole per gram fresh weight (μM/g. F.wt.) of the sample was determined using standard curve [29]. In order to estimate free proline content, seedlings (1.0g) were homogenized in sulfosalicylic acid (SSA (3% w/v, 10 mL) followed by centrifugation of homogenate (10000 g) for 20 min. The supernatant (2.0 mL) was mixed with glacial acetic acid (2.0 mL), ninhydrin reagent (2.0 mL) and the contents were incubated for 1hr at 100 ± 1°C. To each tube of reaction mixture, toluene (4.0 mL) was added to extract the red colored chromophore under vortex. The absorbance of chromophore containing toluene layer was measured at 520 nm. In blank, SSA (2.0 mL, 3% v/v) was used in place of the supernatant (Figure 2b). Concentration of proline in micro gram per gram fresh weight (μg./g. F.wt) of the sample was evaluated from a standard curve of L-proline. For determination of MDA content (μg./g. F.wt), plant material (0.1 g) was homogenized in TBA (1 ml, 0.25% w/v prepared in 10% TCA] at room temperature and then heated at 95 ± 1°C for 30 min. The homogenate was centrifuged at 8,000 rpm for 30 min and the absorbance of supernatant was taken at 532 nm and 600 nm using TBA (0.25% in 10% TCA) as blank. The concentration of malonaldehyde content (MDA) in (μg./g. F.wt) was calculated by using an extinction coefficient of 155 mM-1 cm-1 All the data plotted are an average of three independent experiments with five replicates each and presented as means ± SD (n=5) [30].
Effect of GNPs on the average shoot length (ASL) and Average Root Length (ARL) along with physiological parameters, antioxidant potential, proline and hydrogen peroxide contents in the seedlings of B. juncea was investigated in MS media under laboratory conditions. For this purpose, citrate stabilized GNPs was synthesized and characterized. Uv-visible spectra revealed absorption corresponding to chloroauric acid at 218 nm. Reduction of chloroauric acid has afforded GNPs with absorption at 224 nm and coupled with characteristic plasmonic absorption at 536 nm of GNPs (Figure 1a). The absorption of GNPs corresponding to 536 nm refers to the particle size around 50 nm [31]. TEM revealed the formation of GNPs in the form of mixtures bearing spheres with particle size ranging 15 to 45 nm (Figure 2a) and nanorods with length ranging 145 to 24 nm and diameter 30 nm (Figure 2a).
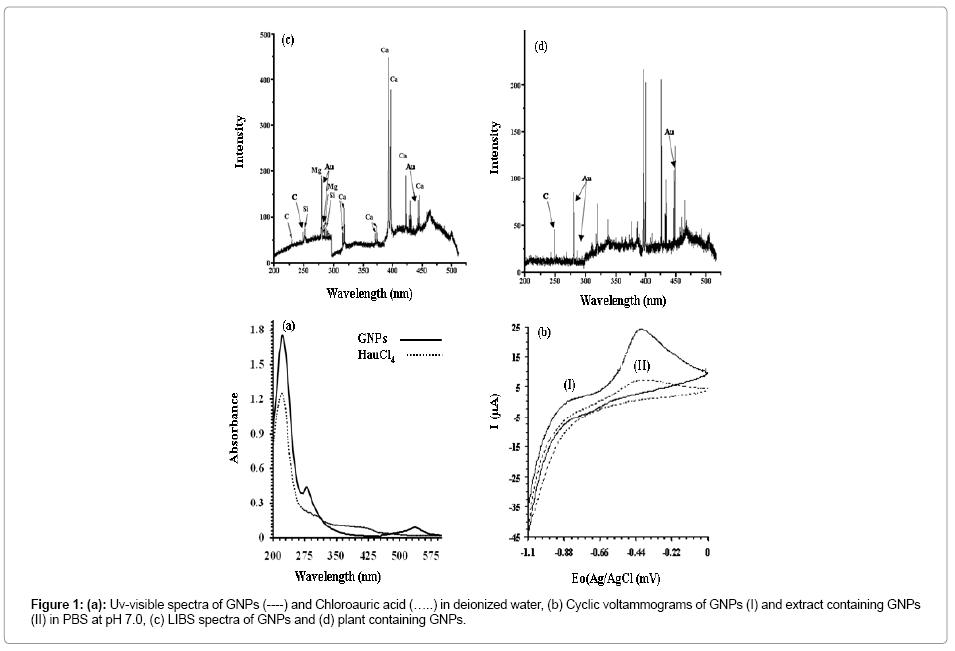
Figure 1: (a): Uv-visible spectra of GNPs (----) and Chloroauric acid (…..) in deionized water, (b) Cyclic voltammograms of GNPs (I) and extract containing GNPs (II) in PBS at pH 7.0, (c) LIBS spectra of GNPs and (d) plant containing GNPs.
