Journal of Clinical Trials
Open Access
ISSN: 2167-0870
ISSN: 2167-0870
Commentary - (2014) Volume 4, Issue 1
The normal heart beat intervals are neither strictly stationary nor completely random, and continuously shift from one period to another. Decoding the ECG identifies this “hidden” information that imparts inherent complexity to the heart-beating interval time series. Loss of this complexity in cardiovascular disease is manifested as a reduction in Heart Rate Variability (HRV) and this reduction correlates with an increase in both morbidity and mortality. Because HRV measurements are noninvasive and easy to perform, they have emerged as an important tool in cardiology. However, the identities of specific mechanisms that underline the changes in HRV that occur in cardiovascular diseases remain largely unknown. Changes in HRV have mainly been interpreted on a neural basis, i.e. due to changes in autonomic impulses to the heart: sympathetic activity decreases both the average heart beat interval and HRV, and parasympathetic activity increases both. It has now become clear, however, that the heart rate and HRV are also determined by intrinsic properties of the pacemaker cells that comprise the sinoatrial node, and the responses of these properties to autonomic receptor stimulation. Here we review how
changes in the properties of coupled-clock mechanisms intrinsic to pacemaker cells that comprise the sinoatrial node and their impaired response to autonomic receptor stimulation are implicated in the changes of HRV observed in heart diseases.
Keywords: Cardiac denervation; Coupled-clock pacemaker system; Fractal-like behavior of the heart rate
The normal heart beat intervals are neither strictly stationary nor completely random (i.e., chaotic), and continuously shift from one period to another. This chaotic heart rhythm is due to nonlinear oscillators interacting together in a complex dynamic [1]. Decoding the ECG identifies this “hidden” information that imparts inherent complexity to the heart beating interval time series by revealing the existence of fractal-like dynamic behaviors that operate over multiple time scales. Loss of this complexity in cardiovascular disease is manifested as a reduction In Heart Rate Variability (HRV), and this reduction correlates with an increase in both morbidity and mortality [2]. Because HRV measurements are noninvasive and easy to perform, they have emerged as an important tool in cardiology. However, the identities of specific mechanisms that underline the changes in HRV that occur in cardiovascular diseases remain largely unknown. Changes in HRV have mainly been interpreted on a neural basis, due to modification of autonomic impulses to the heart: sympathetic activity decreases both the average heart beat interval and HRV, and parasympathetic activity increases both (Figure 1). Therefore, an imbalance of flux between the two arms of the autonomic system in the presence of cardiovascular disease has been thought to be the basis of changes in both the heart beat interval and HRV. The Sinoatrial Node (SAN) is the tissue area within the heart from which the cardiac impulse originates. Because parasympathetic and sympathetic nerves discharge neurotransmitters that bind to β-adrenergic or cholinergic receptors of pacemaker cells within the SAN tissue and modulate the heart rate and rhythm, the readout of HRV is a direct output of pacemaker cell function. Specifically, experimental evidence [3-5] has indicated that the graded changes in the rate of action potential firing by the SAN are nonlinear functions of the graded autonomic receptor stimulation, indicating that intrinsic properties of pacemaker cells within the sinoatrial node are not only determinants of heart rate, but also are important determinants of HRV [6]. Cardiac control by autonomic neural impulses is much more complex than only sympathetic and parasympathetic nerve stimulations. For example, in the recent years the complex role of intrinsic cardiac ganglia in maintaining the adequate cardiac output has been revealed [7,8]. The average number of ganglia per porcine is in the range of 350, and the majority of which are distributed in the atria with a smaller part is located in the ventricular. Moreover, it was shown recently that these cardiac ganglia can directly affect SAN function [9]. In this paper we describe the basic methods for assessment of HRV, discuss novel perspectives on the dynamic of the coupled nonlinear oscillators intrinsic to pacemaker cells residing within the SAN, and how signaling via autonomic receptors on pacemaker cells links neural impulses to the intrinsic pacemaker cell signaling pathways to alter normal automaticity. Finally, we hypothesize and provide evidence how deficits in intrinsic regulatory properties of pacemaker cells during cardiac disease can affect heart rate and HRV.
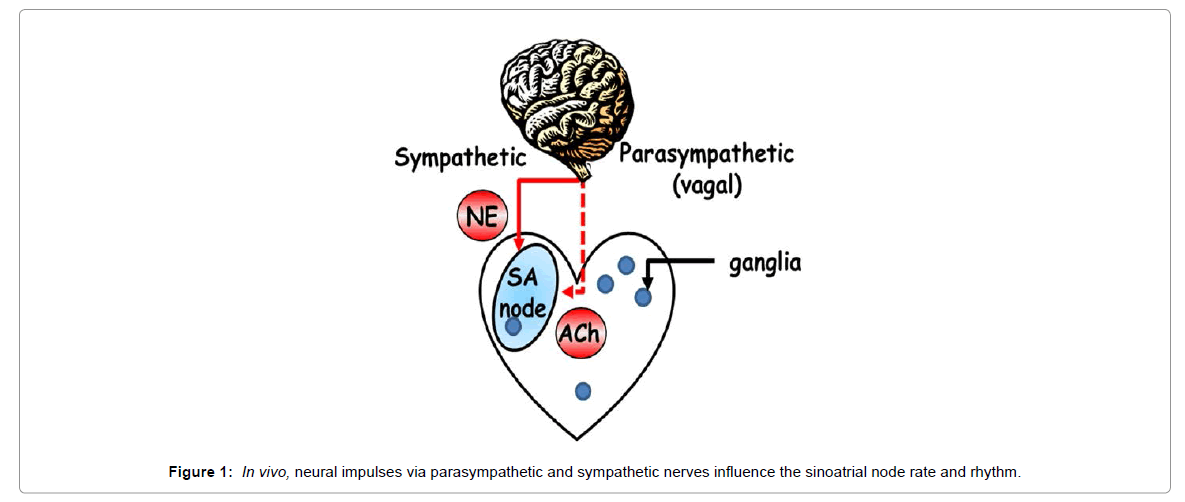
Figure 1: In vivo, neural impulses via parasympathetic and sympathetic nerves influence the sinoatrial node rate and rhythm.
