Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 3
In order to better understand and elucidate the influence of rootstock/scion interactions under salinity constraint, three grafting combinations have been used: Superior Seedless/110R, Muscat d’Italie/SO4 and Syrah/1103 Paulsen. Superior Seedless/110R combination behaves and displays the same results as ungrafted Superior Seedless variety under both control and stress conditions. On the other hand, since the sensitive rootstock SO4 could improve the behavior of Muscat d’Italie variety against salinity, it highlights the beneficial effect of this combination even the two genotypes are not really tolerant. This gain of performance at both vigor and salinity tolerance levels would result from hybrid vigor. Likewise, even the rootstock 1103P is moderately sensitive, it reaches to improve the behavior of Syrah which is a very sensitive variety. Comparative analysis of the different combinations showed that rootstocks do not behave the same in cases they are used individually or in combination with scion. Indeed, the more or less sensitive rootstocks SO4 and 1103P (taken individually) behave better at the salinity tolerance level since combined with their respective scions. Moreover, varieties tested individually display a different pattern when used with a rootstock. Generally, they behave better (Muscat d’Italie and Syrah cases) or at least stay unchanged (Superior Seedless case) against salinity when they are grafted. The displayed behaviors among the different combinations reveal the existence of interactions between genotypes of rootstock and scion, and that not only rootstock dictates the global phenotype. These interactions enhance and improve generally the phenotype of the grafting combination. With a sensitive rootstock (whatever scion is, tolerant or sensitive), we always obtain a gain of performance which is more pronunciated with sensitive rootstocks.
Keywords: Grapevine; Grafting; Interaction; Salinity; Vigor
The practice of grafting, which combines a scion and a rootstock to form a new plant with a blend of characteristics, is a technique known from ancient times both in fruit trees and viticulture. It has been used historically to change variety, enhance vigor, or increase limestone tolerance. Since the late nineteenth century, grafting became a common practice in viticulture after the phylloxera epidemic. Grape growers usually select their rootstock for their vineyard mainly depending on their vigor and resistance to diseases, particularly phylloxera and nematodes; assuming that rootstock confers its properties to the scion. Practically, this is not as simple for some characters; indeed, extensive research on rootstocks revealed that several aspects of scion behavior such as adaptation to abiotic stresses were yet dependent on rootstock features but also related to rootstock-scion interactions. The different genotypes of grapevine tested by ref. [1] displayed different behaviours with regard to salinity, behaviors that oscillate between sensitivity and tolerance. However, genotypes that seem to better cope with abiotic stresses (salinity in particular) are more often susceptible to diseases or calcareous soils. For example, 140R genotype is considered to be the one who better resists to salinity; while SO4 is famous for its nematodes resistance, even very sensitive to saline and calcareous soils. By integrating the results of several authors works [1-10] the following classification of grapevine genotypes according to their degree of salinity tolerance could be postulated and used:
Superior Seedless>Muscat d’Italie>110R>1103P>Syrah>SO4
Our present work aims to the analysis of interactions between scions and rootstocks also under saline stress. Three grafting combination have been used, their choice was largely imposed by the availability of plant material. In a first attempt, analysis and interpretation of results will be done for each combination lonely; then, in order to better understand rootstock/scion relationship under saline condition, a comparative analysis will be performed within all combinations.
Plant material and culture procedure
Three varieties (Superior Seedless, Syrah and Muscat d’Italie) and 3 rootstocks (110 Richter, 1103 Paulsen and SO4) were used in our study according to the following grafting combinations:
- Superior Seedless/110 Richter corresponding to a tolerant variety grafted on moderately tolerant rootstock,
- Muscat d’Italie/SO4 corresponding to a moderately tolerant variety grafted on a very sensitive rootstock,
- Syrah/1103 Paulsen corresponding to a very sensitive variety grafted on a moderately sensitive rootstock.
