Immunome Research
Open Access
ISSN: 1745-7580
ISSN: 1745-7580
Research Article - (2015) Volume 11, Issue 1
Mutalysin-II, from Lachesis muta muta snake venom, is an endopeptidase with hemorrhagic activity. To identify a conformational epitope we used Blue Star Sting, the latest version of the web based Sting Millenium Suite, as alternative computational analysis tool to select and design peptides. Pre-selected cut-off values of the accessibility and hydrophilicity parameters were used to select amino acid residues as potential conformational epitopes. A peptide (P117-Y116-C115-Q194-C195-L197-N198-K199-P200-Y5-L48) was manually drawn on Swiss-PDB-Viewer package and synthesized by Fmoc-synthesis. Immunization of rabbits with this peptide induced antibodies that recognized Mutalysin-II and protected against the hemorrhagic factors present in Lachesis venom. The Sting Millennium Suite was able to predict conformational epitopes in this class of proteins. Three amino acids (K199, Y5 and L48) were identified as essential in the interaction between the peptide and the neutralizing antibodies.
<Keywords: Lachesis muta muta venom; Antivenom; Sting millennium; Synthetic peptides; Epitopes; Conformational epitope prediction
Snakebites are considered a neglected tropical condition in the World and an important public health problem in Brazil [1]. The pit viper envenomation is generally characterized by severe hemorrhage, blood incoagulability and renal insufficiency. In Bothrops sp. and Lachesis sp. Snake Venoms Metalloproteinases (SVMPs) are the most important contributors to local and systemic hemorrhage observed in human snakebite victims [2]. The venom of the pit viper L. muta muta contains a proteinase, called mutalysin-II (Mut-II) a P-I snake venom metalloproteinase (P-I SVMP) and belongs to a major class of metalloproteinases known as the metzincins [3]. Mut-II is a 22.5 kDa single chained protein with broad substrate specificity and traces of hemorrhagic effects [3,4]. Like other SVMPs, mutalysins proteolytically degrade fibrinogen and other extracellular matrix components such as laminin, fibronectin, and type IV collagen or gelatin [4].
Due to the medical importance of the envenomation caused by the L. muta muta and the role played by P-I SVMPs in these accidents, polyclonal and monoclonal antibodies against these zinc-endopeptidases may be considered as highly useful tools to investigate the structural determinants of toxicity and antigenicity/ immunogenicity of these molecules. Furthermore, these antibodies could have great potential for the preparation of more efficacious antivenoms for passive immunotherapy or even for vaccination. Effectively, the monoclonal antibody LmmAbB2D4 [5] and rabbit polyclonal antibodies [6] against Mut-II efficiently neutralize the hemorrhagic effects of whole L. muta muta venom and certain Bothrops venoms. Prediction and mapping of neutralizing epitopes on these toxic proteins are vital steps to create epitope-based antivenoms design (i.e. antivenoms produced using synthetic peptides corresponding to identified epitopes as immunogens). We have showed that synthetic peptides corresponding to linear and conformational epitopes previously mapped using laboratory methods, are capable of inducing neutralizing antibodies against L. muta muta crude venom [6,7]. Although laboratory techniques, such as Phage display [8] and Spot method [9], are effective in identifying epitopes, they are expensive and time consuming. Therefore, computational analysis can provide a rapid method to identify functional epitopes using data routinely collected from toxins. STING Millennium Suite (SMS) [10] is web-based, publicly available software developed to aid researches in their quest for translating information about the structures of macromolecules into knowledge. Using SMS it is possible to analyze the sequence/structure relationship, quality of the structure, nature and volume of intra and inter chain atomic contact and the relative conservation of amino acids at the specific sequence position based on multiple sequence alignment to mention only a few [11]. In the present investigation, we used STING Millennium Suite (SMS) as an alternative computational analysis tool to design and select a peptide with characteristics of conformational epitopes on Mut-II.
