Research Article - (2021)Volume 6, Issue 1
Acute Lymphoblastic Leukemia (ALL) is known to be the most common malignancy among children. The early onset of ALL suggests a key role of genetic factors in its development. Killer Cell Immunoglobulin-like Receptor (KIR) gene complex that encodes a group of key receptors expressed at the surface of natural killer cells has been of great interest as a possible genetic factor affecting the susceptibility to ALL. In this case-control study, we aimed to verify whether the inheritance of specific KIR genes or genotypes were associated with susceptibility to childhood B-ALL in southwestern Iranian population. To this end, KIR genotyping was carried out for 120 patients with childhood B-ALL and 170 unrelated healthy controls using PCR-SSP method. The frequencies of 11 KIR genes and KIR2DS4 variants were investigated among patients and healthy controls. We found no association between KIR genes and genotypes, centromeric and telomeric genotypes as well as the number of activating or inhibitory KIRs and susceptibility to childhood B-ALL. Moreover, no association between childhood B-ALL and KIR B-content scores was detected. However, AA genotype carriers homozygous for KIR2DS4 deleted variant were found to be at a higher risk of developing childhood B-ALL compared with those who inherited at least one KIR2DS4 full-length variant. It seems that the inheritance of different KIR genes does not affect the risk of childhood B-ALL in southwestern Iranian population. However, among individuals with AA genotype, homozygosity for KIR2DS4 deleted allele seems to increase the risk of childhood B-ALL.
Acute lymphoblastic leukemia; Killer cell immunoglobulin-like receptor; AA genotype; Malignancy
Acute Lymphoblastic Leukemia (ALL) is caused by transformation and excessive proliferation of lymphoid progenitor cells [1]. ALL is the most common malignancy among children and childhood ALL accounts for about 80% of all ALL patients [1,2]. ALL can arise from both B cell and T cell precursors. B-ALL comprises about 88% of all of ALL patients. In B-ALL, patient age and WBC count at diagnosis are independent prognostic factors. Patients diagnosed between the ages of 1 and 10 have a better outcome compared with those <1 or = 10 years old [3]. Patients with WBC count = 50,000/mm3 at diagnosis have worse outcome than those with WBC count <50,000/mm3. According to National Cancer Institute/Rome categorization patients between 1-10 years with a WBC count <50,000/mm3 are considered ‘standard risk’ while the remainder are termed ‘high risk’ [4]. Unlike patients with B-ALL, age and WBC count at diagnosis are not considered as determining prognostic factors in patients with T-ALL. The etiology of this leukemia is unknown. Due to the early onset of this malignancy, it is likely that there is a critical role for genetic factors in increasing the risk of ALL. A notable genomic region that has been questioned to have an association with ALL is Killer Cell Immunoglobulin-like receptor (KIR) gene complex. Many studies have been accomplished in order to investigate this association in different populations [5-10]. KIR gene loci, located on chromosome 19q13.4, contain a family of highly homologous genes including 6 activating genes, 8 inhibitory genes and 2 pseudogenes [11]. Due to the polygenic and multi-allelic nature of this genomic region, different individuals may vary in both the number and allele of KIR genes [12]. The diversity in KIR gene repertoire is also reported among different populations [13,14].
KIR genes encode one of the most important receptors expressed at the surface of Natural Killer (NK) cells and some T cells. KIRs have a key role in the function and development of NK cells [15]. The ligands for most of KIR receptors are Huma Leukocyte Antigen (HLA) class I molecules. Viral infection and transformation of cells lead to the down-regulation of MHC class I molecules at the cell surface which can be sensed by NK cells through KIRs, leading to spontaneous destruction of the target cells [16].
There are two major KIR haplotypes, termed A and B. The genes 2DL4, 3DP1, 3DL2, and 3DL3, termed framework loci, are present in all haplotypes. A haplotypes have a fixed gene content consisting of seven genes and two pseudo genes; 2DL1, 2DL3, 2DL4, 2DS4, 3DL1, 3DL2, 3DL3, 2DP1 and 3DP1. Unlike A haplotypes, B haplotypes vary in gene content but must include at least one of the B-specific genes: 2DS1, 2DS2, 2DS3, 2DS5, 2DL2 and 2DL5 [17]. Considering KIR haplotype zygosity, all individuals have either AA genotype or Bx genotype (having one or two B haplotypes).