Grafted cuttings are soaked at their base in exuberone solution (commercial IBA) in order to promote rhizogenesis, paraffined, then planted in pots filled with inert sandy soil and grown for 75 days in greenhouse under controlled conditions (25 ± 2°C temperature, 16 hr photoperiod, natural light and 70 ± 5% relative humidity). Irrigation (70% of field capacity) was made every two days with a nutrient solution composed by Long Ashton [11] macronutrients and [12] micronutrients. Iron was brought in the form of EDTA-K-Fe according to ref. [13].
Salt treatment
After two weeks of culture where cuttings started budding and initiating their root system and two additional weeks of acclimation period, rooted cuttings started to be irrigated with nutrient solution enriched with NaCl (100 mM), gradually applied by adding 25 mM NaCl every 15 days until reaching 100 mM.
Studied parameters
Biomass production: At the end of the culture, leaves, stems and roots are separately harvested. Samples are then weighed before and after drying at 60°C for 48 hours to determine their fresh and dry weights.
Foliar area and leaves number: Foliar area was determined for 4 leaves randomly taken from each level of the plant (basal, medium and upper) using a planimeter (Area Meter, type LI-3000A, LI-COR).
Chlorophyll content: Chlorophyll content is determined on fresh leaves at the final harvest of plants.
Mineral nutrition status: To study the pattern of ions transport and storage in whole plant, we determined the concentrations of mineral ions (sodium, chloride, potassium, calcium and magnesium) in the different organs (root, stem and leaves).
Methods
Index of sensitivity: Index of sensitivity is calculated according to the following formula [14]:
IS = 100 x (ΔMSNaCl − ΔMScontrol ) / ΔMScontrol
ΔMSNaCl : variation of dry weight production under NaCl treatment.
ΔMStémoin : variation of dry weight production under control condition.
Chlorophyll content determination: Chlorophyll content is determined on samples of fresh leaves harvested at the end of the culture from each genotype. Leaves are soaked with cooled acetone (100%) and then mixed using a polytron apparatus to a fine residue allowing extraction of chlorophyll pigments. A serial centrifugation and supernatant recovery steps is then performed until obtaining an extract whose absorbance is measured at a wavelength equal to 652 nm [15,16]. The evaluation of the chlorophyll content is done according to the following formula:
Total chlorophyll content (mg.g-1)=Acetone extraction volume (ml) × DO reading × 10/dry extract weight (g)
Mineral analyses: Samples of leaves, stems, and roots were ground to a powder in a stainless steel Dangoumeau grinder. After drying for 48 hr at 70°C, the powder was digested with a 4:1 (v:v) mixture of nitric and perchloric acids. The mixture was then filtered, diluted with distilled water, and analysed for mineral elements contents. K+, Ca2+, Na+ and Mg2+ contents were determined with atomic absorption spectroscopy (Perkin Elmer Atomic absorption spectrometer 3110). Cl- is determined, from the same acid extracts supplemented with acetic buffer (acetic acid 10 and, nitric acid 0.1N) and gelatin, using a digital chloridometre (type Haake Büchler) according to the principle of colorimetric titration.
K+/Na+ selectivity: The K+/Na+ selectivity is defined by the following formula:
SK+/Na+ = S1 / S2
S1 = ΔQK / (ΔQK+ ΔQNa)
S2 = [K+] / ([K+] + [Na+])
ΔQ : amount of element accumulated within the organ during the period of treatment (meq).
[---] : concentration of element (meq/ml).
Statistical analyses
Statistical Analysis System (1988) program has been used to perform all ANOVA analyses and Duncan’s test (α=0.05) to compare averages values for each analyzed parameter.