Molecular modeller and Identification of conformational epitopes
We used the website BSS platform to predict/select those amino acid residues that could constitute a potential conformational epitope. The BSS itself is not a program for predicting epitopes; it is a web based software for interactive analysis of the sequence/structure/function of proteins and their complexes with DNA and ligands, allowing the user to interactively analyze molecular structures, cross-referencing visualized information with correlated attributers stored in the BSS database [11]. BSS has a module, (the JPD) [28] that communicate a large set of the physicochemical properties of proteins at a residueby- residue level through a unique graphic interface. JPD currently connects to a total of 1540-numerical/textual descriptors for any given protein structure deposited in the PDB in a residue by residue manner ((8.89 × 1010 records -105000 structures with 2.2 chains in average, with 250 amino acids in average per chain) [10]. We adapted the JPD to select the potential amino acid residues with epitopic function. For this, we used the JPD to select amino acid residues that are both hydrophilic and also have high accessibility in Mut-II. Twenty-five residues were selected by the combination of these parameters, they are: K19, N21, N25, N36, N59, Q60, N64, N70, K74, N86, S89, N104, Y116, P129, K149, N153, S158, P172, Q185, M186, K190, K192, Q194, L197, K199 and can be visualized in the 3D structure of Mut-II (Figure 1A and Table 1). The conformational nature of the epitope residue assemblage is evident since the selected residues are not found in a linear sequence in the primary structure. The residues were grouped into two clusters, according to the spatial distance in the 3D structure. The group containing the highest number of selected residues had a peptide designed. The 3D peptide structure was manually designed in SPDBV tool combining the selected amino acid residues with their spatial neighbours to obtain a peptide with physical-chemical characteristic similar to that of the potential epitopic region of the Mut-II. Thus, the cluster possessing the residues: Y116, P117, Q194,L197, K199 (Green at Figure 1A), along with its spatial neighbours, Y5, L48, C115, C195, N198, P200 (Figure 1A), were combined, to obtain the peptide P117-Y116-C115-Q194-C195-L197-N198-K199- P200-Y5-L48 (Figure 1B).

Figure 1: Visualization of the residues selected by STING as epitopes in the Mut-II and a peptide proposed in a 3D structure. In (A): visualization of amino acid residues on the molecular surface of the Mut-II. The amino acid residues selected by STING are in green and the neighbor amino acid residues used to propose the peptide are in yellow. (B): Location of the peptide in the predicted 3D structure of Mut-II. In blue, the C-terminal region; in orange, the amino acids that compose the peptide; and in magenta, the essential amino acids identified by AlaScan.
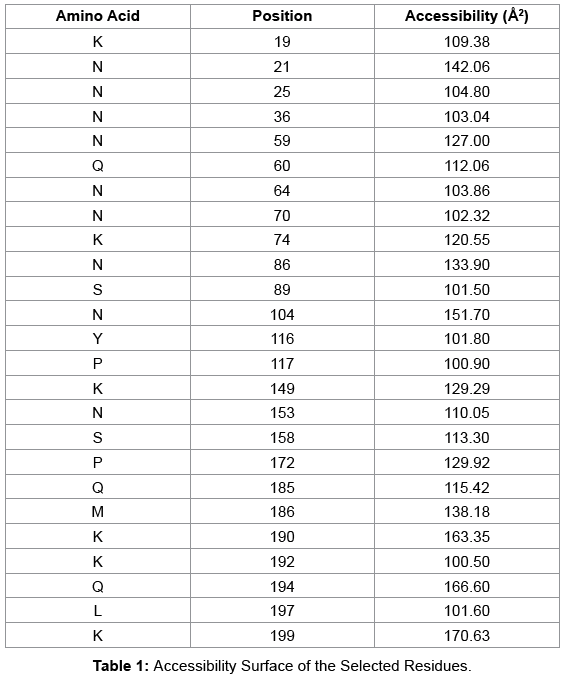
Table 1: Accessibility Surface of the Selected Residues.