HRV analytic methods are used to quantify the statistical variability of the adjacent intervals in the ECG recordings in vivo, electrical activity in SAN tissue, or action potentials in single pacemaker cells [10]. The major methods for quantifying HRV are categorized as time domain, frequency domain, geometric, and nonlinear. Time-series analysis is the simplest means of evaluating HRV to identify variation in beat-to-beat intervals over time: 1) SDNN, standard deviation of all beat intervals; 2) SDANN, standard deviation of 5-minute average beat intervals; 3) RMSSD, square root of the mean of the squares of successive beat intervals; 4) pNN50, the percentage of intervals with 50 ms or more different from the preceding interval; 5) CV, coefficient of variation of all beat intervals.
Fast Fourier transformation of the beat intervals has traditionally been employed to quantify cyclic fluctuations of the beat intervals in the frequency domain at frequencies lower than the heart rate [11]. For this measurement in humans the ECG is usually recorded over periods ranging from 5 min (approximately 256 beats) to 24 hours. For a short ECG recordings (~5 min), only two peaks are present in the beating interval power spectrum: a High Frequency (HF) peak, which in humans is present in the frequency domain between 0.15 and 0.4 Hz (between 2 and 6 heart cycles); and a Low Frequency (LF) peak, which in humans is present between 0.04 and 0.15 Hz (between 6 and 25 heart cycles). Two other power spectrum peaks can be defined within longer ECG recordings: Ultralow-Frequency (ULF), which in humans is present in the segment below 0.0033 Hz (below 300 heart cycles) and Very Low Frequency (VLF), defined in humans as the segment between 0.0033 and 0.04 Hz (between 25 and 300 heart cycles).
Poincaré plot analysis (also called a return map) is a geometrical technique that was first applied by Woo et al. [12]., to beat intervals from patients with cardiac disease. In a Poincaré plot, each beat interval is visualized as a function of the preceding interval (Figure 2). A higher scattering around the mean beat interval implies the presence of a “complex” dynamic within the time-series of beating intervals where a low scattering (e.g., “cigar shape”) implies an absence of a complex dynamic.
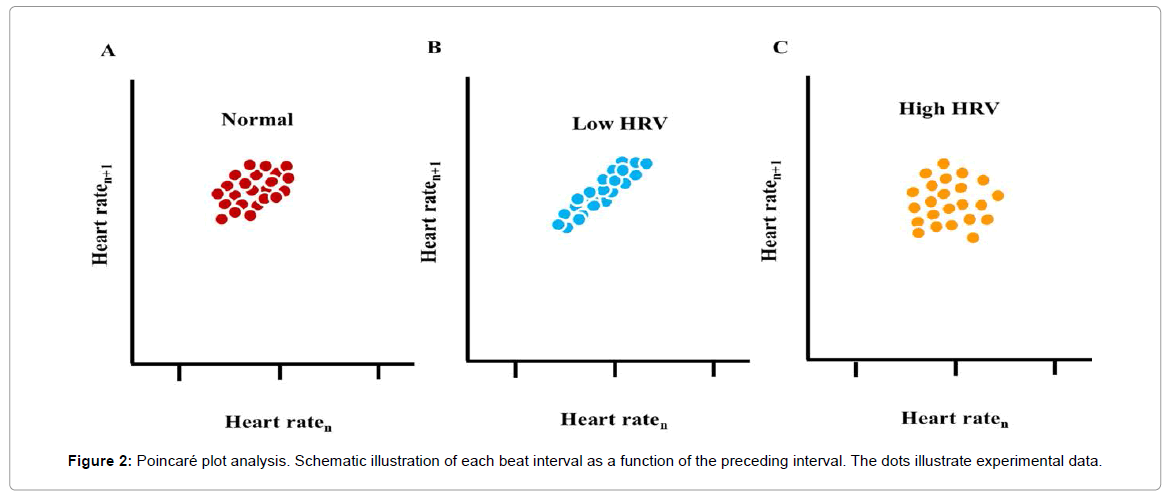
Figure 2: Poincaré plot analysis. Schematic illustration of each beat interval as a function of the preceding interval. The dots illustrate experimental data.
Considering the nonlinear nature of biological systems, nonlinear analyses are required to fully characterize complex dynamics. For example, differences in the time series of heart-rate intervals lie not only in the distribution of the inter-beat variations around the mean heart rate, but also in their time ordering: different dynamic patterns can occur at different frequency scales that correspond to the actual frequency of an underlying physiological process [13]. Although the nature of a specific mechanism that produces this complexity within the heart beat intervals has yet to be discovered, that this complexity has been documented to be reduced in advanced age and in the context of cardiovascular disease may imply the HRV is necessary for healthy heart function. Fractal analysis of the heart beats is a non-linear method to decode the secrets hidden in the heart-beat intervals [14]. One method to identify and quantify fractal-like behavior is the power law function slope (Figure 3). Power spectra that embody fractal-like behavior have a high density of power at low frequencies which exponentially decrease at high frequencies [15]. Plotting this exponential relationship in a loglog scale transforms it to a linear function, the slope of which is defined as β. In a log-log plot of healthy mammals, the power law slope in VLF is linear with a negative slope, β~-1. Short- and long-term fractalscaling exponent quantified by Detrended Fluctuation Analysis (DFA) characterizes the degree of correlation among time scales embedded within the heart beat intervals (Figure 3) [16]. The short-term exponent, α1, is a measure of the degree to which the beat intervals are correlated on a scale of 3-11 beats, and the long-term exponent, α2, is a measure of the degree to which the beat intervals are correlated on a scale of 12-20 beats. Note that if β or α is less than 0.5, the system dynamic is described as white noise (i.e., a random signal with a constant power spectral), and if β or α equals 1.5, the system dynamic is described as Brownian noise (i.e., an integral of white noise that has more energy at lower frequencies).