Plant growth: Biomass production:
Superior Seedless/110R combination: In control condition, the Superior Seedless variety produces as much biomass when cultivated on its own roots or grafted on the 110R, meaning that the use of 110R rootstock does not improve its growth (Figure 1). Own rooted Superior Seedless growth is significantly greater than 110R rootstock. Under salt stress condition (100 mM NaCl), there was a reduction of 50% biomass for this combination, as for own rooted Superior Seedless. Although the individually use of 110R rootstock leads to a growth reduction of 50%, its growth (estimated by its dry weight production) is significantly lower than the Superior Seedless/110R combination. Thus, the Superior Seedless/110R combination adopts the same behavior and displays the same results as the own rooted Superior Seedless variety under both control and stress conditions. In this particular case of grafting, the best genotype (i.e., Superior Seedless) imposes its phenotype to the rootstock/scion combination. Indeed, this combination remained as tolerant as the best genotype of rootstock and scion and its level of tolerance was not dictated by the weakest genotype of the combination nor by the rootstock. However, this combination may be considered very attractive, although not introducing any improvement of behaviour with regard to salt stress, as the 110R rootstock is resistant to nematodes and able to tolerate 17% of active limestone, character lacking in Superior Seedless scion.
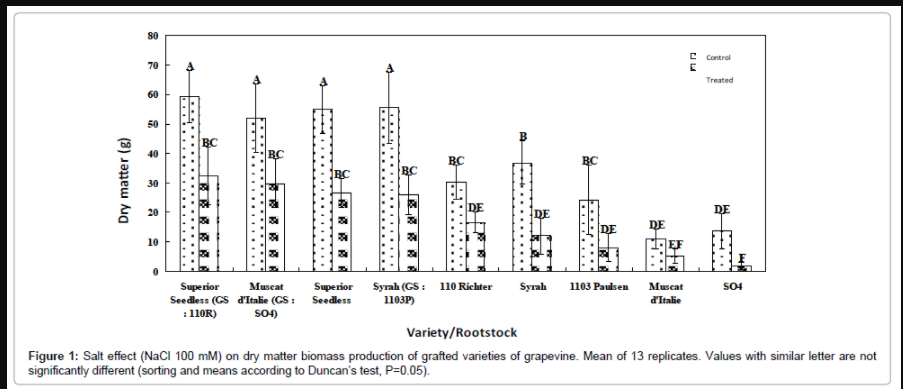
Figure 1: Salt effect (NaCl 100 mM) on dry matter biomass production of grafted varieties of grapevine. Mean of 13 replicates. Values with similar letter are not significantly different (sorting and means according to Duncan’s test, P=0.05).
Muscat d’Italie/SO4 combination: Under control condition, the use of the SO4 rootstock, although not renowned for its vigor, greatly improves the growth of Muscat d’Italie variety (Figure 1). Indeed, own rooted Muscat d’Italie produces barely 11 g of DW, while when grafted on the SO4, it reaches a production of DW 5 times higher (51 g). Under salt stress condition (100 mM NaCl), even there was a 45% reduction in DW production, this combination manages to grow much better and to generate 6 times more DW than the own rooted Muscat d’Italie (30 and 5 g, respectively). Thus, although the SO4 rootstock is very sensitive (the more sensitive among the tested rootstocks) it could improve the behavior of Muscat d’Italie grafted variety. Moreover, DW production of this combination under salt stress exceeds three-fold own rooted Muscat d’Italie cultivated under control condition ; which proves and emphasizes the beneficial effect of this combination even the genotypes are not classified as salt tolerant. This gain of performance, both in terms of vigor level and salinity tolerance could arise from the hybrid offspring. This increase in performance could be due to a genetic effect or a physiological stimulation mechanism.
Syrah/1103 Paulsen combination: Under control condition, the use of the weakly vigorous 1103P rootstock improves significantly the growth of Syrah variety (Figure 1). Indeed, grown on its own roots, Syrah produces 37 g of DW, while once grafted on the 1103P, it reaches 55 g. Under salt stress condition, there was a reduction in DW production of 60% for this combination and despite this considerable reduction, it still manages to grow much better than the own rooted grown Syrah variety and to produce twice its DW (25 and 12 g, respectively) and almost the same DW production of Syrah grown under control condition, meaning that this combination leads to a beneficial effect even both genotypes are more or less sensitive to salinity. Thus, although the 1103P rootstock is moderately sensitive it reaches to improve the behavior of the Syrah variety which itself is very sensitive. This combination also displays a gain of performance in terms of vigor and salinity tolerance.
