Journal of Pollution Effects & Control
Open Access
ISSN: 2375-4397
ISSN: 2375-4397
Research Article - (2015) Volume 3, Issue 1
The purpose of the present study was to assess the effects of consumption of catfish (Clarias gariepinus) exposed to palm kernel oil (PKO) biodiesel on functions and oxidative status of liver of albino rat. The author measured the responses of growth performance (average daily weight gain, feed conversion ratio, body weight, and relative liver weight), oxidative stress, and indices of liver functions of rat placed on feed formulated with catfish exposed to borehole water mixed with different concentrations of PKO biodiesel ( 0 % w/v, 0.1 % w/v, 0.25 % w/v). The results showed that fish exposed to PKO biodiesel had no effect on relative liver weight, but average body weight was significantly reduced, serum levels of total and direct bilirubin were significantly elevated while those of bilirubin and globulin were significantly reduced. However, specific activities of catalase (CAT), superoxide dismutase (SOD) and glutathione-s-transferase (GST) of serum of rats fed with catfish exposed to lower concentration of PKO biodiesel (BD 0.1) were significantly higher than those of rats fed with catfish exposed to higher concentration of PKO biodiesel (BD 0.25) (p<0.05) while both are significantly lower than that of the control rats (p<0.05). Serum level of MDA in test rats increased significantly (p<0.05) as the concentration of PKO biodiesel increased. The data clearly demonstrate that consumption of catfish exposed to PKO biodiesel can result in liver dysfunction at a degree not dependent on concentration of PKO biodiesel by a mechanism of oxidative stress which is dependent on the concentration of PKO biodiesel.
Keywords: Palm kernel oil; Biodiesel; Oxidative status; Hepatic; Catfish
Biodiesel is defined by ASTM International as a fuel composed of monoalkyl esters of long-chain fatty acids derived from renewable vegetable oils or animal fats meeting the requirements of ASTM D6751 [1]. Vegetable oils and animal fats are principally composed of triacylglycerols (TAG) consisting of longchain fatty acids chemically bound to a glycerol (1,2,3-propanetriol) backbone. The chemical process by which biodiesel is prepared is known as the transesterification reaction, which involves a TAG reaction with a short-chain monohydric alcohol normally in the presence of a catalyst at elevated temperature to form fatty acid alkyl esters (FAAE) and glycerol. The conversion of TAG to biodiesel is a stepwise process whereby the alcohol initially reacts with TAG as the alkoxide anion to produce FAAE and diacylglycerols (DAG, reaction [2].
Free radicals are atoms or molecules with unpaired electrons. Free radicals in the organism are of endogenous and exogenous origin. Endogenously, they occur in mammals during mitochondrial electron transport chain, oxidative reactions in phagocytic and endothelial cells, redox cycles, arachidonic acid metabolism, and autooxidation reactions [3]. Industrial pollutants, drugs, diet, ionizing radiation, ultraviolet light, cigarette smoke, and xenobiotics are exogenous sources of free radicals [3]. Free radicals are classified as Reactive Oxygen Species (ROS) and reactive nitrogen species (RNS). Superoxide radical, hydrogen peroxide, hydroxyl radical, hypochlorous acid, singlet oxygen, alkyl radical, hydroperoxyl radical, and alkoxyl radical are effective as ROS whereas nitric oxide is the most well known RNS [4,5]. Reactive oxygen species cause protein denaturation, damage in nucleic acids, and lipid peroxidation. As a result, impairment occurs in the membrane structure and cellular functions. Reactive oxygen species are involved in pathogenesis of many diseases [6]. When reactive oxygen species start to increase, phospholipids, glycolipids, glycerides, and sterols present in the cell membrane are decomposed. Peroxides, alcohols of these, ethane and pentane, nitric oxide and malondialdehyde are formed. Ultimately, oxidative damage is shaped [7].
Liver is the primary organ for the detoxification, as well as being the target of many of environmental chemicals including biodiesel. Today, we are all exposed to mixtures of different chemicals at a time because of industrialization and fast life style. Hepatic toxicity of chemical and/ or biological mixtures may have serious effects on the functions of liver, which is the non-stopping machine for biotransformation as well as several very important physiological processes.