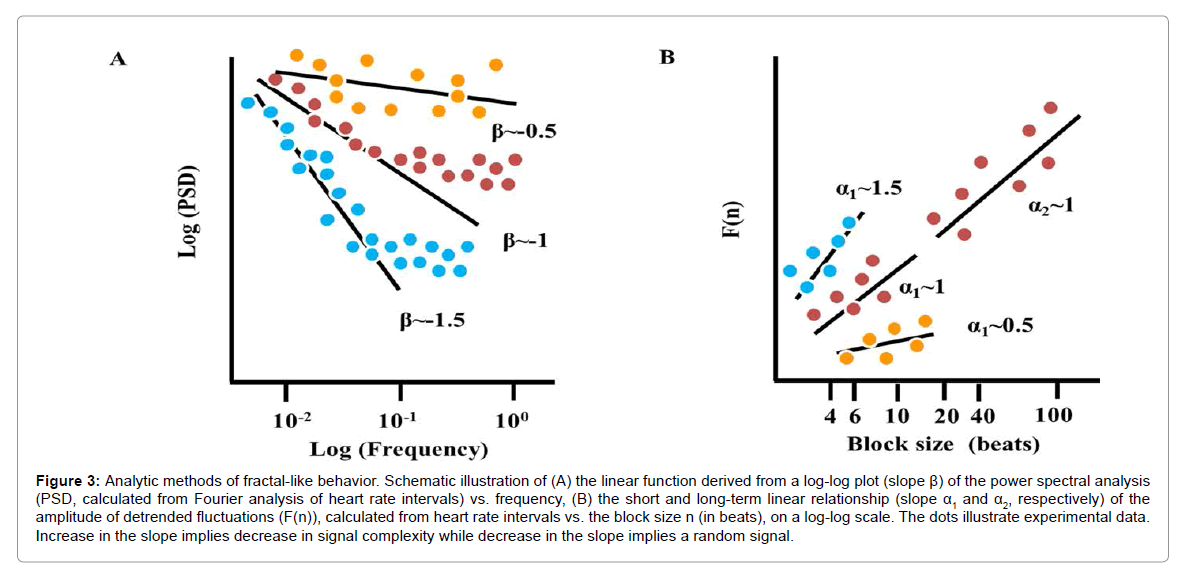
Figure 3: Analytic methods of fractal-like behavior. Schematic illustration of (A) the linear function derived from a log-log plot (slope β) of the power spectral analysis (PSD, calculated from Fourier analysis of heart rate intervals) vs. frequency, (B) the short and long-term linear relationship (slope α1 and α2, respectively) of the amplitude of detrended fluctuations (F(n)), calculated from heart rate intervals vs. the block size n (in beats), on a log-log scale. The dots illustrate experimental data. Increase in the slope implies decrease in signal complexity while decrease in the slope implies a random signal.
Assessment of approximate entropy is another nonlinear method to assess HRV, and it is used to quantify the regularity and complexity of time-series data. Larger values of entropy correspond to greater apparent randomness or irregularly, whereas smaller values correspond to instances of recognizable (i.e., Brownian Noise) within patterns in the data. That the level of approximate entropy is lower than apparent randomness, but higher than recognizable patterns, may imply that the healthy heart operates with optimum entropy efficiency.
The combination of functional or structural indices such as the left ventricular ejection fraction with indices derived from HRV analysis has been proposed as a useful and reliable method for risk assessment among patients with heart disease [17]. In a random effects meta-analysis, a low SDNN was associated with an increase of a first cardiovascular event in populations without known cardiovascular disease [18]. Moreover, patients with a high cardiovascular risk profile have lower time-domain HRV indices, and a reduced LF power and LF:HF ratio [19]. Finally, entropy generally decreases with cardiac vascular disease risk progression [20]. Identification of the mechanisms for changes in HRV will enable the design of specific novel treatments for different cardiac diseases. Evidence from in vivo and ex vivo experimental studies suggests that HRV is not only dependent upon the autonomic nervous system impulse balance, but may also depend on intrinsic mechanisms of SAN tissue: 1) even in the presence of complete denervation, the fractal-like behavior of the heart in vivo is still present [21], although the fractal-like slope (β) is reduced; 2) in isolated hearts (i.e., when the heart is completely detached from both hormonal and neural input), there is a non-linear relationship between the HRV and heart beating rate in numerous species [22]; 3) fractal-like behavior is present in monolayers cultured cells with pacemaker-like activity [23].