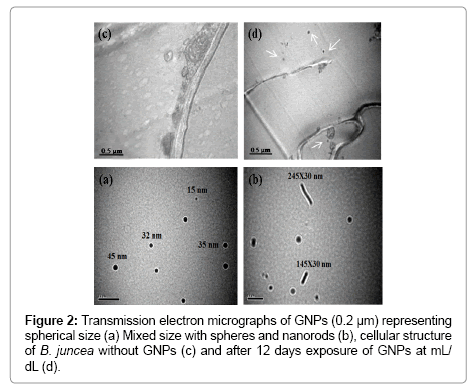
Figure 2: Transmission electron micrographs of GNPs (0.2 μm) representing spherical size (a) Mixed size with spheres and nanorods (b), cellular structure of B. juncea without GNPs (c) and after 12 days exposure of GNPs at mL/ dL (d).
Accumulation of GNPs in Seedlings
The accumulation of GNPs in seedlings of B. juncea has clearly been reflected qualitatively through cyclic voltammetry (Figure 1b), LIBS (Figure 1c and d) and TEM (Figure 2c and d). Extract of the seedlings supplemented with GNPs exhibit oxidation peak currents of 12.12 μA at -0.46 V, whereas, the voltammograms of the extract of seedlings without GNPs supplementation in PBS (200 mM, pH 7.0) does not indicate any electrochemical behavior (Figure 1b). LIBS of GNPs indicates characteristic atomic lines corresponding to Au at 278.32, 282.43, and 438.33 nm [22] (Figure 1c). The sample prepared from the leaves of the plant supplemented with GNPs showed atomic lines corresponding to Au at 279.69, 280.49, and 431.59 nm (Figure 1d). The voltammograms GNPs showed oxidation peak current of 19.8 μA at -0.46 (Figure 1b). TEM images of longitudinal section of root of seedlings grown in MS media showed normal anatomy of various cell organelles of the root cell (Figure 2c). The root cell of seedlings grown in the MS media supplemented with GNPs shows the location of nanoparticles within the root at different applied concentration of GNPs (Figure 2d). The spectral, microscopic and electrochemical observations indicate the accumulation of GNPs in the seedlings of B. juncea.
Impact assessment of GNPs on B. juncea
Physiological data revealed a marginal increase in the ASL with overall decrease in the ARL (Figure 3a). With concentration of GNPs, a marginal increase in the ASL ranging 6.81to 7.36 cm was observed in the seedlings of B. juncea (Figure 3a). A remarkable growth in ASL of B. juncea in MS media devoid of GNPs was observed over the seedlings grown in the MS media supplemented with GNPs. With concentration of GNPs, a marginal increase in the ASL was recorded. This has contributed a simultaneous increase in the ARL of B. juncea ranging 9.42 to 11.62 cm up to the supplement of 100 ppm of GNPs. Further increase in the concentration of GNPs has shown the decline in the ARL to 9.43 cm. Such decline in the ARL of the seedlings might be caused by the diversion of metabolic energy and metabolites from the processes involved in the normal growth and development to the processes involved in alleviating the toxic effects imposed by GNPs [32,33].
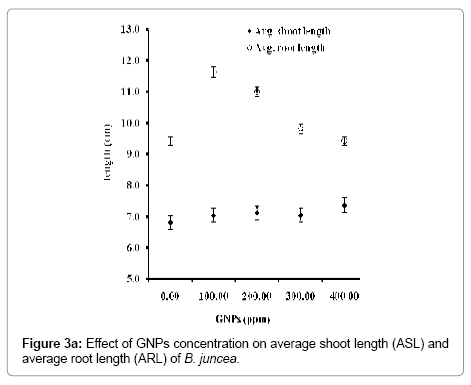
Figure 3a: Effect of GNPs concentration on average shoot length (ASL) and average root length (ARL) of B. juncea.
