Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2013) Volume 4, Issue 1
The effect of live and dry diets on the growth and survival of cultured chub (Leuciscus cephalus) larvae was measured from 5 days post-hatch and over a 24 days period. At its conclusion, larvae fed artificial dry diets were significantly shorter, weighed less and suffered higher mortality rates than those fed Artemia and a mixed diet of Artemia spp. and a commercial feed.
Keywords: Leuciscus cephalus; Artemia; larviculture; chub.
Fish hatcheries produce and stock-out substantial numbers of fishes annually in order to supplement wild fish populations, conserve threatened species, compensate for disturbance and support recreational fisheries [1]. Supplementing riverine populations with cultured rheophylic fishes is a common management practice [2,3], including the stocking of chub Leuciscus cephalus, a cyprinid inhabiting river systems across Europe [4]. Whilst large numbers of these fish are supplied through hatcheries for restocking both lotic and lentic sports fisheries, knowledge on their optimum culture practises is limited, with minimal data available on their larviculture [5,6].
Production of high quality juvenile fishes is arguably the most difficult part of the aquaculture production process [7] due to the, often, poor understanding of larval nutritional requirements [8]. Successful feeding is difficult as developing larvae are usually very small and fragile with an undeveloped digestive system [9]. Finfish larvae in hatchery environments have tended to be fed live food [10], with high dependence on Artemia spp. as a starter diet [11]. Artemia possess a strong amino acid composition that make them suitable for larval finfish culture [12] owing to the presence of low molecular weight peptides and free amino acids that, together with their autolytic capacity and high solubility, result in a relatively easy digestion of the protein in Artemia by fish larvae [13]. However, Artemia production requires considerable resources to consistently produce the required volume [14] and this can prove restrictive [10]. Thus, there have been attempts to shift from Artemia to dry diets to improve this aspect of the fish culture process [11]. Consequently, the aim of this study was to determine whether commercially available dry larval rearing diets were capable of producing superior growth and survival rates in L. cephalus compared to feeding Artemia.
The aim of the study, therefore, was to determine whether commercially available dry larval rearing diets can attain superior levels of growth and survival compared to Artemia for fish used in restocking programmes. The study investigated the use of starter feeds as well as a combination of live and dry feeds in a series of four rearing experiments. It is envisaged that the results would aid the standardisation of methods for rearing of chub (L. cephalus) larvae, as very little is currently known about the culture of this species. The results could also be used to standardise the rearing of other species.
A total of 8000 chub larvae were produced and maintained from broodfish sourced from the River Trent, Calverton Fish Farm, Nottingham, UK. Fish were used across four treatments, where each treatment was conducted in a 2.7 × 0.5 × 0.15 m feeding trough. The troughs were separated into four watertight sections of 0.68 m length, with each section having its own screened water in- and outlet. The flow rate through each of the tanks was set at 3.8 L-1 min. All treatments were randomly allocated to the troughs. Prior to larvae introduction, water in the troughs was heated to 20°C, with this maintained via a matrix heater in the header tank. Water temperature, pH and oxygen levels were measured daily and did not differ significantly throughout the trial. The filtration system consisted of a sedimentation tank and trickle filter and water was passed through a UV filter before returning to the troughs. At their release into the troughs, the larvae were 5 days post-hatch and had been fed Rotifers (Brachionus calyciflorus) for 3 days. The fish were split equally among the 16 troughs (n 5 500).