The SAN is an anatomically and electrophysiologically heterogeneous structure; its pacemaker cells express a unique set of ion channels and Ca2+ proteins necessary for the generation and propagation of electrical activity in the heart. Recent evidence [24] shows that the spontaneous beating of pacemaker cells isolated from the SAN, even in the absence of receptor stimulation by neuronal input, is driven by Ca2+ activation of calmodulin-adenylyl cyclase (AC)- dependent protein kinase A (PKA) and Ca2+/calmodulin-dependent protein kinase II (CaMKII) which effect phosphorylation of proteins that drive biophysical mechanisms leading to coupling of clocks within pacemaker cells (Figure 4) [25]: the Sarcoplasmic Reticulum (SR) is a Ca2+ clock that spontaneously generates chaotic Local Diastolic Ca2+ Releases (LCRs) that activate an inward Na+-Ca2+ exchanger current, which accelerates the rate of diastolic membrane depolarization. Na+- Ca2+ exchange current and the f-channel current, another member of the ensemble of sarcolemmal electrogenic molecules (“membrane clock”), concurrently drive the diastolic membrane depolarization to ignite the next Action Potential (AP). This spontaneous diastolic phase is a unique feature of the pacemaker cells that is also functional in other peacemaking tissue of the heart (i.e. atrioventricular node, bundle of His, bundle branches, and His-Purkinje system). During late diastolic depolarization L-type channels are activated and Ca2+ influx via these channels induce Ca2+ release from the SR via ryanodine channels. This released Ca2+ binds to calmodulin that activates AC, inactivates L-type channel and modulates the Na+-Ca2+ exchange current, contributing to the Ca2+ transient decay by causing Ca2+ efflux from the cell. A substantial component of Ca2+ influx via the L-type channels is pumped directly into the SR, and replenishes cell Ca2+ load to balance Ca2+ efflux via Na+-Ca2+ exchange. The occurrence of AP-triggered Ca2+ release synchronizes the depleted SR Ca2+ state that temporarily suspends generation of spontaneous LCRs. Moreover, membrane depolarization during the AP activates K+ channels, which effect AP repolarization.
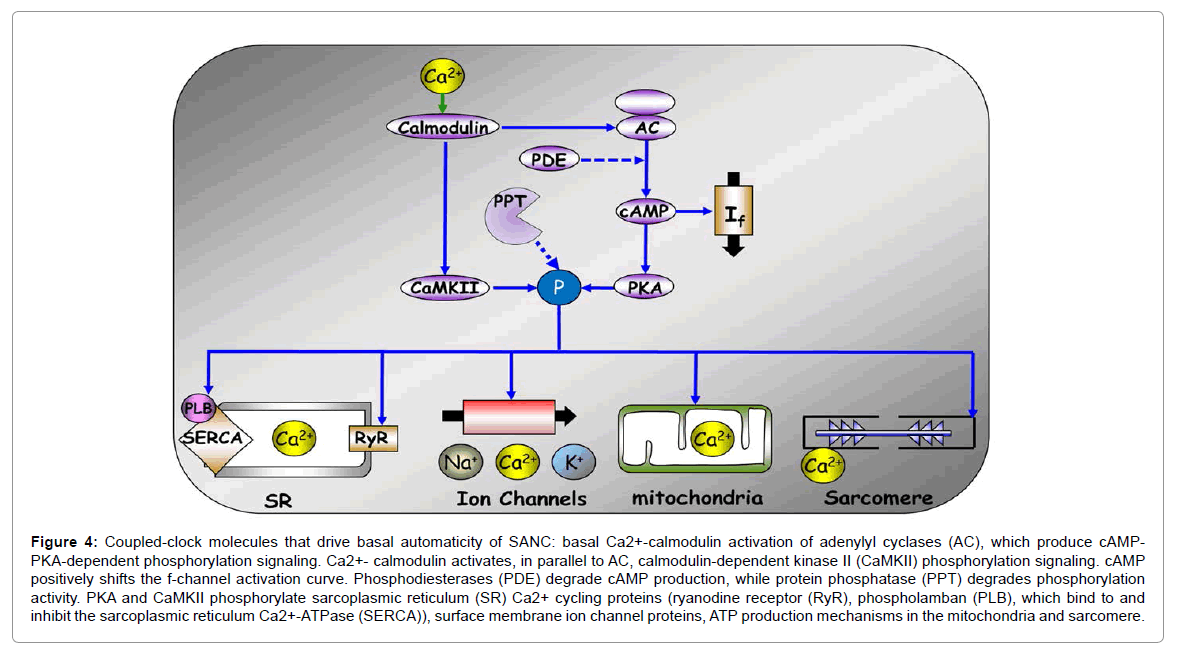
Figure 4: Coupled-clock molecules that drive basal automaticity of SANC: basal Ca2+-calmodulin activation of adenylyl cyclases (AC), which produce cAMPPKA- dependent phosphorylation signaling. Ca2+- calmodulin activates, in parallel to AC, calmodulin-dependent kinase II (CaMKII) phosphorylation signaling. cAMP positively shifts the f-channel activation curve. Phosphodiesterases (PDE) degrade cAMP production, while protein phosphatase (PPT) degrades phosphorylation activity. PKA and CaMKII phosphorylate sarcoplasmic reticulum (SR) Ca2+ cycling proteins (ryanodine receptor (RyR), phospholamban (PLB), which bind to and inhibit the sarcoplasmic reticulum Ca2+-ATPase (SERCA)), surface membrane ion channel proteins, ATP production mechanisms in the mitochondria and sarcomere.
The rate at which this intrinsic coupled-clock system ticks becomes further accelerated, or reduced, respectively, by adrenergic or cholinergic autonomic receptor stimulation (Figure 5). Other neurotransmitters, (e.g., serotonin, substance P, and opioid peptides), hormonal influences (e.g., epinephrine, atrial-natriuretic peptide, brain-natriuretic peptide), and mechanical factors (e.g., atrial stretch and arterial pressure) also modulate the in vivo heart rate. Adrenergic autonomic receptor stimulation of isolated pacemaker cells synchronizes intrinsic clock mechanisms via Ca2+ calmodulin activation of AC/PKA and CaMKIIdependent phosphorylation of Ca2+ and membrane proteins [26]. Cholinergic receptor stimulation, in contrast, reduces Ca2+-calmodulin activation of AC/PKA and CaMKII signaling and desynchronizes intrinsic clock mechanisms [27].