With concentration of GNPs, the antioxidant potential in terms of the activities of antioxidative enzymes increased regularly in the seedlings (Figure 3b). Such increase in antioxidative enzyme activity may be due to the stress over the seedlings of B. juncea imposed by GNPs. This resulted in improved protection against ROS by detoxifying H2O2, preventing the harmful effects caused by further reactions of H2O2 (Figure 3c). Cell antioxidant system, i.e., antioxidant enzymes and metabolites, closely controls the ROS production and their toxic effects under oxidative stress conditions [34]. APX plays a critical role in detoxifying H2O2 under stress conditions induced by GNPs. A marginal variation in the APX activity was recorded up to the concentration of GNPs 200 ppm. Further increase in the concentration of GNPs contributed a remarkable increase in the APX activity with maximum at 400 ppm concentration of GNPs. This corresponds to 1.53 times increase in APX activity more over the control. Furthermore, this has contributed a simultaneous increase in the GPX activity by 1.28 times greater than the control. Studies further indicated that at 200 ppm concentration of GNPs, the CAT activity of the seedlings was found to be increased by 1.37 times than control. Further increase in concentration of GNPs, did not contributed to any significant variation in the CAT activity of the seedlings. Such variations in the CAT activity depending on the concentration of GNPs may be due to the scavenging and converting H2O2 into O2 and H2O up to the 400 ppm concentration of GNPs [6]. The GR activities among seedlings grown in MS media supplemented with different concentration of GNPs were determined. It was found that at highest level of supplementation of GNPs to the MS media i.e. at 400 ppm the GR activity increased by 1.41 folds with respect to the control. Higher APX, GR and GPX activity than CAT activity at higher concentration of GNPs suggests that APX, GPX and GR provide better defense mechanism against GNPs induced oxidative damage in B. juncea.
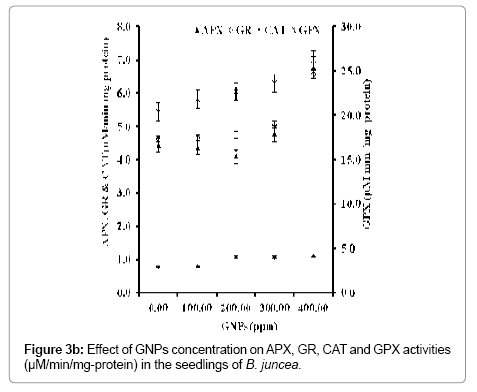
Figure 3b: Effect of GNPs concentration on APX, GR, CAT and GPX activities (μM/min/mg-protein) in the seedlings of B. juncea.
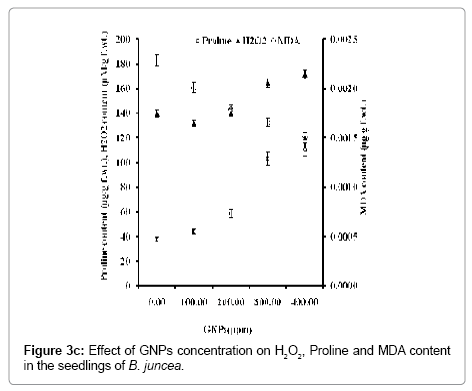
Figure 3c: Effect of GNPs concentration on H2O2, Proline and MDA content in the seedlings of B. juncea.
The evidence of the generation of ROS has clearly been reflected due to the increase in proline and H2O2 contents with concentration of GNPs (Figure 3c). At highest supplemented concentration of GNPs (400 ppm), seedlings accumulate 2.9 times higher level of proline content in B. juncea than the control, supporting that GNPs induce oxidative stress in the seedlings. This shows higher scavenging of free radical and protects the native protein structure and maintains NADH/ NAD+ ratio. Thus in the present investigation, proline accumulation helps the seedlings to alleviate the GNPs induced stress and to maintain the normal cellular metabolism. With proline content, the H2O2 content can also be safely taken as a reliable biochemical marker to evaluate the tolerance to GNPs induced stress in B. juncea. At the highest GNPs concentration H2O2 content in seedlings was increased to 1.2 times with respect to control seedlings. With concentration of GNPs, increase in H2O2 content indicates the induced stress imposed by the GNPs in the seedlings of B. juncea [13,14,26]. With concentration of GNPs, the potential of four antioxidative enzymes viz., APX, CAT, GPX and GR in terms of their activity of was increased (Figure 3b). This has rendered a simultaneous increase in the proline and H2O2 contents (Figure 3c) that indicates the production of ROS leading to the stress imposed by GNPs over seedlings of B. juncea. The results indicate that free radical-mediated oxidative damage plays an important role in the GNPs induced cellular toxicity in the seedlings of B. juncea with better protection against free radicals by maintaining specific activity ratio of antioxidant enzymes.
Here, we have for the first time reported the effect of different concentrations of citrate stabilized gold nanoparticles (GNPs) on variations in physiological and biochemical parameters of seedlings of B. juncea, grown in minimal salt media under laboratory conditions. Characterization of GNPs and their accumulation into seedlings has been ascertained through spectra, microscopy and cyclic voltametry. The supplement of GNPs imposed a marginal increase in the average shoot length ranging 6.81 to 7.36 cm in the seedlings of B. juncea. This has contributed a greater increase in the average root length up to 11.62 cm of the seedlings at 100 ppm concentration of GNPs. With concentration of GNPs, a regular increase in the antioxidative enzyme activities, H2O2 and proline contents were recorded. Results indicate that production of reactive oxygen species depends on the concentration of GNPs, that impose physiological and biochemical stress over the seedlings of B. juncea.