There are two distinct KIR gene clusters due to linkage disequilibrium. The C4 gene cluster in the centromeric half consists of 2DL2, 2DL5, 2DS2 and 2DS3, and the T4 gene cluster in the telomeric half comprises 2DL5, 3DS1, 2DS1 and 2DS5. According to the presence or absence of C4 and T4 clusters, Bx genotypes are classified as C4T4, C4Tx, CxT4 and CxTx. C4T4 denotes the presence of both C4 and T4 while CxTx represents the absence of both C4 and T4. Further, C4Tx indicates the presence of C4 and the absence of T4; and CxT4 represents the absence of C4 and the presence of T4 [18].
The single activating KIR gene in A haplotype, 2DS4, has a null allele with a 22 base pair deletion in its coding region. Due to the high population frequency of this null allele, it is probable that individuals who are homozygous for A haplotype do not express any activating KIRs.
Although the association between KIR genes and childhood ALL has been studied, due to KIR gene diversity among different populations, we aim to investigate this association in Iranian population.
Study population
We studied 120 children with well-documented B-ALL who were diagnosed based on the standards for ALL diagnosis in the 2008 WHO classification of lymphoid neoplasms [19], along with 170 healthy individuals as controls. The study protocol was approved by our university Ethics Committee (approval number: IR.SUMS. REC.1398.1116). Written informed consent was obtained from all patients’ parents or legal guardians prior to their inclusion in the study. Patients were recruited from Amir Oncology Hospital affiliated with Shiraz University of Medical Sciences (SUMS). We received the remaining blood samples containing EDTA anticoagulant of each patient which were collected for the routine laboratory tests. Genomic DNA was extracted from whole blood using a salting out method [20]. The demographic and laboratory information was extracted from patients’ medical files. DNA samples of 170 unrelated healthy adults with no history of autoimmune diseases and malignancies were obtained from SUMS Immunogenetics Bio bank and used as the control group.
KIR Genotyping
KIR genotyping was performed by polymerase chain reaction using sequence specific primers (PCR-SSP) for 2DL1, 2DL2, 2DL3, 2DL5, 3DL1, 2DS1, 2DS2, 2DS3, 2DS5, 3DS1 genes. Primer sequences were based on those designed [21]. In order to determine KIR2DS4 gene variants, (full-length (fl) and deleted (del), an alternative pair of primers were used based on sequences reported [22]. An additional pair of primers specific for non-polymorphic sequences of the HLA-DRA gene was used in every reaction as internal positive control. DNA samples with known KIR patterns obtained from Institute for Cancer Research, SUMS, were used as positive controls. PCR mixture without template DNA was used as negative control. Total PCR volume for each reaction was 10 μl containing 5 μl of 2x master mix, 5 pmol of each specific primer, 0.5 pmol of each internal control primer, and 30 ng of genomic DNA. DNA amplification was carried out using following condition: initial heating at 95°C for 3 min, then 10 cycles of 20 s at 94°C, 30 s at 65°C and 30 s at 72°C followed by 20 cycles of 20 s at 94°C, 30 s at 61°C and 30 s at 72°C continued with 72°C for 5 min for further extension. PCR products of different KIR genes were verified by agarose gel electrophoresis (2%) containing DNA safe stain. To differentiate between 2DS4 fl and 2DS4 del alleles, electrophoresis was done on 3.5% agarose gel. In order to allow PCR product size identification, 50 bp DNA ladder was loaded in every gel electrophoresis. DNA bands were detected using a gel documentation system.
KIR genotypes were then determined for each sample based on allele frequency database. Centromeric and telomeric KIR genotypes were determined according to the study [23]. We also used the KIR B-content score (which can be vary from 0 to 4) as the number of centromeric and telomeric gene-content motifs containing B haplotype-defining genes [23].
Statistical analysis
To compare KIR gene and genotype frequencies in patient and control groups, two-tailed chi-square with Yates’ correction was applied. Odds Ratio (OR) and 95% Confidence Interval (CI) were calculated for every significant difference. All data were analyzed using Graph Pad Prism version 6 and a P-value<0.05 was considered statistically significant.