Comparative analysis of the different combinations: It is a complicated task to make a comparison between all these different combinations due to the lack of suitable device. In practice, homogeneous combinations with a same rootstock tested with all varieties, and/or a single variety tested and grafted on different rootstocks would be a suitable experimental protocol; unfortunately, these combinations have failed due to the lack and the unavailability of plant material. The comparative analysis of the different combinations highlights first that rootstocks do not behave the same way whether they are used individually or in combination with scion. Indeed, more or less sensitive SO4 and 1103P rootstocks (individually) behave better in terms of salinity tolerance once in combination with their respective scions. Similarly, own root grown varieties display a different behavior once grafted on a rootstock; generally, their behavior improves (Muscat d’Italie and Syrah) or at least remains the same (Superior Seedless) under salt stress. Behaviors displayed by the different combinations reveal the existence of interactions between the different genotypes of rootstock and scion. These interactions lead generally to a better behavior of the rootstock/scion combination. On the other hand, with a sensitive rootstock (regardless of scion, tolerant or sensitive) there’s always a gain in performance of the resulting combination. This gain of performance is more pronounced as the more sensitive is the rootstock. All the rootstocks used in the three combinations resulted in a greater production of DW by the older leaves (basal and median) and therefore a better dilution effect on accumulated ions, showing thus the importance of the conferred vigor.
Leaves number: Under control condition, leaves number of own rooted Superior Seedless is the same as when grafted on 110R, either when they are subjected to salt stress, although they show a reduction comparatively to their respective controls. Superior Seedless grafting on 110R does not improve the number of leaves in plants regardless of the culture conditions. Grafting of Muscat d’Italie on SO4 allows increasing the number of leaf in both control and salt stress conditions compared to own rooted Muscat d’Italie. Concerning the Syrah / 1103P combination, no improvement in the number of leaves is recorded, whatever conditions are.
Leaf area: Grafting of Superior Seedless variety on 110R rootstock leads to an increase of the average leaf area which decreases under salt stress condition, in an equal value to that of the own rooted variety. Syrah/1103P combination does not display any leaf area modification in neither condition. On the other hand, the use of the SO4 rootstock improves the leaf area in all conditions.
Total chlorophyll content: Lowest chlorophyll levels are generally obtained for rootstocks, stressed or not. Grafting seems to be also beneficial to this parameter except in the case of the Muscat d’Italie/ SO4 combination which is in contradiction with results obtained for the other measured parameters. Thus, in control or salt stress conditions, the grafted varieties displays always best contents of chlorophyll.
Index of sensitivity: According to the index of sensitivity values shown in Figure 2, Superior Seedless grafted on 110R is the most tolerant combination, expected result since the Superior Seedless variety and 110R rootstock are the most tolerant genotypes used within our experiments. On the other hand, Figure 2 shows also that grafting improves the salinity tolerance and therefore decreases the salt sensitivity of the variety. Thus, a ranking of salinity tolerance could be established among these combinations: Seedless/110R>Syrah/1103P>Muscat d’Italie/SO4. This result clearly indicates the importance of grafting and rootstock effect in grapevine salinity tolerance acquirement. Our study demonstrated that a sensitive variety (Syrah) grafted on a moderately tolerant rootstock (1103P) becomes more tolerant and better behaves under salinity constraint than a moderately tolerant variety (Muscat d’Italie) grafted on a sensitive rootstock sensitive (SO4). Thus, in our particular case, the rootstock confers its ability and character of salinity tolerance especially to a sensitive variety. On the other hand, a more or less tolerant variety like Muscat d’Italie loses its tolerance once grafted on a sensitive rootstock, comparatively to another combination of grafting characterized by a sensitive variety grafted on a more or less tolerant rootstock, and to the same variety, because we have already demonstrated that in all cases the grafting improves the behavior of the variety under salinity condition.