McCormick [8] reviewed biodiesel emission data published through 2001 for heavy-duty engines. It was clear there were substantial reductions in the emissions of particulate matter (PM), carbon monoxide (CO), and hydrocarbon (HC), although there was an increase in nitrogen oxides (NOx) obtained by using B20 (blends of 20% biodiesel with 80% conventional diesel). A more detailed analysis of blends of biodiesel emissions data has indicated that the solid carbon fraction of the PM was reduced and the soluble organic fraction (SOF) was increased [9]. Some studies have compared biodiesel and conventional diesel extracts using mutagenicity assays [10]. The acute toxicity of biodiesel was assayed by the Microtox test, which demonstrated an increased toxicity in the semivolatile fraction compared with the particulates [11], and was shown to be a potent inflammatory agent [12] compared with exposure of diesel PM extracts. Moreover, the type of motor, fuel, particle size, and chemical composition has all been shown to influence the mechanisms and magnitude of adverse manifestations [13].
Most biodiesel studies have focused on the analysis of exhaust emission material, motor efficiency, and the different processes for obtaining biodiesel with oleaginous microorganisms and palm, soybean, kitchen oil, palm oil, or animal fat [13,14]. Due to leaks in storage, fuels pollute underground waters. In that case, humans and animals are exposed to biodiesel cutaneously and orally. There are numerous studies investigating the effects of inhaled diesel and biodiesel fuels. However, there are very few studies on the fuels taken orally, directly [15,16] or indirectly through the food chain [17]. As a result of paucity of information, the present investigation was embarked upon to study the effects of catfish exposed to palm kernel oil (PKO) biodiesel on the functions and oxidative status of liver of rats.
Reagents and solvents were of analytical grade and are products of British Drug House, Poole, England.
Palm Kernel Oil (PKO)
Palm kernel oil was purchased at the local market in Effurun, Nigeria. 100 g PKO was used for the transesterification process. The ethanol used (99% pure) is an analytical grade with boiling point of 78°C; while the NaOH used was also an analytical grade product of Aldrich Chemicals, England. The blender used was a Dry and Wet mill Blender with a clear glass (1,250 cc capacity) containers and stainless steel cutting blades. Other major materials used include scales, translucent white plastic container with bung and screw-on cap, funnels, PET bottles and thermometer.
Preparation of bio-diesel from PKO
Biodiesel was prepared from PKO in accordance with the method described by Aremu et al. [18].
Experimental water and fish treatment: The Biodiesel from PKO was diluted with borehole water to obtain 0.25 and 0.1% v/v. Twentyfour healthy juvenile catfish (Clarias gariepinus) were obtained from a commercial fish pond at Ekpan in Delta State, Nigeria and acclimatized for ten days prior to the commencement of the experiment. The catfish were grouped into three (3) of eight catfish and were kept in 30 L plastic aquaria. Group A served as control and the catfish here were cultured in borehole water while those in Groups B and C were exposed to the different mixtures (0.1% v/v and 0.25% v/v respectively) of Biodiesel from PKO. The catfish were fed ad libitum with commercial fish meal for 30 hrs during which the experiment lasted. Each of these treatments had three replicates.
At the end of the 30 h experimental period, the catfish were harvested, oven dried at 40°C and used as a source of protein (25%) to formulate diet for albino rats. The diet for each group was formulated by mixing known quantities of sources of each food class comprising corn starch (52%), oil (4%), maize cob (4%), sucrose (10%) and vitamin/mineral mixture (5%). The food items were mixed together and manually made into pellets to feed albino rats. Proximate analysis of the experimental diet is shown in Table 1.
| NUTRIENT | Experimental diet | ||
| CONTROL | BD0.1 | BD0.25 | |
| PROTEIN (%) | 20.2±2.2a | 18.39±2.1a | 18.19±1.9a |
| FAT (%) | 7.7±1.2a | 4.88±1.2b | 3.96±1.1b |
| ASH (%) | 6.2±1.1a | 7.38±0.7a | 7.30±0.7a |
| FIBRE (%) | 5.3±0.5a | 9.22±1.1b | 5.56±0.9a |
| WATER (%) | 6.3±0.7a | 8.83±1.2b | 7.83±1.1ab |
| CARBOHYDRATE (%) | 54.3±2.4ab | 51.30±2.4a | 57.16±2.5b |
Table 1: Proximate composition of experimental diet.
Experimental rats and treatment: Thirty albino rats of (51 ± 2.0 g) were procured from the animal house of Department of Anatomy, University of Benin, Benin-City, Nigeria. All animals were housed under standard laboratory conditions with free access to water adlibitum and balanced pellets food. The housing temperature was (25 ± 1°C) with 12 h dark /light cycle and 50% humidity. All ethical guidelines on the use of animals for investigational purposes were followed and the experiment protocol was approved by Federal University of Petroleum Resources, Effurun (FUPRE), Nigeria ethics committee.