Frequency of KIR genes and genotypes in patients and controls
In order to examine whether there is a relationship between KIR genes and childhood B-ALL, we performed KIR genotyping for 120 patients with B-ALL and 170 healthy controls. KIR gene profiles of patients with childhood B-ALL and controls are depicted in Tables. As presented in Table 1, no significant difference was observed in the frequency of KIR genes between patients and controls. There was also no significant difference in the distribution of either AA and Bx genotypes or centromeric and telomeric KIR genotypes between patients and controls (Table 2). No significant difference was detected in B-content scores as well as Bx genotype subsets between patients and controls (Tables 3 and 4).
| B-ALL | Standard-risk B-ALL | High-risk B-ALL | Controls | B-ALL | Standard-risk | High-risk | Standard-risk | |
|---|---|---|---|---|---|---|---|---|
| KIR genes | (n=120) | (n=77) | (n=43) | (n=170) | vs. | vs. | vs. | vs. |
| Controls | Controls | Controls | High-risk | |||||
| N | P-value | |||||||
| % | ||||||||
| 3DL1 | 116 | 73 | 43 | 163 | 0.97 | 0.96 | 0.38 | 0.32 |
| 96.60% | 94.80% | 100% | 95.90% | |||||
| 2DL1 | 120 | 77 | 43 | 170 | >0.9999 | >0.9999 | >0.9999 | >0.9999 |
| 100% | 100% | 100% | 100% | |||||
| 2DL3 | 112 | 71 | 41 | 149 | 0.16 | 0.4 | 0.24 | 0.78 |
| 93.30% | 92.20% | 95.30% | 87.60% | |||||
| 2DS4 | 116 | 73 | 43 | 165 | 0.88 | 0.61 | 0.57 | 0.32 |
| 96.60% | 94.80% | 100% | 97.10% | |||||
| 2DS1 | 34 | 24 | 10 | 65 | 0.1 | 0.35 | 0.1 | 0.48 |
| 28.30% | 31.10% | 23.20% | 38.20% | |||||
| 2DS2 | 54 | 35 | 19 | 93 | 0.13 | 0.23 | 0.29 | >0.9999 |
| 45% | 45.40% | 44.10% | 54.70% | |||||
| 2DS3 | 42 | 27 | 15 | 64 | 0.74 | 0.81 | 0.87 | >0.9999 |
| 35% | 35.00% | 34.80% | 37.60% | |||||
| 2DS5 | 24 | 16 | 8 | 51 | 0.08 | 0.18 | 0.19 | 0.96 |
| 20% | 20.80% | 18.60% | 30.00% | |||||
| 3DS1 | 38 | 24 | 14 | 66 | 0.26 | 0.31 | 0.56 | >0.9999 |
| 31.60% | 31.10% | 32.50% | 38.80% | |||||
| 2DL2 | 54 | 35 | 19 | 92 | 0.15 | 0.26 | 0.32 | >0.9999 |
| 45% | 45.40% | 44.10% | 54.10% | |||||
| 2DL5 | 58 | 37 | 21 | 94 | 0.29 | 0.36 | 0.56 | >0.9999 |
| 48.30% | 48.00% | 48.80% | 55.30% | |||||
Note: P-value was calculated for the presence versus the absence of each KIR gene based on a 2 × 2 contingency table.
Table 1: Frequency of KIR genes in patients with B-ALL compared to the healthy controls.
| B-ALL | Standard-risk B-ALL | High-risk B-ALL | Controls | B-ALL | Standard-risk | High-risk | Standard-risk | |
|---|---|---|---|---|---|---|---|---|
| KIR genotype | vs. | vs. | vs. | vs. | ||||
| (n=120) | (n=77) | (n=43) | (n=170) | Controls | Controls | Controls | High-risk | |
| N | P-value | |||||||
| % | ||||||||
| Cent-KIR genotypes | ||||||||
| AA | 66 | 42 | 24 | 76 | 0.13 | 0.3 | 0.24 | 0.8 |
| 55% | 54.50% | 55.80% | 44.70% | |||||
| AB | 46 | 29 | 17 | 73 | ||||
| 38.30% | 37.60% | 39.50% | 42.90% | |||||
| BB | 8 | 6 | 2 | 21 | ||||
| 6.60% | 7.80% | 4.60% | 12.40% | |||||
| Tel-KIR genotypes | ||||||||
| AA | 80 | 53 | 27 | 101 | 0.45 | 0.27 | 0.4 | 0.17 |
| 66.60% | 68.80% | 62.70% | 59.40% | |||||
| AB | 36 | 20 | 16 | 62 | ||||
| 30% | 25.90% | 37.20% | 36.50% | |||||
| BB | 4 | 4 | 0 | 7 | ||||
| 3.30% | 5.20% | 0% | 4.10% | |||||
| KIR genotypes | ||||||||
| AA | 46 | 30 | 16 | 52 | 0.21 | 0.25 | 0.52 | >0.9999 |
| 38.30% | 38.90% | 37.20% | 30.60% | |||||
| Bx | 74 | 47 | 27 | 118 | ||||
| 61.60% | 61.00% | 62.70% | 69.40% | |||||
Abbreviations: Cent: Centromeric, Tel: Telomeric.