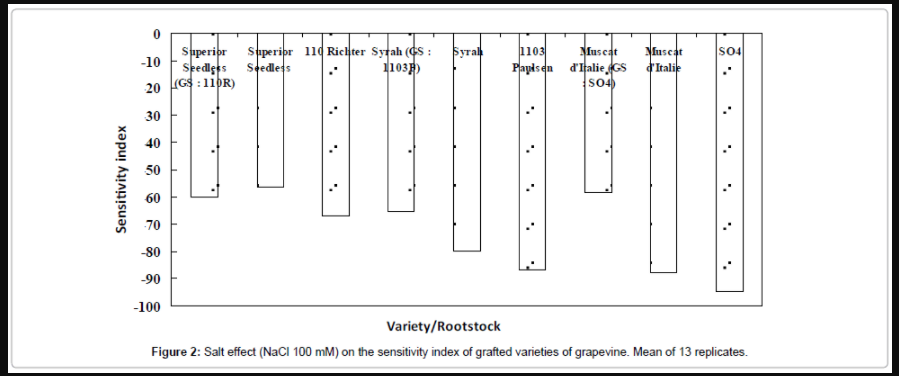
Figure 2: Salt effect (NaCl 100 mM) on the sensitivity index of grafted varieties of grapevine. Mean of 13 replicates.
Plant mineral nutrition: NaCl effect on sodium accumulation:
Superior Seedless/110R combination: Salt treatment (100mM NaCl) induces increase in sodium concentrations in all organs, particularly, in leaves where this combination shows an increase in sodium concentration and thereby displays a higher capacity of sodium accumulation than the 110R rootstock but a lower one in comparison with the own rooted Superior Seedless variety. Thus, Superior Seedless, when grafted on 110R, displays a foliar content of sodium greater than that of the rootstock (cultivated on his own roots) and lower than the own rooted scion (Figure 3). At the stem and root levels, no difference was recorded between this combination, rootstock and scion; indicating that grafting of Superior Seedless on 110R does not change nor improve capacity of sodium accumulation in stem or roots. In conclusion, 110R rootstock sets to the combination its sodium storage capacity, with a slight increase in leaves (Figure 3).
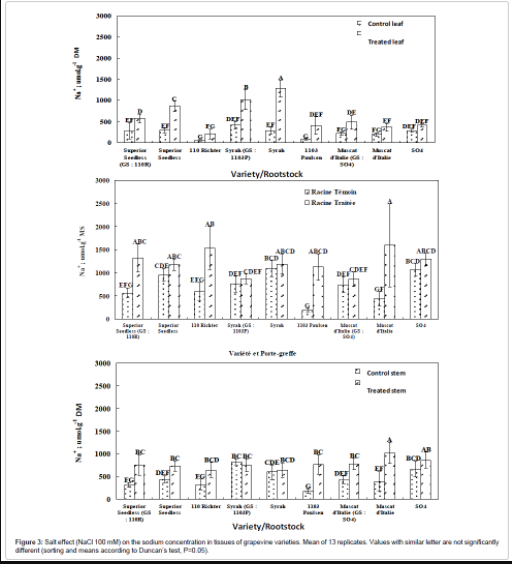
Figure 3: Salt effect (NaCl 100 mM) on the sodium concentration in tissues of grapevine varieties. Mean of 13 replicates. Values with similar letter are not significantly different (sorting and means according to Duncan’s test, P=0.05).
Muscat d’Italie/SO4 combination: For this combination, NaCl treatment induced an increase in the concentration of sodium only in leaves and stem but not in roots. Moreover, its foliar sodium content was identical to those of Muscat d’Italie and SO4 cultivated individually. This kind of grafting allows the plant to display sodium concentrations in stem or roots equally to those of the rootstock (cultivated alone) but never reaches those of own rooted scion (Figure 3). Once again, the rootstock imposes its sodium storage ability to the combination.