The animals were grouped into three with each group containing ten rats. The rats in Group 1 served as the control and they were fed on the control diet, which was formulated with catfish cultured in borehole water. Animals in Groups 2 and 3 were fed on diet formulated with catfish exposed to the different mixtures of biodiesel (0.1 and 0.25% v/v respectively). The feeding lasted for a period of thirty (30) days (after an acclimatization period of ten days) during which the weight and feed intake were monitored.Each of these treatments had three replicates.
At the end of the feeding exercise, the rats were anaesthetized by placing them in a jar containing cotton wool soaked with chloroform before being sacrificed by jugular puncture. The liver of experimental rat was removed into a beaker containing ice cold 0.25 M sucrose solution. The blood was obtained through their jugular veins. Each blood sample was thereafter centrifuged at 3500 rpm for about 15 min using refrigerated centrifuge RC650s and the serum obtained was preserved at -8°C until required for use. A portion of the liver was homogenized for biochemical studies and enzyme assays. MDA determination was based on method described by Bird et al. [19]. The method described by Jollow et al. [20] was used to determine reduced glutathione (GSH) concentration. Catalase activity was determined according to the method of Sinha [21]. The activity of superoxide dismutase (SOD) was determined by the method of Misra and Fridovich [22]. The cytosolic glutathione s-transferase activity was determined spectrophotometrically at 37°C (340 nm) by the procedure described by Habig et al. [23]. The activity of Gamma glutamyltranspeptidase (GGT) was determined following the method described by Tietz [24]. Concentrations ofserum bilirubin and albumin were determined by a method earlier reported [24]. The determination of serum globulin concentration was carried out using the method described by Tietz [25] by subtracting the concentration serum albumin from the total protein content determined using Biuret method as described by Henry et al. [26]. Activities of alanine transaminase (ALT) and aspartate transaminase (AST) in the liver of experimental animals were determined following the procedure reported by Reitman and Frankel [27] modified by Schmidt and Schmidt [28].
All numerical results were obtained from the three (3) groups (control and treated). Data obtained were presented as mean ± SEM and subjected to statistical analysis using a one way analysis of variance (ANOVA) by employing the method of Steel and Torrie [29]. Significant difference between the treatment means was determined at 95% confidence level using Duncan’s Multiple range test [30]. Posthoc comparison using Duncan’s New Multiple Range Test (DNMRT) was employed. DNMRT is a One-Way Analysis of Variance (ANOVA) that allows multiple comparisons. The error rate of each pairwise comparison is considered rather than an overall rate, and allows a higher rate for pairs of sample averages that are further apart when ordered by size.
Rats were placed on diets formulated with catfish exposed to biodiesel contaminated water as source of protein. The performance of rats placed on the diets formulated with catfish exposed to biodiesel (test diet) is presented in Table 2. Average daily weight gain of rats placed on the BD 0.25 diets is 1/3rd that of rats placed on the control diet. In contrast, there was no significant difference in the average daily feed intake between the control diet and the test diet (p>0.05). The feed conversion ratio (FCR) of test rats was significantly higher than that of the rats placed on control diet (p<0.05).
| Group of Rats | Average daily weight gain/loss (g/rat/day) | Average daily feed intake (g/rat/day) | Feed Conversion Ratio (FCR) |
| Control | 0.20±0.003a | 6.39±0.18a | 10.65±0.23a |
| BD0.1 | 0.012±0.002b | 6.33±0.27a | 15.85±0.19b |
| BD0.25 | 0.007±0.001c | 6.36±0.16a | 15.90±0.21b |
Table 2: Performance of rats placed on diet formulated with catfish exposed to biodiesel polluted water.
The growth of rats placed on diets formulated with catfish exposed to biodiesel was monitored over a period of 30 days and the growth response curve is presented in Figure 1. Generally, both test and control rats showed positive growth response. The growth of controls was found to be significantly higher than that of the rats placed on diets formulated with catfish exposed to biodiesel.