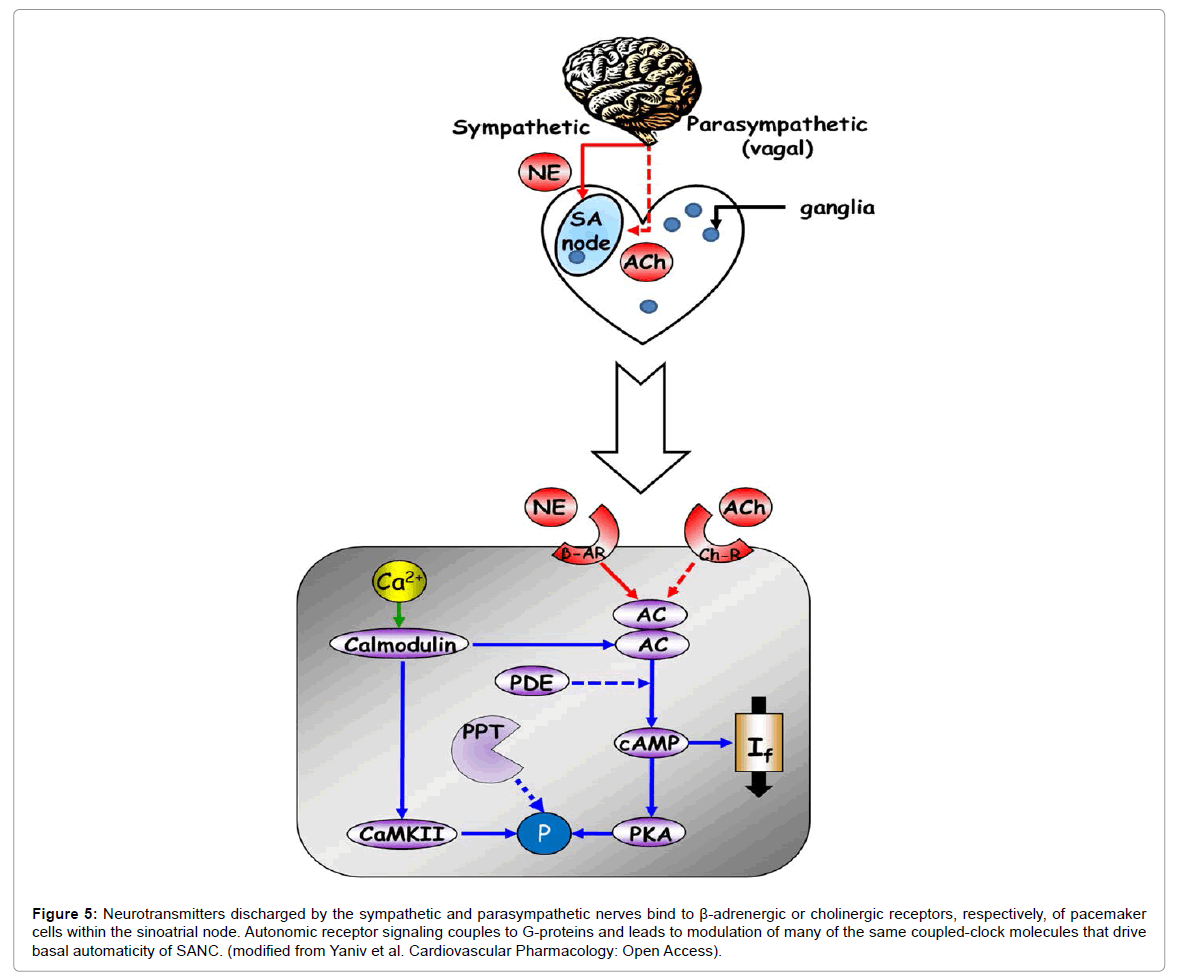
Figure 5: Neurotransmitters discharged by the sympathetic and parasympathetic nerves bind to β-adrenergic or cholinergic receptors, respectively, of pacemaker cells within the sinoatrial node. Autonomic receptor signaling couples to G-proteins and leads to modulation of many of the same coupled-clock molecules that drive basal automaticity of SANC. (modified from Yaniv et al. Cardiovascular Pharmacology: Open Access).
Variations in the coupled-clock function of pacemaker cells residing in the SAN lead to variation in the rate of action potential firing and can induce scale-invariant dynamics in the cardiac impulse, thus imparting variability in the heart rate and rhythm measured by the ECG in vivo. As evidence, in isolated rabbit SANC, beat-to-beat variations in the spontaneous AP firing rate are directly correlated with average variations in the period of local Ca2+ releases that are controlled by the coupled-clock system period [5]. However, it is not known whether or how the pacemaker cells contribute to HRV in vivo [28,29]. Here we discuss evidence that impaired intrinsic properties of pacemaker cells becomes manifest in an altered heart rate and HRV in the context of heart disease, the main causes of morbidity and mortality in developed countries [30].
Heart failure
Heart failure is a highly prevalent syndrome that affects close to 6 million adults in the US alone, with 600,000 new cases diagnosed each year [31]. Patients with chronic heart failure have a higher heart rate, reduced SDNN and RMSDD and reduced power within VLF and LF frequency domains [32]. These HRV parameters become further reduced in patients with decompensated chronic heart failure compared to those with stable chronic heart failure [32]. It is particularly noteworthy that in heart failure patients there is an increase of the amplitude of LF in the early stages and a reduction of the LF component at the end stage [33]. Furthermore, in heart failure patients the Poincaré plot reveals fewer complex dynamics [12]. Finally, it has been observed that with heart failure heart beat regularity is decreased and α1 of the DFA analysis is reduced (i.e., fractal-like behavior in heart failure becomes more like Brownian noise) [34].