Neutralizing ability of anti-peptide antibodies
In order to confirm it as a conformational epitope, the peptide sequence PYCQCLNKPYL was synthesized and its immunogenicity tested. The peptide was synthesized using Fmoc Chemistry Technique [7,29]. The N-terminal portion was acetylated and the C-terminal portion was amidated. Subsequently, it was purified by HPLC in C18 columns and the major peak was validated by mass spectrometry, having an expected monoisotopic mass of 1382.67Da (data not shown).
After, the peptide was incorporated into liposomes using the dehydration-rehydration method [30]. Then, three New Zealand white rabbits were immunized with liposome containing the peptide. As a control, two rabbits were also immunized with empty liposomes, containing no synthetic peptides. After the first immunization, eight boosters were performed as described before. Seven days after the last immunization the rabbits were bled and their sera were obtained by centrifugation at 3000 rpm for subsequent analysis in ELISA and Western Blotting.
Antibodies raised against the designed peptide were able to recognize both purified mut-II and L. muta muta crude venom in ELISA (Figure 2A) and in Western Blotting (Figure 3A) in a similar manner compared to anti-mutalysin-II serum (Figure 2B and Figure 3B). Fifteen days after the last immunization, we conducted a hemorrhagic assay to verify whether this peptide would also be capable of inducing the production of neutralizing antibodies against the hemorrhagic toxins in L. muta muta venom. Thus, 20μg/Kg of L. muta muta venom were injected intradermally into immunized rabbits and after 24 hours the rabbits were euthanized in a CO2 chamber and their skins were removed in order to analyze the hemorrhagic lesions caused by the venom. Figure 4 shows the visceral side of the skins removed from the dorsum of the rabbits. The two rabbit controls, which received only the liposome without peptides during their immunizations, showed the greater hemorrhagic injury. In contrast, the three rabbits immunized with the peptide showed no signs of bleeding, thus indicating that immunizations were efficient for the production of neutralizing antibodies against bleeding factors in the venom of L. muta muta.

Figure 2: Immunoreactivity of anti-peptide and anti-mut-II against the designed peptide, mutalysin-II and L. muta muta venom by ELISA. (A) ELISA showing the reaction of anti-peptide antibodies against peptide, mut-II or L. muta muta whole venom. (B) Anti-mut-II was also tested against the same antigens, showing a stronger but similar pattern of bands recognition compared to anti-peptide serum. In both cases microtiter plates were coated with 10 μg/mL solution of each antigen. The absorbance of the samples was determined at 492 nm.

Figure 3: Reactivity of anti-peptide and anti-mut-II sera with mutalysin-II or Lachesis muta muta whole venom analyzed by western blotting. 20 μg of Lachesis muta muta crude venom (CV) and mutalysin-II (Mut-II) were transferred to a nitrocellulose membrane and incubated with rabbit polyclonal anti-peptide (A), anti-mut-II (B) or non-immune (C) sera (all sera were diluted 1:25). Anti-peptide and mut-II recognized bands around 23 kD.

Figure 4: The in vivo neutralization assays of the hemorrhagic activity of L. muta muta venom. The immunized and control rabbits were challenged with L. muta muta venom 15 days after the last immunization by intradermal injection of an amount equivalent to 1 MND/kg.
To verify which amino acids residues in the peptide were important in the interaction with rabbit anti-PYCQCLNKPYL polyclonal antibodies, a series of alanine analogs of the peptide was prepared by Spot synthesis and assayed for reactivity against anti-PYCQCLNKPYL antibodies. In each spotted peptide, an amino acid at a time was substituted by alanine. The results (Figure 5) indicate that in the C-terminal part of the peptide some residues could not be changed into alanine without decreasing their antibody binding capacity: this is the case for residues Lys8, Tyr10 and Leu11. These residues are, thus, key contributors in this antigenic region of the predicted conformational epitope of mutalisin-II. Residues in the N-terminal part of the peptide play minor roles in antibody recognition.

