Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2014) Volume 3, Issue 1
Ginsenosides, which belong to group of triterpenoid saponins, are considered to be main constituents responsible for the biological effects of ginseng drug. Effect of organic and inorganic elicitors and other additives on the ginsenoside biosynthesis and biomass production by the in vitro cultivated adventitious root cultures of Korean ginseng was investigated in present study. Elicitation potential of natural mixtures (coconut water, pineapple extract, casein hydrolyzate, yeast extract, malt extract), organic elicitors (putrescine, spermidine, spermine, jasmonic acid), inorganic ions (calcium) and their complexes with organic acids (titanium ascorbate, titanium citrate) was tested. Changes in concentration of 9 main ginsenosides (Re, Rg1 , Rf, Rg2 , Rb1 , Rb2 , Rc, Rg3 and Rd) were monitored using HPLC-PDA and LC/MS/MS techniques. Jasmonic acid was the most effective elicitor of ginsenoside production (about 3.5 mg/g of DW in comparison with 1.4 mg/g of DW in control); however, its application (similar to spermine) was followed with significant reduction of the biomass growth. Among tested additives, casein hydrolyzate showed the highest increase of the ginsenoside content (2.2 mg/g DW) together with no effect on the growth of the culture. Both tested titanium complexes showed slight inhibition of the saponin accumulation in comparison with untreated control.
Keywords: Panax ginseng; Adventitious roots; Saponins; Elicitation
Korean ginseng belongs among the most popular adaptogens in the world, which is traditionally used for longevity promotion and improvement of tolerance to physical, chemical and biological stress [1]. Ginseng products are being distributed in 35 countries around the world and a total world ginseng production is roughly estimated to be 80,000 tons [2]. A number of various pharmacological effects of the ginseng drug include among others also anti-tumour activity, anti-diabetic effects, enhanced liver function, adjusted blood pressure, improved climacteric disorder and sexual functions [3]. The most of the medicinal properties of the ginseng is ascribed to ginsenosides, which belong to triterpenoid dammarane saponins [4]. The saponin content is one of the most important parameter in evaluation of ginseng quality [5,6].
Elicitation is one of the techniques used various chemical and physical factors (elicitors) as a trigger for increasing of production and accumulation of secondary metabolites [7]. Ones of the most widely used types of elicitors are signal molecules produced by plant under stress condition and pathogen attack. Jasmonic acid (JA) and salicylic acid derivatives belong to this group of signaling compounds, which are involved in many physiological processes such as developmental changes or defense responses. JA and especially its derivative methyl jasmonate (MeJA) have been found as very effective elicitors of saponin production in ginseng tissue cultures [8,9].
Besides the signaling molecules, there are other organic compounds as well as complex mixtures, which are able to induce secondary metabolism. Promotion of secondary metabolism, including saponin biosynthesis, in plant tissues by application of chitosan, yeast extract or casein hydrolyzate is well known [7]. Except the complex organic mixtures, there are also small organic molecules, which can activate phytoalexin production in plants [10]. It has been reported, that polyamines putrescine and spermidine induced production of isoflavonoids in vitro, which are thought to be typical defensive secondary metabolites [11]. Effect of polyamines on secondary metabolism could be caused by their influence on membrane transport due to the regulation of proton pumps (H+-ATPases) [12,13]. However, the suppression of the H+-ATPase activity by several organic and inorganic inhibitors leading to saponin overproduction in ginseng suspension cultures have been recently reported [14,15]. Except of organic elicitors, there are a number of inorganic stimuli such as heavy metal ions, used for the enhancement of secondary metabolism [15,16]. Nevertheless, most of the metallic ions with high elicitation potential used for that purposes (e.g. vanadium, cadmium ions) [15,17] are toxic also for animals and human beings, which is limiting for their biotechnological applications. Titanium, which is not considered to be toxic for vertebrates, has biological effects on plant growth and metabolism [18]. This element is not essential for plants, but it can be beneficial or toxic in dependence on its concentration [19]. Thus, the study of the titanium effect on saponin production in ginseng root cultures was involved in this work.