HRV analysis has been also found to be effective in predicting mortality risk in heart failure patients. An average SDNN<50 ms for more than four weeks in a row is associated with a three-fold increase in mortality risk [35,36]. Moreover, a reduced VLF power is a powerful and independent risk predictor of mortality in these patients [37].
Because an increase in sympathetic and a decrease in parasympathetic activity have been documented in heart failure patients [38], it has been assumed that a modification in the autonomic nervous impulse to pacemaker cells is the major reason for changes in HRV. However, changes in intrinsic mechanisms in pacemaker cells have also been documented. In single pacemaker cells (i.e., the autonomic nervous system is not intact) from heart failure rabbits the intrinsic beat interval increases compared to the control. Moreover, membrane ionic channel remodeling accompanies heart failure: both If and Iks densities become reduced [39]. Finally, in SAN of rabbits with heart failure, the bradycardiac response to acetylcholine is much higher, while the response to sympathertic activity is preserved [40]. Zaza et al. [4], however, have shown that when the β adrenergic receptor is stimulated, the spontaneous AP firing rate in isolated SANC increases and the HRV becomes reduced. That the relationship between HRV and the beating rate is non-linear indicates that changes in spontaneous beating interval in single isolated SANC are not simply surrogates of autonomic receptor stimulation. Therefore, an increase in sympathetic activity during heart failure not only increases the beating rate in pacemaker cells residing in the SAN tissue, but also likely reduces the HRV. In other terms, changes in the properties of cells that comprise the sinoatrial node are also likely implicated in the reduction of HRV observed in heart failure patients.
Note that catecholamine levels are enhanced in heart failure patients and long-term activation of adrenergic receptor on pacemaker cells augments the severity of symptoms accompanying heart failure [41]. The response to this activation is blunted due to desensitization of the β receptors [42]. Therefore, the modulation of intrinsic mechanisms in pacemaker cells is also blunted and both components can affect long term changes in HRV.
Myocardial ischemia
Myocardial ischemia is associated with sinus bradycardia (i.e., a sinous node beating rate of less than 50 bpm). Myocardial ischemia is present in many cardiac diseases and its early detection is of substantial concern. Compared to patients without ischemia, night time α1 was significantly lower (fractal-like behavior became more like white noise) than during the daytime in patients with postoperative myocardial ischemia [43]. Because ischemia impairs heart beat initiation within the sinoatrial node, due to a failure of its resident pacemaker cells [44], changes in the properties of pacemaker cells are also likely implicated in the change of HRV and fractal-like slope in the context of myocardial ischemia. Although changes in If, IKr and ICa,L in single isolated pacemaker cells are not involved in the ischemia-induced increase in beating interval, reductions in ICa,T and INCX substantially contribute to a reduced rate of diastolic depolarization that underlies an increase in the beating interval.
Myocardial infarction
In patients with a previous myocardial infarction, both the time and frequency domain measures of HRV are reduced, but the approximate entropy is significantly higher than in healthy subjects [45]. Moreover, compared to healthy subjects, the slope fractal-like behavior within the heart-beat intervals is somewhat steeper in patients who have sustained a myocardial infarction [21]. Time and frequency domain HRV analyses indicate that VLF power of HRV is the strongest independent predictor of ventricular tachycardia in patients that had sustained prior myocardial infarction [46]. Interestingly, although different parameters in both time and frequency domains can predict mortality of patients following acute myocardial infarction [47], the fractal-like short-term scaling exponent (α1) of the heart-beat interval is a better predictor of death than the traditional measures of heart rate variability [48]. Because parasympathetic tone decreases after acute myocardial infarction and the balance of sympathetic to parasympathetic tone increases [49], it has been assumed that modification in the autonomic nervous system impulse is the reason for changes in HRV following an infarction. However, because the number of viable pacemaker cell also decreases following myocardial infarction, their absence can contribute to changes in HRV and increase in entropy observed in vivo. In fact, sick sinus syndrome (see below) following an acute myocardial infarction appears to persist chronically in a high number of these patients [50].
Arrhythmia
The occurrence of major arrhythmic events in patients with arrhytmogenic right ventricular cardiomyopathy is associated with a reduced SDNN in the time domain and reduced LF power in the frequency domain [51]. Just prior to an arrhythmogenic event the LF/ HF ratio increases [52]. Because ventricular arrhythmias can induce pH changes and cause reentrant tachycardia, in which the reentry circuit returns to the atrium via accessory AV connections, changes in signaling intrinsic to the sinoatrial node may cause changes in HRV. Moreover, patients with a transplanted heart (i.e., the autonomic nervous system is not intact) have respiratory sinus arrhythmia heart rate fluctuations similar to those in healthy subjects [53].
Brugada syndrome
Patients with Brugada syndrome have mutations in the gene encoding cardiac sodium channels and this disease is the major cause of ventricular tachyarrhtmia in young patients [54]. In these patients SDNN [55] and LF power [56] of beating interval are reduced. A lack of sympathetic drive in the presence of sustained parasympathetic, drive together with a reduction in cAMP level (due to a direct feedback from a reduction in Na that decreases the Ca2+ balance in the cell), were documented in patients with Brugada syndrome [54]. More experimental data on the changes in characteristics of the SAN function in Brugada syndrome are needed to understand the mechanisms behind the reduction in HRV.