Table 2: Distribution of KIR genotypes in patients with B-ALL and healthy controls.
| B content score | B-ALL | Standard-risk B-ALL | High-risk | Controls | B-ALL | Standard-risk | High-risk | Standard-risk |
|---|---|---|---|---|---|---|---|---|
| B-ALL | vs. | vs. | vs. | vs. | ||||
| (n=120) | (n=77) | (n=43) | (n=170) | Controls | Controls | Controls | High-risk | |
| N | P-value | |||||||
| % | ||||||||
| 0 | 46 | 30 | 16 | 52 | 0.16 | 0.19 | 0.2 | 0.16 |
| 38.30% | 38.90% | 37.20% | 30.60% | |||||
| 1 | 48 | 31 | 17 | 61 | ||||
| 40% | 40.20% | 39.50% | 35.90% | |||||
| 2 | 20 | 10 | 10 | 41 | ||||
| 16.60% | 12.90% | 23.20% | 24.10% | |||||
| 3 | 6 | 6 | 0 | 16 | ||||
| 5% | 7.80% | 0% | 9.40% | |||||
| 4 | 0 | 0 | 0 | 0 | ||||
| 0% | 0% | 0% | 0% | |||||
Table 3: B-content scores in patients with B-ALL and healthy controls.
| Bx genotype subsets | B-ALL | Controls | P-value |
|---|---|---|---|
| (n=74) | (n=118) | ||
| N | |||
| % | |||
| C4T4 | 8 | 18 | 0.56 |
| 10.80% | 15.30% | ||
| C4Tx | 30 | 38 | |
| 40.50% | 32.20% | ||
| CxT4 | 14 | 28 | |
| 18.90% | 23.70% | ||
| CxTx | 22 | 34 | |
| 29.70% | 28.80% | ||
Table 4: Distribution of Bx genotype subsets in patients with B-ALL and healthy controls.
Distribution of KIR genes and genotypes in B-ALL subgroups
Of 120 patients with B-ALL, 77 patients were classified as “standard-risk group” and 43 patients as “high-risk group”. Our results revealed no significant difference in the frequency of KIR genes between patients in the standard-risk or the high-risk group and controls, as well as between patients with standard-risk and those with high-risk B-ALL (Table 1).
Among 77 patients in the standard-risk group, 30 patients (39%) were found to be carriers of AA genotypes and 47 (61.0%) carriers of Bx genotypes. No significant difference was observed in the distribution of AA and Bx genotypes between standard-risk group patients and controls. Moreover, no significant difference was found in the distribution of centromeric and telomeric KIR genotypes between standard-risk group patients and controls.
Of 43 patients in the high-risk group, 16 (37.2%) were AA and 27 (62.7%) were Bx genotype carriers. No significant difference was found in the distribution of AA and Bx genotypes between highrisk group patients and controls. There was also no significant difference in the distribution of centromeric and telomeric KIR genotypes between high-risk group patients and controls. Furthermore, we found no significant difference in the distribution of AA, Bx, centromeric and telomeric KIR genotypes between these two groups (Table 3).