The aim of this study was to compare effect of organic and inorganic elicitors or other nutrient additives on saponin production and biomass growth in non-transformed adventitious roots of Panax ginseng.
Root cultures development and cultivation
Root cultures were developed according to modified method published previously [20]. Briefly: the plantlets of Panax ginseng C. A. Meyer were germinated from extirpated embryos on MS medium supplemented with 4.5 mM 2,4-D and 0.046 mM kinetin (both Sigma – Aldrich, Prague, Czech Rep.). The plantlets were rooted on MS medium with 1/3 concentration of inorganic nutrients. Adventitious roots isolated from the plantlets were cultivated on liquid SH medium with addition of 5mg/l IBA (Sigma – Aldrich, Prague, Czech Rep.) in 250 mL Erlenmeyer flasks in climabox with horizontal shaking in the dark at 24°C and subcultivated each 4 weeks.
Application of elicitors and additives
The tested elicitors and additives were added in to the medium after three weeks of cultivation. The cultures were harvested on 28 day after elicitation. Jasmonic acid (purity ≥ 97.0%), putrescine (purity ≥ 98.5%), spermine (purity ≥ 97.0%), spermidine (purity ≥ 98.0%) and calcium chloride (CaCl2·2H2O) as well as casein hydrolyzate, malt extract, pineapple extract and yeast extract were purchased from Sigma – Aldrich (Prague, Czech Rep.). Coconut water was prepared from fresh fruits bought in supermarket in Prague (Czech Rep.). Titanium ascorbate and titanium citrate complex solutions were prepared according to previously described methods [19]. Stock solutions of all tested additives except jasmonic acid were prepared as water solutions. Jasmonic acid was firstly dissolved in DMSO (1% v/v) then deionized water was added to final concentration of 10 mg per mL. The stock solutions were filtered using sterile syringe filter (Fisherbrand MCE, 25mm, 0.22μm, Fisher Scientific, Inc., Waltham, MA, USA) and appropriate volume was added to sterilised medium. Before a sterilization, pH of the cultivation media for spermine and spermidine was adjusted to lower value (approx. 5) then it was done in media for other additives (pH value 5.7-5.8) in order to strong alkaline reaction of both polyamines was equilibrated.
Biomass growth assessment
The biomass growth was calculated according to slightly modified method described in Collin and Edwards [21]. It was expressed as growth value (GV) of fresh weight, which is the ratio of fresh weight at time of harvest (after 4 weeks of cultivation) to fresh weight of the culture in time of elicitor addition.
Saponin extraction
Harvested roots were lyophilized and grinded with mortar and pestle to fine powder. Powdered material (0.3 g) was extracted two times for 12 hours with methanol at room temperature (24°C) and then the extract was evaporated to dryness under vacuum. Dry extract was redissolved in ultrapure deionized water (5 mL) in ultrasonic bath for 3 min. Then the samples were applied on conditioned SPE columns (OASIS HLB, 200 mg, Waters, Milford, MA, USA) and washed with ultrapure water (10 mL) Columns were washed with ultrapure water (10 mL) and the saponins were eluted out with 5 mL of methanol. Samples were dissolved in mobile phase and analyzed using HPLCPDA and LC/MS/MS methods.
HPLC-PDA and LC/MS/MS analysis
Samples from experiments involved application of complex organic mixtures, polyamines and jasmonic acid were quantified on HPLC with PDA detection. The system consists from quarternal gradient pump (Q-Grad, Watrex, Prague, Czech Rep.), autosampler Midas and column thermostat Mistral (both Spark Holland, Emmen, The Netherlands) and PDA detector UV6000LP (Thermo Finnigan, San Jose, CA, USA). Peak identity was confirmed using LC/MS/MS by comparison of fragmentation spectra and retention times with commercial standards (Sigma-Aldrich, Czech Rep.). Analysis was performed using Kinetex C18 column (5 μm, 250 × 4.6 mm, Phenomenex, Torrance, CA, USA) at temperature 30°C. Compounds were separated by gradient elution with mobile phase consisting of water (A) and acetonitrile (B) both with formic acid (0.1% v/v). The gradient started at 15% B in A, then followed to 30% B in 30 min, 50% B in 45 min and 100% B in 48 min. Flow rate was 1 mL/min. Injected volume was settled to 5 μL.