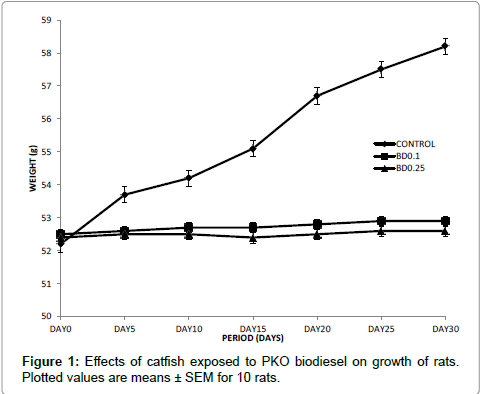
Figure 1: Effects of catfish exposed to PKO biodiesel on growth of rats. Plotted values are means ± SEM for 10 rats.
Figure 2 presents the relative liver weight of rats placed on diet formulated with catfish exposed to biodiesel. Although the value of relative liver weight of test rats was numerically lower than that of control rats, there was still no significant difference between the control and the test rats (p>0.05).
The serum levels of some indicators of liver function were monitored. Table 3 presents the results of serum concentrations of bilirubin, albumin and globulin of rats placed on feed formulated with catfish exposed to biodiesel over a period of 30 days. Generally, it was observed that concentrations of direct and total bilirubin of rats increased significantly (p<0.05) as the level of biodiesel increased while the serum concentrations of albumin and globulin decreased significantly (p<0.05) with increasing level of biodiesel. The concentration of direct bilirubin in BD 0.25 rats was about 2 folds that of the control. There was no significant difference (p>0.05) between the concentration of serum total bilirubin of BD 0.1 rats and the control, while that of BD 0.25 rats was significantly higher than that of control (p<0.05). The serum globulin concentration of control rats was about 1½ fold that of the BD 0.25 and BD 0.1 rats.
| Group of Rats | Bilirubin | Albumin (g/dL) | Globulin (g/dL) | |
| Direct (mM/L) | Total (mM/L) | |||
| CONTROL | 1.04±0.02a | 5.45±0.21a | 16.89±1.35a | 10.54±1.09a |
| BD0.1 | 1.26±0.02b | 6.02±0.52ab | 14.66±1.64b | 7.72±1.30b |
| BD0.25 | 1.94±0.02c | 6.56±0.64b | 12.31±1.55b | 6.48±0.97b |
Table 3: Serum concentration of selected liver function indicators of rats placed on diet formulated with catfish exposed to biodiesel polluted water over a period of 30 days.
Figure 3 presents the level of serum malondialdehyde (MDA) of rats placed feed formulated with catfish exposed to biodiesel. The general trend observed was that serum level of MDA increases significantly (p<0.05) with increasing level of biodiesel. The serum MDA levels of test rats were about 1 ½ fold that of the control.
Results of concentration of serum reduced glutathione (GSH) of rats placed on diet formulated with catfish exposed to biodiesel over a period of 30 days is presented in Figure 4. Here, concentration of serum GSH of test rats was significantly lower than that of the control (p<0.05). GSH concentration of serum of control rats was twice that of the rats placed on diet formulated with catfish exposed to biodiesel.
Specific activities of selected liver enzymes of rats exposed to biodiesel were assayed and the result is presented in Table 4. Specific activities of alanine transaminase (ALT), asparatate transaminase (AST) and Gama glutamyltranspeptidase (GGT) of serum of BD 0.1 and BD0.25 rats were not significantly different (p>0.05) but significantly lower than those of control rats. Conversely, specific activities of catalase (CAT), superoxide dismutase (SOD) and glutathione-stransferase (GST) of serum of BD 0.1 is significantly higher than that of BD 0.25 (p<0.05) while both were significantly lower than that of the control rats (p<0.05). Generally, specific enzyme activities of the selected enzymes decreased as the level of biodiesel increased.
| Group of Rats | AST | ALT | GGT | CAT | SOD | GST | |
| CONTROL | 24.9±1.19a | 14.6±1.64a | 67.9±4.36a | 0.82±0.03a | 1.21±0.02a | 1.32±0.02a | |
| BD0.1 | 18.8±1.94b | 10.4±1.35b | 53.2±4.26b | 0.61±0.02b | 0.92±0.01b | 0.88±0.02b | |
| BD0.25 | 15.2±1.69b | 10.0±1.89b | 47.3±4.90b | 0.55±0.03c | 0.81±0.02c | 0.56±0.04c | |
Table 4: Specific activity (U/mg protein) of selected enzymes of the liver of rats placed on diet formulated with catfish exposed to biodiesel polluted water over a period of 30 days.
