Journal of Thermodynamics & Catalysis
Open Access
ISSN: 2157-7544
ISSN: 2157-7544
Editorial - (2015) Volume 6, Issue 1
Understanding the molecular basis of substrate promiscuity shown by some enzymes has been quite elusive. Enzymes that catalyze modification of aminoglycoside antibiotics show a wide range of substrate promiscuity. Comparisons based on amino acid sequence similarity have been more confusing than leading to any explanations. For example, several aminoglycoside modifying enzymes that catalyze different reactions and have less than 5% sequence identity and less than 12% sequence similarity show high overlap in their substrate profiles while some others despite catalyzing the same reaction and having >50% sequence similarity can have significant differences in their substrate profiles [1].
The aminoglycoside-N3- acetyltransferase IIa (AAC-IIa) and the aminoglycoside-N3- acetyltransferase IIIb (AAC-IIIb) are two acetyltransferases that catalyze the acetylation of aminoglycosides at the N3 site of the 2-deoxystreptamine ring [2]. Their amino acid sequence show >65% similarity and they are structurally very similar [3,4]. However, while AAC-IIIb is highly promiscuous and catalyzes acetylation of many aminoglycosides from kanamycin and neomycin group, AAC-IIa is unable to modify neomycin’s [4,5]. Kinetic and thermodynamic properties of enzyme-aminoglycoside complexes are also highly different between the two enzymes [4,5]. NMR studies showed that while AAC-IIIb shows a highly dispersed 15N-1H HSQC spectrum, typical for a protein with a well-defined structure in solution, AAC-IIa yields a highly overlapped spectrum with many missing signals, indicative of a highly flexible protein with unstructured segments [4]. Therefore, in this work, we investigated structural properties of AAC-IIa by analytical ultracentrifugation and differential scanning calorimetry (DSC) to compare with the data acquired with AAC-IIIb in an effort to understand impact of structural differences on their substrate profiles. These studies highlighted significant differences in structural and dynamic properties between these two enzymes.
Unlike AAC-IIIb, DSC scans of AAC-IIa showed multiple transitions (Figure 1A). The formation of the binary enzymeaminoglycoside and the ternary enzyme-aminoglycoside-coenzyme A (CoASH) also showed multiple but different transitions (Figure 1B and C). Specifically, the presence of a low temperature transition was noted which was not detected with AAC-IIIb [6]. Previous work with AAC-IIa was performed at 29°C, which is well above the start of the first transition. Therefore, kinetic and thermodynamic properties of the enzyme was tested at temperatures below 20°C to determine whether the enzyme is more structured at that temperature regime. Another difference between the two enzymes is that the order of ligand addition to form the ternary enzyme-aminoglycoside-CoASH. The melting temperature of the ternary complexes of AAC-IIIb is dependent on the order of ligand addition and varies with each aminoglycoside [6]. There is no such dependence with AAC-IIa.
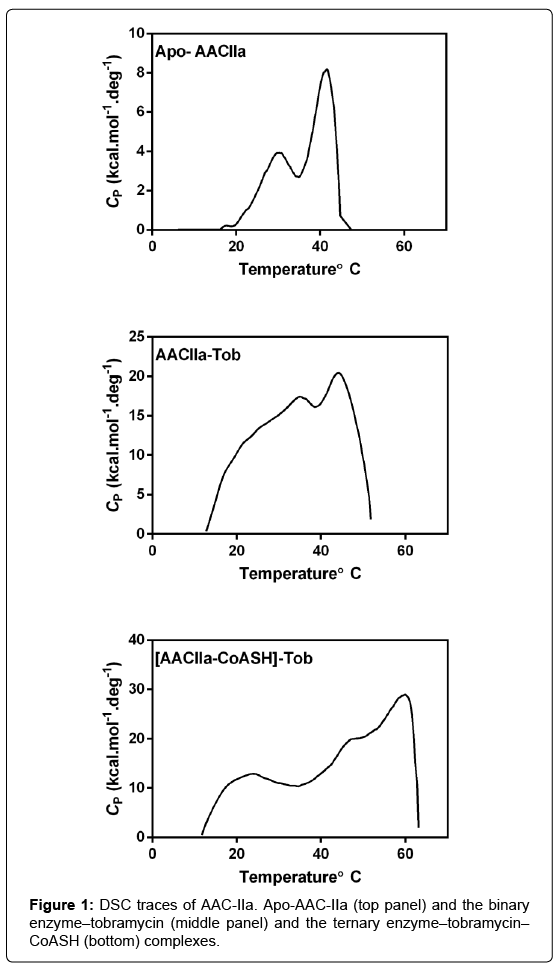
Figure 1: DSC traces of AAC-IIa. Apo-AAC-IIa (top panel) and the binary enzyme–tobramycin (middle panel) and the ternary enzyme–tobramycin– CoASH (bottom) complexes.