Quantitative analysis of the samples from tests of effect of both titanium complexes was carried out with LC/MS/MS on mass spectrometer Q-Trap 4000 (AB Sciex, Framingham, MA, USA) equipped with UPLC chromatographic system Ultimate 5000 (Dionex, Sunnyvale, CA, USA) using slightly modified MRM method previously published by Sun and Chen [6]. Separation was carried out on Kinetex C18 column (2.6 um, 150 x 2.1 mm, Phenomenex, Torrance, CA, USA), mobile phases and separation conditions were the same as in HPLCPDA analysis with slight modification of gradient, injected volume was reduced to 2 μL and the flow rate of mobile phase was 0.2 mL/min. Both positive (ginsenosides Rg1, Rf, Rb1, Rb2, Rg3, Rc, Rd) and negative (Re, Rg2) mode was used for the detection of measured saponins.
Tests involving comparison of organic elicitors as well as tests of titanium application were each carried out as two independent experiments. Each treatment was prepared in triplicate as minimum. One-way ANOVA followed with post-hoc Tukey test at p ≤ 0.05 was used for the assessment of statistical significance of the differences. STATISTICA 7.0 (StatSoft, Inc., Tulsa, OK, USA) software was used for all statistical analyses.
Comparison of elicitation effects of organic and inorganic additives and mixtures on production of ginsenosides by adventitious root cultures of Korean ginseng showed significant differences among used elicitors. The highest accumulation of ginsenosides was measured in the cultures treated with jasmonic acid (JA), where the amount of total ginsenosides in dry biomass was more than two times higher (about 3.5 mg/g) in comparison with control (1.4 mg/g) (Figure 1). The differences among all tested concentrations were not statistically significant. Similar increase of saponin production (from 2 to 4-fold) was described in different ginseng root cultures elicited with JA derivative methyl jasmonate (MeJA) [7,22]. Jasmonic acid and its derivatives such as MeJA are well known regulators of plant response to herbivore and pathogen attack. These molecules are involved in the regulation of many biosynthetic pathways leading to production of secondary metabolites. The saponin biosynthesis in Panax ginseng is probably strongly influenced by the jasmonate-dependent regulations [23]. It was confirmed by the other works focused on effect and utilization of MeJA (JA derivative) as an elicitor of ginsenoside production in vitro [9]. Moreover, presence of JA influenced particular ginsenosides profile due to changes of ratio between two main ginsenoside groups in the tissue, which was described for MeJA as well [24,25]. The JA-type elicitors strongly affected production of panaxadiol group of ginsenosides, whereas concentration of panaxatriol-derived ginsenosides reminded almost unchanged. The ratio of the main representatives (Rg1 and Rb1) of both ginsenoside groups is shown in the Table 1. Although the amount of total ginsenosides in DW was approximately equal for all tested concentrations of JA (5, 10 and 50 mg/L), biomass production strongly decreased with growing JA concentration (Figure 2). Moreover, the fresh weight biomass of the cultures was reduced (negative GV) during the cultivation at the highest concentration of JA (50 mg/L) (Figure 2). The increment of the dry weight was inhibited as well, but the decrease was not so dramatic (data not shown). Suppressed growth of biomass is a typical effect of application of JA-pathway regulators [22,24]. Thus, the ginsenoside productivity of the JA elicitation is decreased by the low biomass growth of the cultures.