As presented in Table 4, we observed no significant difference in the distribution of B-content scores between either group of patients and controls as well as between the two groups of patients. Patients in standard-risk or high-risk group were compared with controls and also with each other in terms of the number of aKIR versus iKIR genes. As shown in Table 6, no significant difference was observed.
| Number of aKIRs vs. iKIRs | B-ALL (n=120) |
Standard-risk B-ALL (n=77) |
High-risk B-ALL (n=43) |
Controls (n=170) |
B-ALL | Standard-risk | High-risk | Standard-risk |
|---|---|---|---|---|---|---|---|---|
| vs. | vs. | vs. | vs. | |||||
| Controls | Controls | Controls | High-risk | |||||
| N | P-value | |||||||
| % | ||||||||
| iKIR>akIR | 88 | 53 | 35 | 110 | 0.16 | 0.58 | 0.08 | 0.3 |
| 73.30% | 68.80% | 81.40% | 64.70% | |||||
| aKIR>iKIR | 10 | 8 | 2 | 26 | ||||
| 8.30% | 10.40% | 4.70% | 15.30% | |||||
| iKIR=aKIR | 16 | 6 | 34 | |||||
| 18.30% | 20.80% | 13.90% | 20% | |||||
Table 5: Comparison of the number of activating KIR (aKIR) genes versus inhibitory KIR (iKIR) genes between patients with B-ALL and healthy controls.
KIR2DS4 variants in patients with B-ALL and controls
Our data revealed no significant difference in the distribution of KIR2DS4 variants in patients with B-ALL and controls (Tables 6 and 7). However, when we compared B-ALL patients and controls with AA genotypes in terms of KIR2DS4 variants, we found a significant difference (P=0.03). Further analysis revealed that KIR2DS4 del/del genotype was significantly more frequent among AA genotype carriers in the patient group than those in the control group (P=0.023, OR 4.26, 95% CI: 1.28-12.46).
| KIR2DS4 | B-ALL | Standard-risk | High-risk | Controls | B-ALL | Standard-risk vs. | High-risk | Standard-risk vs. | B-ALL with AA genotype | Controls with AA genotype | AA B-ALL vs. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| B-ALL | B-ALL | vs. Controls | Controls | vs. | High-risk | AA controls | |||||
| (n=116) | (n=73) | (n=43) | (n=165) | Controls | (n=46) | (n=52) | |||||
| Alleles | N | P-value | N | P-value | |||||||
| % | % | ||||||||||
| 40 | 27 | 13 | 68 | 0.37 | 0.68 | 0.32 | 0.51 | 4 | 19 | ||
| fl | 17.20% | 18.50% | 15.10% | 20.60% | 4.30% | 18.30% | 0.005 | ||||
| 192 | 119 | 73 | 262 | 88 | 85 | ||||||
| del | 82.80% | 81.50% | 84.90% | 79.40% | 95.60% | 81.70% | |||||
| Genotypes | N | P-value | N | P-value | |||||||
| % | % | ||||||||||
| 8 | 4 | 4 | 21 | 0.2 | 0.07 | 0.61 | 0.15 | 0 | 4 | 0.03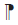 |
|
| fl/fl | 6.90% | 5.50% | 9.30% | 12.70% | 0% | 7.70% | |||||
| 84 | 50 | 34 | 118 | 42 | 37 | ||||||
| del/del | 72.40% | 68.50% | 79.10% | 71.50% | 91.30% | 71.20% | |||||
| 24 | 19 | 5 | 26 | 4 | 11 | ||||||
| fl/del | 20.70% | 26.00% | 11.60% | 15.80% | 8.70% | 21.20% | |||||
Note: fl: Full-length variant, del: Deleted variant
Comparison of the distribution of KIR2DS4 variants in AA genotype carriers between patients with B-ALL and healthy controls based on a 2 × 2
contingency table: del/del versus other genotypes (P=0.023, OR 4.26, 95% CI: 1.28-12.46); fl/fl versus other genotypes (P=0.16); fl/del versus other.
Table 6: Distribution of KIR2DS4 variants in patients with B-ALL and healthy controls.