The current study demonstrated that catfish exposed to biodiesel from PKO significantly affects function and oxidative status of liver of albino rats. This is the first investigation to report that catfish exposed to biodiesel from PKO decreased liver antioxidant status of albino rats. In addition, these data are the first to demonstrate liver dysfunction of albino rats fed with catfish exposed to environmentally relevant concentration of biodiesel from PKO.
The performance of rats placed on the formulated diets as reported in Table 1 suggested that the biodiesel from PKO decreased the nutrient in the catfish exposed to it thereby increasing the FCR. It could also be that the albino rats could not convert the catfish exposed to biodiesel from PKO into useful nutrients for body growth and development creating a situation where quantity of food consumed by the animals was not commensurate with body weight eventhough positive growth response was observed (Figure 1). A number of studies have demonstrated positive growth response to consumption of contaminated water and food from contaminated water bodies [31-33]. Data from this study are in agreement with these earlier findings.
Since organ weight evaluation is an essential part of the toxicologic and risk assessment of drugs, chemicals, biologics, food additives, and medical devices, the relative liver weights of the experimental rats were determined (Figure 2). These results revealed that consumption of fish exposed to biodiesel form PKO did not significantly affect relative liver weights of albino rats. However, Haley et al. [34] reported that organ weight changes themselves were not necessarily toxic effects and emphasized that the organ weight data should be assessed in the context of the entire study. Therefore, the data on relative liver weight may not imply non-toxic effect.
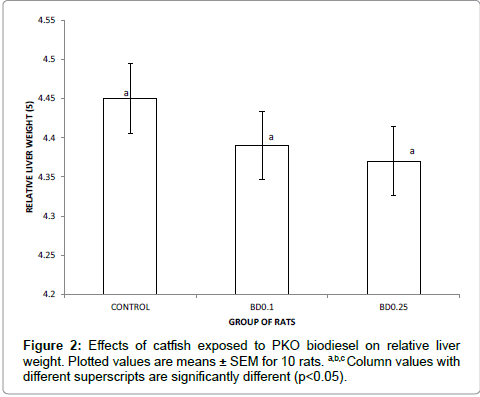
Figure 2: Effects of catfish exposed to PKO biodiesel on relative liver weight. Plotted values are means ± SEM for 10 rats. a,b,c Column values with different superscripts are significantly different (p<0.05).
Bilirubin is an endogenous compound that can be toxic [35], especially in neonates. Bilirubin is the ultimate breakdown product of haemoglobin and serves as a diagnostic marker of liver and blood disorders. Elevated serum bilirubin of rats observed in this study (Table 2) suggested liver dysfunction which may not be unconnected with catfish exposed to biodiesel from PKO. The liver is responsible for clearing the blood of bilirubin. It does this by the following mechanism: bilirubin is taken up into hepatocytes, conjugated (modified to make it water soluble) and secreted into the bile, which is excreted into the intestine. Increased total bilirubin causes jaundice and can signal a number of problems. Studies had shown that if direct (that is, conjugated) bilirubin is normal, then the problem is an excess of unconjugated bilirubin and the location of the problem is upstream of bilirubin excretion. Anemia, viral hepatitis, or cirrhosis can be suspected. If direct bilirubin is elevated, then the liver is conjugating bilirubin normally, but is not able to excrete it. Bile duct obstruction by gallstones or cancer should be suspected [17]. In this study, rats fed with catfish exposed to PKO biodiesel presented elevated serum concentrations of both total and direct bilirubin suggesting that the liver is not able to excrete bilirubin which is an evidence of liver dysfunction.
Serum albumin and globulin concentrations are some biochemical indices for monitoring liver function in the blood (Table 2). Abnormal levels of these proteins have been reported to be associated with haemolysis or increased breakdown of RBC and/ or liver damage [36]. Decreased serum concentrations of albumin and globulin as observed in this study lend credence to the submission that the liver function may be impaired. Both globulin and albumin are produced by the liver. If the liver is damaged, it may no longer be able to produce these proteins. The results presented on serum proteins are consistent and all pointing to the fact that the liver may have been damaged by consumption of the contaminated fish.
Hepatic ALT, AST, GGT, GST, CAT, SOD, GSH and lipid peroxidation product (malondialdehyde, MDA) are commonly used as biomarkers of hepatotoxicity [37]. Abnormal values of serum GGT, ALT and AST have been observed for different diseased conditions, but most especially in hepatic diseases (Adeyemi et al., 2009). The experimental data (Tables 3) revealed that the catfish exposed to PKO biodiesel might have altered protein metabolism, among others, at the subcellular level and this may be indicative of impairment of the function of the tissues.