The feeding trail lasted 24 days, after which fish were weaned onto larger grower pellet. Fish were fed one of four diets that comprised of unenriched Artemia and two nutritionally complete commercially available slow sinking cyprinid starter diets (feeds A and B) used to mimic the size of newly hatched Artemia nauplii. For the first 10 days larvae were fed a 200 μm crumb and thereafter crumb of between 3 and 500 μm was used. The composition of the four diets was: (i) Artemia only, (ii) commercial feed A, (iii) Artemia and commercial feed A (1:1) and (iv) commercial feed B. The average composition of Artemia was 52.2% protein, 18.9% lipid and 9.7% ash [15] (% dry matter); whereas diet A was 45.0%, 10.0% and 8.5% respectively and diet B was 60.0%, 14.5% and 11.5% respectively (as fed) (figures supplied by the manufacturers). Food rations of various species of cyprinid larvae vary [16–18] whilst no information exists for the larvae of L. cephalus. Using this information larvae were fed 50% body weight four times a day at for the first 10 days, afterwhich this was reduced to 25% body weight [16]. Commercial fed larvae were provided two rations of dry food and two rations of Artemia to reduce possible consumption of only one food type. On a daily basis, the troughs were siphoned and outlet screens brushed to remove mortalities, uneaten food and faecal matter. The siphoned material was examined using a Meiji RZBD/200 microscope to determine the presence of uneaten Artemia and pellet. The uneaten food was separated, dried for 24 h and weighed.
To determine the effects of diet on fish length and weight, samples were taken every 4th day when 20 randomly selected fish per replicate tank were removed with fine meshed net and over-anaesthetised using Benzocaine. These fish were removed prior to the first feed of that day. Fish lengths were determined from image analysis, obtained using a high power microscope (Meiji RZBD/200) and MA751 photo tube attachment with fibre optic light (Meiji FL-150 High Intensity Illuminator), with image capture by a Lumenera Infinity 2 camera attachment. Total length was then determined using Infinity Analyze 4.5 software. Each fish was also weighed (nearest 0.1 mg). Excess water was blotted from each fish prior to weighing. Prior to the commencement of the trial, a sub-sample of 20 fish from across the treatments were also measured and weighed. The effect of diet on survival was assessed at the conclusion of the 24 days trial through the proportion of remaining live larvae. In statistical tests, parametric tests were only completed after testing for normality and only used when all test assumptions were met. Data were analysed using Minitab 16 Statistical software. A One-way ANOVA was carried out to compare diets against weight, length and mortality. Significance was accepted when P< 0.05.
Daily examination of waste matter revealed no significant differences in the quantity of uneaten Artemia and commercial feed across the treatments for the duration of the trial (P< 0.05). Length data (Figure 1) showed that at the conclusion of the trial, larvae fed on Artemia 1 commercial feed A were significantly longer than larvae fed on commercial feeds A and B (ANOVA, F 5 651.24; d.f. 5 3; P<0.001).
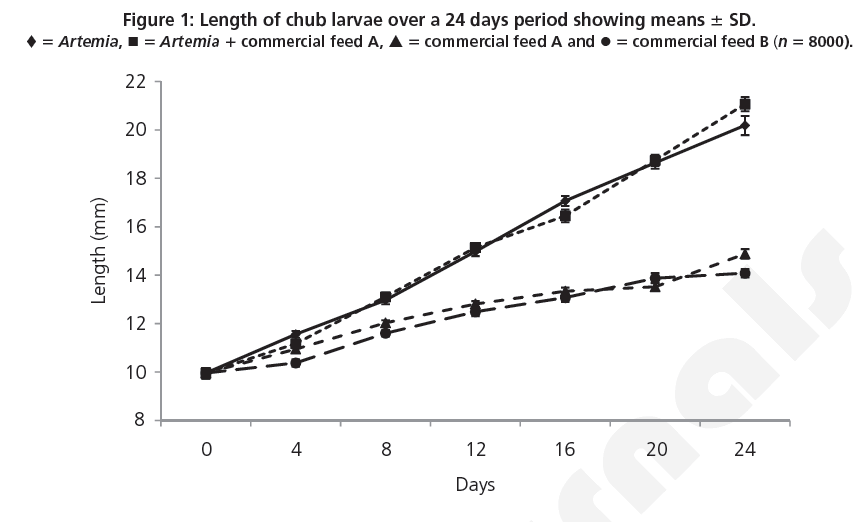
Figure 1: Length of chub larvae over a 24 days period showing means 6 SD. ♦ 5 Artemia, ◾ 5 Artemia 1 commercial feed A, ▴ 5 commercial feed A and • 5 commercial feed B (n 5 8000).