Figure 5: Reactivity of ALA-Scan peptides. Anti-peptide rabbit sera reactivity with cellulose-bound peptides was detected with an alkalinephosphatase coupled anti-rabbit antibody. Spot intensities were measured with the Scion Image software. Reactivity below 75000 was considered negative.
The peptide sequence PYCQCLNKPYL was designed in silico as a conformational epitope previously using BSS analysis, which selected only hydrophobic and solvent accessible amino acid residues above determined threshold. This peptide entrapped in liposomes was used to determine its immunogenicity in rabbits. Liposomes are biodegradable particles considered an excellent form to deliver biologically active substances [35]. The gradual release is essential to increase the lifetime of the peptide during its presentation to the immune system, which can improve the production of specific antibodies [36]. Anti-peptide sera produced in liposomes recognized mut-II and L. muta muta venom in ELISA and Western Blotting.
To measure the in vivo neutralization capacity of anti-peptide antibodies against the hemorrhagic activity of L. muta muta venom, the immunized rabbits were injected with L. muta muta crude venom [12,30]. Pre-existing rabbit anti-peptide antibodies neutralize efficiently the hemorrhagic activity of the crude L. muta muta venom [6]. Thus, it is reasonable to assume that anti-peptide serum binds to the SVMPs of the crude venom. Tanjoni et al. 2003, reported the binding of a monoclonal antibody to the disintegrin-like domain of jararhagin, a PIII-class metalloproteinase, neutralizing completely the hemorrhagic activity of the toxin. This domain is located at the C-terminal portion of the toxin. They supposed that the mechanism of this neutralization is due the proximity of this domain to the catalytic site of the protein, which probably prevented the interaction between enzyme and substrate, thus causing neutralization [37]. Our designed peptide possesses amino acid residues located near the C-terminal region of Mut-II, as shown in Figure 1B. Despite the differences between Mut-II and jararhagin, antibodies anti-peptide of L. muta venom might present allosteric inhibitor mechanism. The binding of the antibody in a region spatially near the C-terminal portion of the Mut-II could cause a steric shielding of the catalytic site, blocking its binding to the substrate and thereby inhibiting the hemorrhagic effect of the toxin. The absence of K199, L48 and Y5 in analogous peptides decreases their reactivity with antibodies (identified by alanine-scanning), therefore suggesting that these amino acids are essential for the interaction with antibodies.
In this way, we suppose that the in silico designed peptide represents a discontinuous or conformational B-cell epitope capable to induce neutralizing antibodies against the hemorrhagic factor of the L. muta muta venom. Parameters such as the hydrophilicity, static accessibility and mobility of short segments of polypeptide chains have been correlated with the location of continuous epitopes in proteins [38]. B cell epitopes contain both continuous (~10%) and discontinuous (~90%) epitopes, therefore the majority of B-cell epitopes are discontinuous. Currently, only a few methods and prediction tools for discontinuous B-cell epitopes are available, and their performance is generally very poor, because they lack experimental verification [39]. Our results suggest that the accessibility and hydrophilicity are also imortant parameters and may be used as facilitators to predict a conformational epitope in toxic proteins. Different bioinformatics programs use specific groups of parameters, varying from one another, including in hydrophobicity, antigenicity, flexibility, accessibility, amino acids considered as special in the interactions with antibodies, segmental mobility, among others, resulting in the prediction of different epitopes, depending on the accuracy of these programs (nowadays, the programs for the prediction of epitopes have a range of accuracy varying from 35-75%) [40]. In conclusion, our results show the generation and characterization of a neutralizing conformational epitope against Mut-II from L. muta muta venom and that the accessibility and hydrophilicity are important parameters to predict conformational epitopes in this class of proteins.