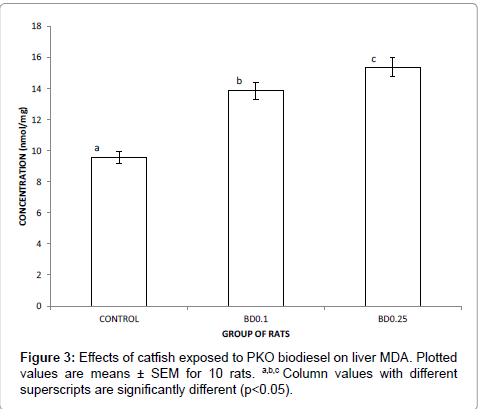
Figure 3: Effects of catfish exposed to PKO biodiesel on liver MDA. Plotted values are means ± SEM for 10 rats. a,b,c Column values with different superscripts are significantly different (p<0.05).
Ecological stressor such as biodiesel generate reactive oxygen species (ROS) such as O2-, H2O2 and OH radicals have potential to interact with many cellular components, causing significant damage to membrane and other cellular structures [38]. Some of the ROS are highly toxic and must be detoxified by cellular responses [39]. The ROS scavenging depends on the detoxification mechanism, which may occur as a result of sequential and simultaneous action of a number of antioxidant enzymes, including superoxide dismutase (SOD), catalase (CAT), and glutathione-S-transferase (GST). The observed decreased activity of SOD, CAT and GST in the liver of rats fed with catfish exposed to PKO biodiesel was indicative of oxidative stress.
Malondialdehyde (MDA) is a lipid peroxidation product generated from reactive oxygen species (ROS), and as such is assayed in vivo as a bio-marker of oxidative stress [40]. Malondialdehyde reacts with deoxyadenosine and deoxyguanosine in DNA, forming DNA adducts, malondialdehyde is reactive and potentially mutagenic. It has been found in heated edible oils such as sunflower and palm oils. The significant increase in the levels of MDA of liver of experimental rats (Figure 3) lend credence to the view that biodiesel caused a reduction in the total antioxidant status by reactive oxygen species.
GSH plays a very important role in the detoxification of xenobiotics. In vitro examinations proved that the free thiol group of glutathione reacts with xenobiotics to form conjugates. These conjugates reveal toxic properties [41]. In this study, catfish exposed to PKO biodiesel quickly depletes hepatocyte glutathione levels (Figure 4). It is therefore, a potential agent to which can lead to further lipid peroxidation.
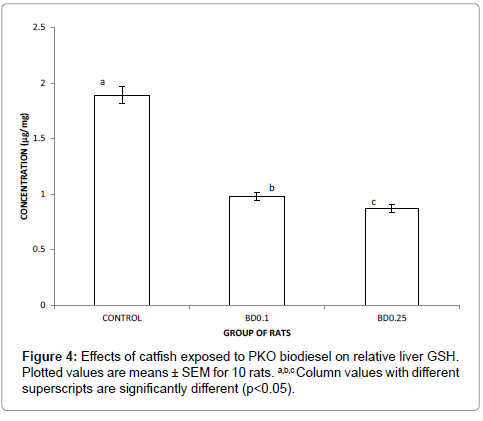
Figure 4: Effects of catfish exposed to PKO biodiesel on relative liver GSH. Plotted values are means ± SEM for 10 rats. a,b,c Column values with different superscripts are significantly different (p<0.05).
In conclusion, the author has shown evidence that consumption of catfish exposure to PKO biodiesel alters growth and performance of albino rats. The indices of liver function observed, including serum levels of bilirubin, albumin, globulin, and activities of AST, ALT, GGT have not been reported previously in investigations of the effects of catfish exposed to PKO biodiesel on albino rats. The data on MDA, GSH and antioxidant enzymes (CAT, SOD, GST) of the liver of experimental rats suggests that consumption of catfish exposed to PKO biodiesel may predispose the organ to a condition of oxidative stress. Future study should consider light micrograph of the liver.
The author declares that there are no conflicts of interest.
The author acknowledges Prof O.B Oloyede and Prof (Mrs) A.T. Oladiji, Department of Biochemistry, University of Ilorin, for their assistance.