Larvae at day 24 that were fed on Artemia and Artemia 1 commercial diet A were found to be significantly heavier than larvae fed on commercial feeds A and B (ANOVA, F 5 86.55; d.f. 5 3; P<0.001) (Figure 2).
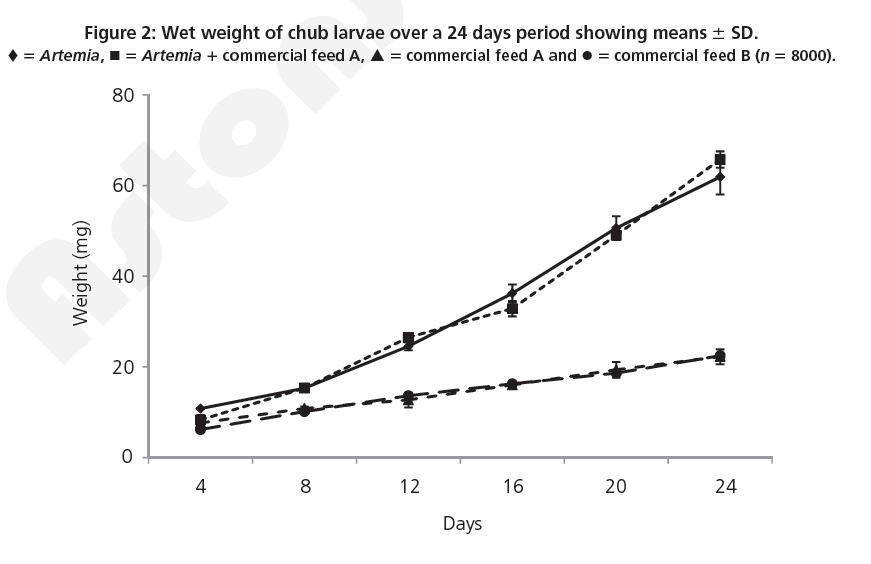
Figure 2: Wet weight of chub larvae over a 24 days period showing means 6 SD. ♦ 5 Artemia, ◾ 5 Artemia 1 commercial feed A, ▴ 5 commercial feed A and • 5 commercial feed B (n 5 8000).
The mortality data (Figure 3) for days 1 to 4 revealed that larvae fed on commercial feed B had a significantly higher mortality than the other three diets (ANOVA, F 5 8.95; d.f. 5 3; P<0.001). Mortality of larvae fed on commercial feed B for days 5–8 and 9–12 were found to have a significantly higher mortality than larvae fed on Artemia, Artemia 1 commercial feed A and commercial feed A (ANOVA, F 5 6.12; d.f. 5 3; P=0.001). Larvae sampled at day 16 were found to have a significantly higher mortality when fed on commercial feed B when compared to larvae fed on Artemia, Artemia 1 commercial feed A and commercial feed A and larvae fed on commercial feed A were found to have a significantly higher mortality than larvae fed on Artemia and Artemia 1 commercial feed A (ANOVA, F 5 27.83; d.f. 5 3; P<0.001). Larvae fed on commercial diet B at days 20 and 24 were found to have a significantly higher mortality than larvae fed on Artemia, Artemia 1 commercial feed A and commercial feed A (ANOVA, F 5 9.19; d.f. 5 3; P=0.001).