The peptide sequence PYCQCLNKPYL was designed in silico as a conformational epitope previously using BSS analysis, which selected only hydrophobic and solvent accessible amino acid residues above determined threshold. This peptide entrapped in liposomes was used to determine its immunogenicity in rabbits. Liposomes are biodegradable particles considered an excellent form to deliver biologically active substances [35]. The gradual release is essential to increase the lifetime of the peptide during its presentation to the immune system, which can improve the production of specific antibodies [36]. Anti-peptide sera produced in liposomes recognized mut-II and L. muta muta venom in ELISA and Western Blotting.
To measure the in vivo neutralization capacity of anti-peptide antibodies against the hemorrhagic activity of L. muta muta venom, the immunized rabbits were injected with L. muta muta crude venom [12,30]. Pre-existing rabbit anti-peptide antibodies neutralize efficiently the hemorrhagic activity of the crude L. muta muta venom [6]. Thus, it is reasonable to assume that anti-peptide serum binds to the SVMPs of the crude venom. Tanjoni et al. 2003, reported the binding of a monoclonal antibody to the disintegrin-like domain of jararhagin, a PIII-class metalloproteinase, neutralizing completely the hemorrhagic activity of the toxin. This domain is located at the C-terminal portion of the toxin. They supposed that the mechanism of this neutralization is due the proximity of this domain to the catalytic site of the protein, which probably prevented the interaction between enzyme and substrate, thus causing neutralization [37]. Our designed peptide possesses amino acid residues located near the C-terminal region of Mut-II, as shown in Figure 1B. Despite the differences between Mut-II and jararhagin, antibodies anti-peptide of L. muta venom might present allosteric inhibitor mechanism. The binding of the antibody in a region spatially near the C-terminal portion of the Mut-II could cause a steric shielding of the catalytic site, blocking its binding to the substrate and thereby inhibiting the hemorrhagic effect of the toxin. The absence of K199, L48 and Y5 in analogous peptides decreases their reactivity with antibodies (identified by alanine-scanning), therefore suggesting that these amino acids are essential for the interaction with antibodies.
In this way, we suppose that the in silico designed peptide represents a discontinuous or conformational B-cell epitope capable to induce neutralizing antibodies against the hemorrhagic factor of the L. muta muta venom. Parameters such as the hydrophilicity, static accessibility and mobility of short segments of polypeptide chains have been correlated with the location of continuous epitopes in proteins [38]. B cell epitopes contain both continuous (~10%) and discontinuous (~90%) epitopes, therefore the majority of B-cell epitopes are discontinuous. Currently, only a few methods and prediction tools for discontinuous B-cell epitopes are available, and their performance is generally very poor, because they lack experimental verification [39]. Our results suggest that the accessibility and hydrophilicity are also imortant parameters and may be used as facilitators to predict a conformational epitope in toxic proteins. Different bioinformatics programs use specific groups of parameters, varying from one another, including in hydrophobicity, antigenicity, flexibility, accessibility, amino acids considered as special in the interactions with antibodies, segmental mobility, among others, resulting in the prediction of different epitopes, depending on the accuracy of these programs (nowadays, the programs for the prediction of epitopes have a range of accuracy varying from 35-75%) [40]. In conclusion, our results show the generation and characterization of a neutralizing conformational epitope against Mut-II from L. muta muta venom and that the accessibility and hydrophilicity are important parameters to predict conformational epitopes in this class of proteins.
This research was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil - CAPES (Toxinologia No 23038000825/2011-63), Fundação de Amparo a Pesquisa do Estado de Minas Gerais, Brazil (FAPEMIG) and by funds of the INCTTOX Program of Conselho Nacional de Desenvolvimento Científico e Tecnológico, Brazil (CNPq). We would like to thank the Núcleo de Estudo de Estrutura e Função de Biomoléculas [Departamento de Bioquímica e Imunologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais (UFMG), Belo Horizonte, Minas Gerais, Brasil] for technical support for mass spectrometry.