| Genotype ID | Genotype | Centromeric genotype | Telomeric genotype | Bx subset | A haplotype-associated KIRs | B haplotype-associated | Framework | KIR pseudogenes | Frequency | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KIRs | KIRs | |||||||||||||||||||||
| B-ALL patients | Controls | |||||||||||||||||||||
| 3DL1 | 2DL1 | 2DL3 | 2DS4 | 2DL2 | 2DL5 | 3DS1 | 2DS1 | 2DS2 | 2DS3 | 2DS5 | 2DL4 | 3DL2 | 3DL3 | 2DP1 | 3DP1 | |||||||
| 1 | AA | A/A | A/A | -- | 46 | 52 | ||||||||||||||||
| 2 | AB | A/A | A/B | CxT4 | 8 | 14 | ||||||||||||||||
| 3 | AB | A/B | A/B | CxT4 | 4 | 8 | ||||||||||||||||
| 4 | AB | A/B | A/A | CxTx | 10 | 19 | ||||||||||||||||
| 5 | AB | A/B | A/A | C4Tx | 20 | 18 | ||||||||||||||||
| 6 | AB | A/B | A/B | C4T4 | 4 | 8 | ||||||||||||||||
| 7 | AB | A/B | A/B | C4Tx | 4 | 6 | ||||||||||||||||
| 8 | AB | A/A | A/B | CxTx | 4 | 1 | ||||||||||||||||
| 9 | AB | A/B | A/B | CxTx | 2 | 3 | ||||||||||||||||
| 10 | AB | A/B | A/A | CxTx | 0 | 1 | ||||||||||||||||
| 11 | AB | A/B | A/B | C4Tx | 0 | 1 | ||||||||||||||||
| 14 | AB | A/B | A/B | C4Tx | 6 | 0 | ||||||||||||||||
| 27 | AB | A/A | A/B | CxTx | 0 | 4 | ||||||||||||||||
| 28 | AB | A/A | A/B | CxT4 | 0 | 1 | ||||||||||||||||
| 62 | AB | A/B | A/A | CxTx | 0 | 2 | ||||||||||||||||
| 68 | AB | A/B | B/B | CxT4 | `` | 0 | 1 | |||||||||||||||
| 69 | AB | A/A | B/B | CxT4 | 2 | 2 | ||||||||||||||||
| 70 | AB | A/B | B/B | C4T4 | 2 | 1 | ||||||||||||||||
| 71 | AB | B/B | A/A | C4Tx | 4 | 8 | ||||||||||||||||
| 73 | AB | B/B | A/B | C4T4 | 2 | 9 | ||||||||||||||||
| 86 | AB | A/B | B/B | CxT4 | 0 | 1 | ||||||||||||||||
| 90 | AB | B/B | A/B | C4Tx | 2 | 3 | ||||||||||||||||
| 94 | AB | B/B | A/B | C4Tx | 0 | 1 | ||||||||||||||||
| 154 | AB | A/A | B/B | CxT4 | 0 | 1 | ||||||||||||||||
| 159 | AB | A/B | B/B | C4Tx | 0 | 1 | ||||||||||||||||
| 202 | AB | A/A | A/B | CxTx | 0 | 2 | ||||||||||||||||
| 381 | AB | A/B | A/B | CxTx | 0 | 2 | ||||||||||||||||
| Total | 120 | 170 | ||||||||||||||||||||
Table 7: KIR gene profiles of patients with childhood B-ALL and healthy controls (Gray and white cells denote the presence and the absence of the related genes, respectively).
NK cells are important components of the innate immunity againstmm tumor or virally infected cells with decreased expression of HLA class I molecules. Leukemic cells have been reported to have downmm regulated expression of HLA class I molecules [24-27]. The reasonmm why NK cells are not capable of destroying these leukemic cellsmm and eradicating them is of question. One possibility is that NK cellmm failure to destroy leukemic cells with aberrant expression of HLA class I molecules may be due to the NK receptors present in these patients. KIRs are key NK receptorsmm most of which recognize HLA class I molecules as their ligandsmm [28]. Moreover, the polygenic and multi-allelic nature of KIRsmm results in a vast diversity of KIR genotypes [29]. Considering thesemm two facts, KIRs are very likely to be the NK receptors responsiblemm for the dysfunctionality of NK cells in the destruction of leukemicmm cells.
Several studies have been carried out in order to investigate whether there is a relationship between KIR genes and the susceptibility to hematological malignancies [30-32]. However, controversial results were obtained. We performed a case-control study to find out whether this relationship exists in patients with childhood B-ALL. Our results revealed no association between none of the activating and inhibitory KIR genes and susceptibility to childhood B-ALL. We also found no relationship between AA and Bx genotypes, as well as centromeric and telomeric genotypes and this disease. Moreover, the number of centromeric and telomeric gene-content motifs that contained B haplotype-specific genes was not found to be associated with B-ALL development.