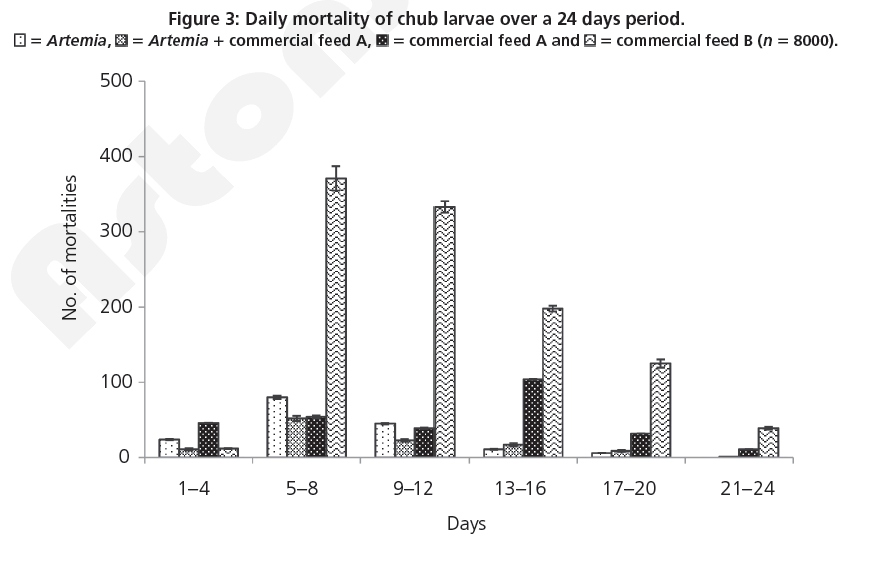
Figure 3: Daily mortality of chub larvae over a 24 days period.  = Artemia,
= Artemia,  =Artemia 1 commercial feed A,
=Artemia 1 commercial feed A, = commercial feed A and
= commercial feed A and  = commercial feed B (n 5 8000).
= commercial feed B (n 5 8000).
L. cephalus larvae fed on a mixed diet of Artemia 1 commercial feed A produced a similar growth rate (in length and weight) as those fed solely on Artemia, with similar survival, but not when fed on commercial feeds A and B in isolation. Similarly, it was revealed that compared to dry food, the feeding of decapsulated Artemia significantly increased the growth rates of L. cephalus and orfe (Leuciscus idus), although these studies did not also commercial fed live and dry feeds [5,19]. Previous studies have also recognised the problems with the use of dry diets in larviculture, producing similar results to this study regarding survival rates. For example, work on striped bass (Morone saxatilis) found larvae that only consumed dry feed failed to grow and subsequently, died soon after day 16 [20]. Whilst the reasons for the poor performance of commercial feed B in this study were not clear, they are likely to be associated with being more difficult to digest than Artemia and commercial feed A [20]. It is clear that the quality and digestibility of dry feeds is a big factor in larval survival.
The slower growth of the larvae on the dry foods is well known. However, Artemia nauplii have been shown to provide exogenous enzymes which can trigger or increase the proteolytic activity in the digestive tract of larvae [21,22] with the commercial micro-diets perhaps lacking these enzymes at the levels required to prepare the gut for digestion. The digestibility of these micro-diets has been shown to be affected by the method of manufacture and many larvae fed on dry starter feeds have been found to perish due to the gut being filled with undigested food [23]. Further factors that may have increased larval growth and survival when using live feed is that they usually have a much better contrast than artificial feeds and so produce a feeding trigger effect by their continuous movement [24], and that as Artemia actively swim, this ensures a good spatial distribution of food items resulting in more frequent encounters for all larvae [24]. Nevertheless, there have been some successes for larval production from feeding micro-diets after feeding live diets [25]. For example, a range of marine species that were commercial feed two live and dry foods demonstrated excellent larval growth and survival [26] although until now this had not yet been shown in any cyprinidae fish species.
This study revealed feeding L. cepahlus for 24 days from 5 days post-hatch with commercial feed produced significantly smaller larvae that suffered higher mortality rates compared to feeding with Artemia and a mix of Artemia with a dry feed. With little prior knowledge on the larviculture of this species, this output should help the process of formulating a larval rearing diet that produces optimal growth and survival rates that minimises resource costs and maximises the number of cultured fish that will ultimately be used in stocking to enhance wild fisheries. Further research, conducted on other cyprinid and non-cyprinid species, could reveal that the combination of dry and live feeds maintain the same growth rate of using solely Artemia due to the consistent quality of dry feeds.
The authors declare that they have no competing interests.
All author’s contributed equally to the production of this manuscript.
We would like to thank the staff at Calverton Fish Farm for their support and guidance in setting up the experiment.