NMR studies showed that the spectra acquired at 12°C were still highly overlapped with narrow dispersion indicating that AAC-IIa was still highly flexible even at low temperatures. There was, however, a difference in the spectra acquired with bound ligands. Unlike the behavior of the enzyme at 29°C, both binary complexes, enzymeaminoglycoside and enzyme-CoASH, yielded spectra with only partial increase in resonance dispersion. The formation of ternary complex was necessary to yield spectra similar to those observed for the binary complexes at 29°C. These observations indicate that the enzyme is still flexible at low temperatures, unlike the properties of the enzyme at 29°C, the binding of either ligand alone is not sufficient to yield a welldefined structure in solution.
The differences observed between AAC-IIa and AAC-IIIb are not results of differences in the state of these enzymes in solution; AUC studies showed that AAC-IIIb yields a monodisperse solution and is a homodimer with or without bound ligands. AAC-IIa also yielded monodisperse solutions; however, it showed an interesting and temperature dependent behavior. As shown in (Figure 2) (top panel), the enzyme is a homodimer (59.8 kDa) at temperatures below the first transition and converts to monomer state (30.4 kDa) with bound ligands. Contrary to this, at 29°C, it is always a monomer regardless of the presence of ligands (Figure 2). The only other aminoglycosidemodifying enzyme that showed an aminoglycoside-dependent change in its solution structure is the aminoglycoside nucleotidyltransferase(4′) (ANT) [7]. These observations suggest that the low temperature transition observed with apo-enzyme may represent dissociation of the dimer. However, since transitions at low temperatures are still observable with bound ligands and some even starts at lower temperatures, suggests that multiple events contribute to the observed transitions observed by DSC. These studies strongly suggest that AACIIa is a highly flexible enzyme with or without bound ligands.
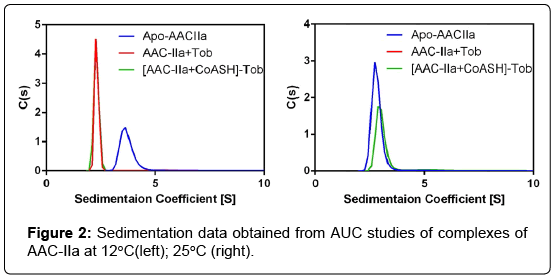
Figure 2: Sedimentation data obtained from AUC studies of complexes of AAC-IIa at 12°C(left); 25°C (right).
As indicated earlier, much less flexible AAC-IIIb can catalyze acetylation of large number of structurally different aminoglycosides, while AAC-IIa is limited to kanamycinlike aminoglycosides. Contrary to this, the aminoglycoside phosphotransferase (3′)-IIIa (APH) is also a highly flexible enzyme and, like AAC-IIa, the apo-enzyme yields NMR spectra typical of intrinsically disordered proteins, which also becomes a welldispersed spectrum by the addition of aminoglycosides, pointing to a highly flexible nature of APH. However, unlike AAC-IIa, APH is probably the most promiscuous aminoglycoside modifying enzyme. These observations also highlight a contrast between flexibility of the enzyme and its substrate profile among the aminoglycoside modifying enzymes. We believe that the effects of ligand binding on the dynamics of the whole protein or its segments may play a more significant role. To this end, binding of CoASH to AAC-IIIb shows that dynamic properties of a large flexible region just above the aminoglycoside binding site is modified significantly by the binding of CoASH despite the fact it CoASH doesn’t interact with this region [8]. Overall, it appears that flexibility alone doesn’t necessarily increase substrate promiscuity of enzymes or even affinity to ligands.