| Treatment | Rg1/Rb1 ratio | Treatment | Rg1/Rb1 ratio |
|---|---|---|---|
| coconut water (10 % v/v) | 1.51 ± 0.29 | Ti - ascorbate (0.1 ppm) | 1.03 ± 0.35 |
| pineaple extract (10 g/l) | 1.13 ± 0.32 | Ti - ascorbate (0.5 ppm) | 1.08 ± 0.19 |
| casein hydrolyzate (0.5 g/l) | 2.00 ± 0.50 | Ti - ascorbate (1 ppm) | 1.18 ± 0.33 |
| yeast extract (1.0 g/l) | 1.25 ± 0.10 | Ti - ascorbate (2 ppm) | 1.00 ± 0.24 |
| increased Ca (1.0 g/l) | 2.12 ± 0.24 | Ti - citrate (0.1 ppm) | 0.98 ± 0.38 |
| malt extract (1.0 g/l) | 1.53 ± 0.77 | Ti - citrate (0.5 ppm) | 0.99 ± 0,10 |
| putrescine (1 mM) | 2.88 ± 0.57 | Ti - citrate (1 ppm) | 1.11 ± 0.20 |
| spermidine (1 mM) | 2.13 ± 0.47 | Ti - citrate (2 ppm) | 0.92 ± 0.04 |
| spermine (1 mM) | 1.03 ± 0.27 | control | 1.16 ± 0.15 |
| jasmonic acid (5 mg/l) | 0.61 ± 0.30 | ||
| jasmonic acid (10 mg/l) | 0.45 ± 0.08 | ||
| jasmonic acid (50 mg/l) | 0.37 ± 0.09 |
Table 1: Ratio of Rg1 (most abundant panaxatriol) and Rb1 (dominant panaxadiol) in adventitious root cultures of Panax ginseng treated with organic and inorganic additives. The data were obtained 4 weeks after additive application. Ratio is expressed as mean of 3 values obtained as biological triplicates ± standard deviation.
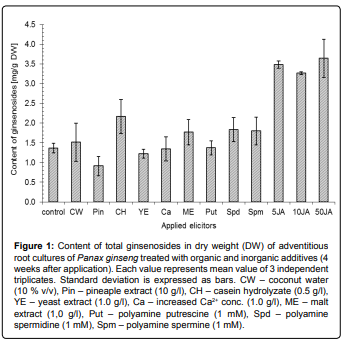
Figure 1: Content of total ginsenosides in dry weight (DW) of adventitious root cultures of Panax ginseng treated with organic and inorganic additives (4 weeks after application). Each value represents mean value of 3 independent triplicates. Standard deviation is expressed as bars. CW – coconut water (10 % v/v), Pin – pineaple extract (10 g/l), CH – casein hydrolyzate (0.5 g/l), YE – yeast extract (1.0 g/l), Ca – increased Ca2+ conc. (1.0 g/l), ME – malt extract (1,0 g/l), Put – polyamine putrescine (1 mM), Spd – polyamine spermidine (1 mM), Spm – polyamine spermine (1 mM).
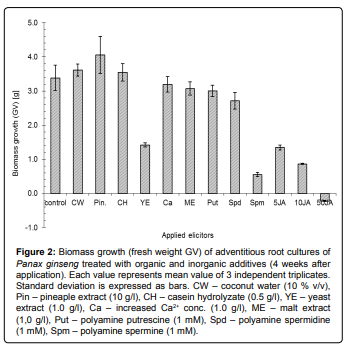
Figure 2: Biomass growth (fresh weight GV) of adventitious root cultures of Panax ginseng treated with organic and inorganic additives (4 weeks after application). Each value represents mean value of 3 independent triplicates. Standard deviation is expressed as bars. CW – coconut water (10 % v/v), Pin – pineaple extract (10 g/l), CH – casein hydrolyzate (0.5 g/l), YE – yeast extract (1.0 g/l), Ca – increased Ca2+ conc. (1.0 g/l), ME – malt extract (1,0 g/l), Put – polyamine putrescine (1 mM), Spd – polyamine spermidine (1 mM), Spm – polyamine spermine (1 mM).
Similar to JA, yeast extract and especially polyamine spermine also significantly reduced growth of the ginseng cultures, but only spermine slightly induced saponin production (Figure 2). Other tested polyamines spermidine and putrescine showed no effect on the growth rate. Although both polyamines are reported to stimulate production of several secondary metabolites (e.g. isoflavones) in vitro [11], there was only moderate increase of the ginsenoside content after spermidine treatment (Figure 1).