Our data were consistent with those which were extracted from the analysis of 185 childhood B-ALL and 33 childhood T-ALL patients along with ethnically matched controls [7]. Similarly, Shahsavar et al. did not find any associations between KIR genes or genotypes and ALL in a study of 38 patients with ALL and 200 healthy controls with Iranian origin [32]. In a family-based association study including 67 families of ALL, AML and CML patients, Varbanova aimed to find genetic differences that predisposed the affected siblings to leukemia, considering that siblings were exposed to identical environmental factors [10]. None of KIR genes and genotypes was recognized as predisposing factors for leukemia, which is consistent with our data on childhood B-ALL.
Our results were also in line with the data were reported by Oevermann et al. who did not find any relationships between KIR genes, genotypes, and KIR B-content scores and susceptibility to ALL in 328 patients with European origin [33]. Conversely, they reported that all aKIR genes were associated with resistance to childhood B-ALL [6]. They studied the six known aKIR genes in Canadian patients of white origin. However, they failed to study iKIR genes in these patients. The conflict between the results from this study and ours may be due to the diversity of KIR gene repertoire among different populations. It would have been of interest if the frequencies of iKIR genes had been determined in this study as well; because of the linkage disequilibrium that strongly exists between some of the aKIR and iKIR genes. Investigating the frequency of iKIR genes would have also helped to learn how KIR genotypes were related to the disease.
KIR2DS3 was reported to be significantly less frequent in ALL patients compared with healthy controls in a study [5]. However, the sample size was small in this study (n=61).
Our results were in stark contrast who reported all six aKIR genes (2DS1, 2DS2, 2DS3, 2DS4, 2DS5 and 3DS1) to be significantly more frequent among patients with childhood ALL compared with controls [9].
When considering the number of both aKIRs and iKIRs, we observed that aKIRs>iKIRs, iKIRs>aKIRs, and aKIRs=iKIRs were distributed similarly in B-ALL patients and controls, which agrees with the results from the study [32].
Moreover, we compared AA genotype carriers in the two groups of patients and controls in terms of KIR2DS4 variants. Our results revealed that the frequency of del/del genotype was significantly higher among patients than controls. It implies that individuals with AA genotypes who have no aKIRs are at a higher risk of developing ALL than those who carry at least one aKIR. This may be due to the lack of enough activating signals for NK cells to become activated and carry out effector functions, in AA genotype carriers homozygous for KIR2DS4 del variant.
Our results are consistent. Who found a higher frequency of the KIR2DS4 del allele in patients with CML compared with healthy controls [31]. Moreover, KIR2DS4 del allele was found to be associated with increased risk of Hodgkin’s lymphoma in a study [30,34,35].
However, in a previous study it was concluded that neither KIR2DS4 del variant nor KIR2DS4 del/del genotype were associated with increased susceptibility to childhood B-ALL. In thier study, the frequency of different KIR2DS4 genotypes including del/del, fl/ fl, and fl/del as well as 2DS4-/-were compared between patients with childhood B-ALL and healthy controls and no significant difference was obtained.
Since KIR molecules interact with MHC class I molecules as their ligands to regulate the function and development of NK cells, it would have been beneficial if MHC class I typing had been carried out in our study as well. Moreover, the product of different KIR alleles may differ in the affinity for their cognate MHC I ligands, the expression level at the cell surface and their signaling strength. Hence the lack of KIR gene polymorphism investigation is another potential limitation of our study. In addition to KIRs, other NK cell receptors take part in NK cell activation and effector function and need to be taken into account when studying NK cell activity. Finally, it would have been of great benefit if we had conducted our study on B-ALL patients with a larger sample size.
This work was done in partial fulfillment of the requirements for the MSc degree in Immunology awarded to Sepide Namdari and was financially supported by a grant from Shiraz University of Medical Sciences (grant number: 19859). We would like to thank the staff of Amir Oncology Hospital and Imam Reza Clinic for their cooperation.
Citation: Namdari S, Bordbar MR, Zekavat OR, Farjadian S (2021) Increased Frequency of KIR2DS4 del/del among AA Genotype Carriers with B-Cell Acute Lymphoblastic Leukemia in Southwestern Iranian Population. Immunogenet Open Access. 6:134.
Received: 11-Jan-2021 Accepted: 25-Jan-2021 Published: 02-Feb-2021 , DOI: 10.35248/igoa.21.6.134
Copyright: © 2021 Namdari S, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.