Sick sinus syndrome
Sick sinus syndrome is associated with abnormal impulse initiation and propagation from the SAN. Patients affected by this disease have sinus bradycardia, sinus pauses or arrest, atrial chronotropic incompetence, and SAN exit block [57]. Sick sinus node dysfunction occurs most commonly in older patients, but even healthy older patient show higher regularity and altered fractal scaling, consisting of a loss of complex variability [58]. In the sick sinus patients, the average spread around the mean interval (quantified by Poincaré plot) is higher than in control patients [59]. Similar results were documented in dogs with sick sinus syndrome [60]. Finally, the fractal-like behavior of heart-beat intervals is closer to a white noise in sick sinus patients [59].
Genetic manipulation of coupled-clock proteins
Genetic manipulation of protein phosphorylation of intracellular Ca2+ and surface membrane ion channels, (i.e., coupled-clock proteins) also affect heart rate and HRV.
Ankyrins are adapter proteins required for targeting channels and transporters to the cytoskeleton in diverse cells. Mutations of ankyrin-B in humans are linked to sick sinus syndrome [61]. Mice with mutation in ankyrin-based pathways exhibit, in addition to bradycardia, an increased resting heart interval variability in vivo. Pacemaker cells from mice with ankyrin-B deletion mutations have reduced expression of Na+/Ca2+ exchanger, Na+/K+-ATPase and IP3 receptor compared to the wild-type mice [61]. Therefore, a failure of intrinsic coupledclock mechanisms not only causes bradycardia but also increases HRV observed in vivo.
Genetic manipulations of intracellular Ca2+ and surface membrane ion protein phosphorylation of the coupled-clock system induce change in both heart rate and rhythm. It is notable that mice in which genetic manipulation of HCN led to a reduced binding of cAMP to HCN (i.e. pacemaker channels) do not have bradycardia, but do develop recurrent sinus pauses. In patients (HCN4-695X mutant carriers) there is no significant difference in HRV, although heart rate variance is slightly higher in mutant carriers than in normal patients [62]. Similarly mean heart rate of GIRK4-deficient mice that lack cardiac IKACh does not differ from wild-type, but LF and HF powers are reduced in vivo [63]. Ryanodine receptor mutations (R4496C) induced in mice that confer a reduced threshold for ryanodine activation and polymorphic ventricular tachycardia, in response to β-adrenergic stimulation exhibit an increase in heart-beat interval and increased HRV in vivo [64]. Finally, loss of Cav1.3 (CACNA1D) function in mice is associated with both increased beating interval and HRV [65].
Direct pharmacological inhibition of coupled-clock proteins
Pharmacological interventions that impact on intrinsic SANC mechanisms (Figure 6), e.g., intracellular Ca2+ [66], can cause the AP firing rate to become irregular. For example, ivabradine, a new selective If current blocker at the level of SAN, significantly decreases LF: HF ratio in healthy volunteers during tilt and exercise [67]. In heart failure patients, ivabradine not only induces a heart rate reduction but also increases indices of heart rate variability [68]. It is important to note, however, that due to the crosstalk between the membrane and Ca2+ clocks that regulates pacemaker cell function, whenever either the membrane or Ca2+ clock becomes directly perturbed, the other clock is perturbed indirectly. Therefore, due to clock crosstalk, the steady-state bradycardia associated with different HCN4 mutations, ivabradine or Ca2+ regulatory proteins is likely mediated, only in part by If inhibition, and in part, by changes in the Ca2+ clock [69].
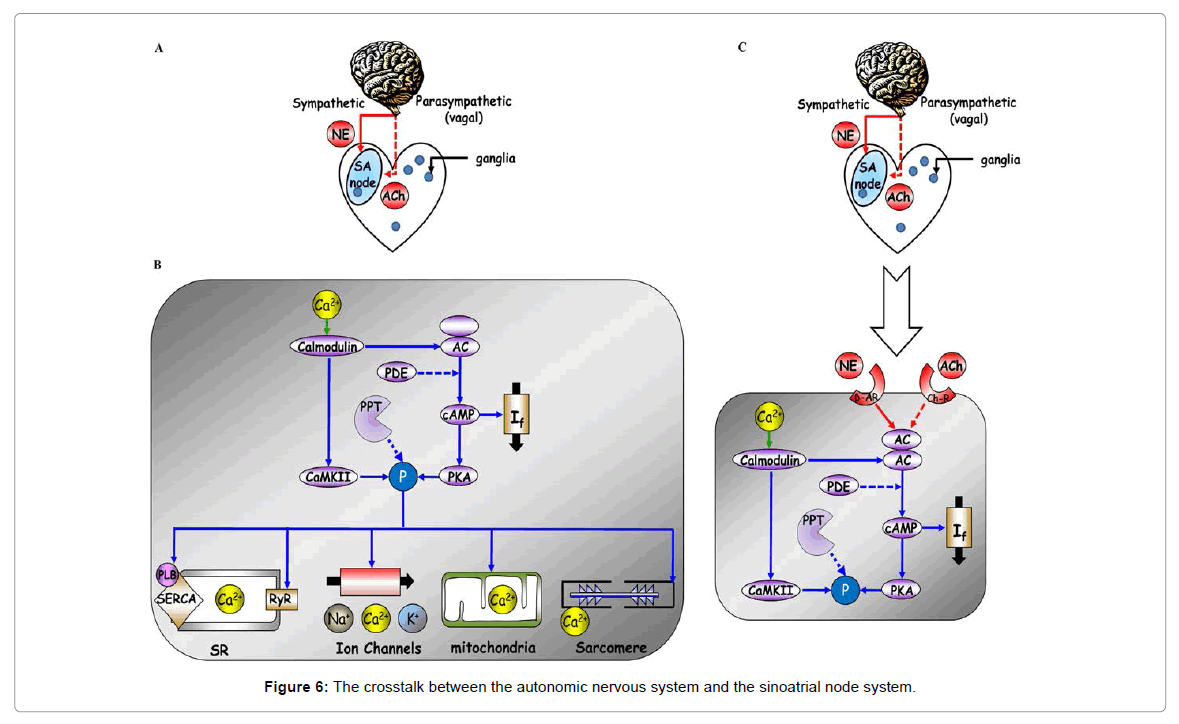
Figure 6: The crosstalk between the autonomic nervous system and the sinoatrial node system.