Casein hydrolyzate seemed to be the most effective additive among tested complex mixtures because of its ability to slightly stimulate saponin production (2.2 mg/g DW) together with no reduction of the biomass (Figure 1). The positive effect of casein hydrolyzate addition on secondary metabolism in vitro has been reported for other in vitro systems such as production of furanocoumarin psoralen in cultures of Psoralea corylifolia [26]. Other natural mixtures had no significant influence on either saponin accumulation or growth of the culture.
In addition, inorganic ions tested in our study have either no effect on saponin accumulation (Ca2+) or slightly suppressed their production (Ti complexes) (Figures 1 and 3). The biomass growth remain unchanged in control as well as treated cultures (Figure 4), although there are some published results described the influence of Ti to plant growth in field experiments [19]. The differences between saponin accumulation, biomass production and panaxadiol/panaxatriol ratio in both types of titanium complexes (Ti citrate and Ti ascorbate) at all tested concentrations were almost negligible. The absence of any physiological response of in vitro cultures may be caused by different availability of nutrients in vitro and in vivo [27]. Thus, in order to elucidate the role of titanium in ginseng secondary metabolism in vitro, the further experiments focused on nutrient availability should follow.
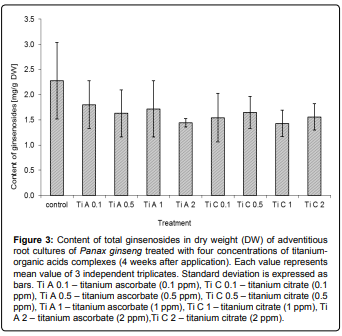
Figure 3: Content of total ginsenosides in dry weight (DW) of adventitious root cultures of Panax ginseng treated with four concentrations of titaniumorganic acids complexes (4 weeks after application). Each value represents mean value of 3 independent triplicates. Standard deviation is expressed as bars. Ti A 0.1 – titanium ascorbate (0.1 ppm), Ti C 0.1 – titanium citrate (0.1 ppm), Ti A 0.5 – titanium ascorbate (0.5 ppm), Ti C 0.5 – titanium citrate (0.5 ppm), Ti A 1 – titanium ascorbate (1 ppm), Ti C 1 – titanium citrate (1 ppm), Ti A 2 – titanium ascorbate (2 ppm),Ti C 2 – titanium citrate (2 ppm).
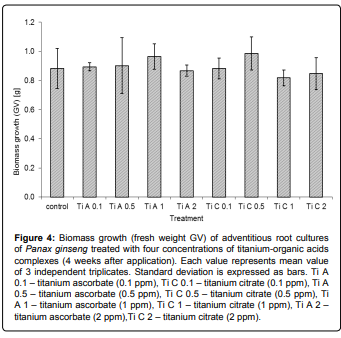
Figure 4: Biomass growth (fresh weight GV) of adventitious root cultures of Panax ginseng treated with four concentrations of titanium-organic acids complexes (4 weeks after application). Each value represents mean value of 3 independent triplicates. Standard deviation is expressed as bars. Ti A 0.1 – titanium ascorbate (0.1 ppm), Ti C 0.1 – titanium citrate (0.1 ppm), Ti A 0.5 – titanium ascorbate (0.5 ppm), Ti C 0.5 – titanium citrate (0.5 ppm), Ti A 1 – titanium ascorbate (1 ppm), Ti C 1 – titanium citrate (1 ppm), Ti A 2 – titanium ascorbate (2 ppm),Ti C 2 – titanium citrate (2 ppm).
The comparison of the various elicitors including complex organic mixtures, polyamines, JA and inorganic ions and their complexes was carried out in this study. Application of titanium complexes with organic acids on ginseng root cultures had slight inhibitory effect on saponin production and no influence on growth parameters. The highest content of ginsenosides in the dry weight was accumulated by the cultures treated with JA. However, the efficiency of the elicitation was diminished due to reduction of biomass. Among the other elicitors, only casein hydrolyzate showed moderate positive effect on ginsenoside production without any suppression of biomass growth.
This work was supported by COST LD13013 project granted by Ministry of Education, Youth, and Sports of the Czech Republic.