Aging
Although aging is not a disease per se, it is a complex process that is characterized by a gradual decline in organ functional reserve [70,71]. Moreover, the prevalence of heart diseases described above increases in advanced age. Reductions in HRV [58], entropy [2] and fractal-like slope [38] have been documented in advanced age. Although a decrease in catecholamine responsiveness increased resting levels of sympathetic tone and decreased parasympathetic tone accompany advancing age [72], the age-associated decrease in HRV may, in part, be associated with changes in intrinsic intracellular coupled-clock mechanisms that accompany advanced age. In fact, aging is associated with deficient intrinsic cAMP-PKA-Ca2+ signaling of sinoatrial node pacemaker cell [73]. Therefore, in this case, changes that occur in the properties of the cells in the sinoatrial node likely contribute to age-associated changes in both HRV and fractal-like behavior.
Inflammation
Inflammation is a response to altered homeostasis and occur by releasing different cytokines e.g. tumor necrosis factor α, interleukin-1,4,6,10 [74] from the endocrine. Inflammatory markers, such as cytokines are correlated with increase in mortality and morbidity in patients with cardiac disease [75], and are associated with a reduction in HRV mainly SDNN, VLF power and LF power in patients with coronary artery disease [76], or with heart failure [77]. Inflammation increases both sympathetic and parasympathetic activities but also the oxidative stress [74]. Therefore, increase in oxidative stress can increase CaMKII activity in sinoatrial node cells [78], and thereby affects intrinsic mechanism of the sinoatrial node.
Dietary fish oil
Omega-3 polyunsaturated fatty acid from fish oil decreases heart rate and increases HRV [79]. Interestingly, this supplement reduces the heart rate to the same level in patients with and without vagal innervation [80]. It was shown recently that If is reduced in rabbits that were fed with fish oil diet, indicating a direct effect on properties intrinsic to pacemaker cells [81]. Note, that as mentioned above due to the crosstalk between the membrane and Ca2+ clocks that regulates pacemaker cell function, whenever either the membrane or Ca2+ clock becomes directly perturbed, the other clock is perturbed indirectly. Therefore, due to clock crosstalk, the steady-state bradycardia associated with reduced If current, is likely mediated, only in part by If inhibition, and in part, by changes in the Ca2+ clock [69].
Cervical vagal stimulation
In recent years non-pharmacological interventions that can modulate the cardiac autonomic tone have been suggested for the treatment of heart failure (for review see [82]). Vagal nerve stimulation reduces arrhythmias and increases long-term survival [82]. Vagal stimulation increases parasympathetic tone and VLF power and reduces LF: HF power. The left vagus nerve is associated with cardiac contractility while the right vagus nerve is associated with the sinoatrial node function. Therefore, due to vagal stimulation the properties intrinsic to pacemaker cells change. These results, however, need to be confirmed in larger studies.
Chronic exercise improves heart rate complexity in cardiac disease states
Based on the guidelines for secondary prevention through cardiac rehabilitation in patients with documented coronary disease, an increase in physical activity may reduce mortality [83]. Following exercise training in coronary disease patients, significant increases in SDNN and the HF power and decreases in the LF: HF are generally observed [84]. Note that a decrease in SDNN is associated with increased cardiac mortality (see above). Although exercise training decreases catecholamine levels and β-adrenergic receptor density [84], in heart transplant patients (i.e., the nervous system is not intact) increases in both HRV and the HF power during acute exercise occur that are similar to increases in normal subjects [85]. Similar results were also obtained with pharmacological inhibition of the nervous system [86]. These data suggest that during exercise a non-autonomic mechanism takes the control of HRV.
In summary, during health and cardiac disease, the HRV is determined by intrinsic properties of sinoatrial node pacemaker cells and their modulation by the competing influences of the two branches of autonomic neural input to the heart. A reduced HRV and power within VLF and LF domains has been observed in patients with heart failure, myocardial ischemia or myocardial infraction. Therefore these patients have reduced complex beating interval dynamics. However, an increased HRV and entropy has been observed in patients with arrhythmias, sinus sick syndrome, or mutations in intracellular Ca2+ or surface membrane ion channel protein. Because all these diseases are associated with increased mortality, it seems that the heart in vivo has an optimum level of heart-beat interval complexity, and reduction in this complexity or an increase in the level of disorder are both associated with increased mortality. Finally, from the details above one can conclude that changes in the intrinsic properties of cells that comprise the sinoatrial node are also likely implicated in the changes of HRV and power-law behavior observed in heart failure, ischemia, myocardial infarction and during occurrence of arrhythmia.
Dissection of mechanisms of the HRV at different hierarchical levels (intact heart, denervated heart, isolated sinoatrial node and isolated pacemaker cell) is required for a complete understanding of the mechanisms that regulate HRV in health and induce changes in HRV in the presence of cardiovascular disease. Moreover, an understanding the changes in the intracellular pathways that link autonomic receptors of pacemaker cells to neural impulses from the brain (e.g., phosphorylation signaling, receptor densities, ionic channel proteins) will, in our opinion, be the key to identification of novel therapeutic strategies for treatment of heart disease.
The work was partially supported by the Intramural Research Program of the National Institute on Aging, National Institutes of Health.