Syrah/1103P combination: In comparison with control samples, this combination shows an increase in sodium concentration restricted to leaf compartment and neither within stem nor roots whose sodium accumulation capacity stayed rather identical to 1103P rootstock (individually cultivated) and own rooted Syrah. However, even this combination performs better than 1103P (i.e., displays a higher concentration of sodium in leaves), it is less efficient than own rooted Syrah which accumulates nearly 1300 μmol/g MS of Na+ in its leaves. Thus, for all tested combinations, foliar sodium accumulation capacity is improved compared to the rootstocks, and this strategy of sodium inclusion in leaves is characteristic of tolerant rootstocks. On the other hand, the behavior of sodium exclusion within roots, which is specific to tolerant varieties, is not transmitted to the combination of grafting. In control conditions, the use of the different rootstocks does not alter sodium content in all organs. Furthermore, the combinations Muscat d’Italie/SO4 and Syrah/1103P display a preferential accumulation of sodium in old leaves, in addition to the dilution effect.
NaCl effect on chloride accumulation:
Superior Seedless/110R combination: This combination adopts the chloride accumulation pattern of 110R rootstock particularly at the root level; however at the foliar level, for which the 110R rootstock displays very high concentrations of chloride, result that fails to reproduce Superior Seedless grafted on 110R, it maintains the same pattern of own rooted Superior Seedless. Thus, it seems that this combination, for its aerial part, mimics the behavior of scion, while for its root part, it follows the rootstock (Figure 4). The strategy of chloride exclusion in the root compartment, specific of tolerant rootstocks, is therefore transmitted by the 110R rootstock to the Superior Seedless/110R combination.
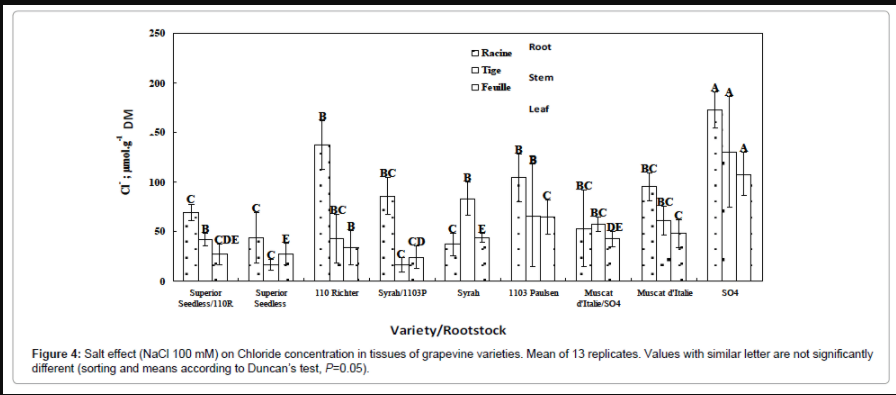
Figure 4: Salt effect (NaCl 100 mM) on Chloride concentration in tissues of grapevine varieties. Mean of 13 replicates. Values with similar letter are not significantly different (sorting and means according to Duncan’s test, P=0.05).
Syrah/1103P combination: The chloride concentration obtained for this combination within roots and stems is significantly lower than those of Syrah and 1103P cultivated individually (Figure 4). Although, at the foliar level, this combination follows the behavior of the 1103P rootstock and displays higher chloride concentration than own rooted Syrah.
Muscat d’Italie/SO4 combination: This type of grafting displays, at all organs levels, chloride concentrations similar to those of the own rooted Muscat d’Italie but lower than those of the rootstock (individually cultivated). The SO4 rootstock was unable to transmit its capacity of chloride accumulation to this combination (Figure 4). Therefore SO4 rootstock, because of its phenotypic sensitivity, does not influence the capacity of chloride accumulation within the Muscat d’Italie/SO4 combination. In conclusion, grafting procedure led to an increase of chloride storage particularly within roots and stems for all combinations, and there was not a preferential distribution of chlorides in the older leaves.
NaCl effect on potassium accumulation
Superior Seedless/110R combination: Potassium concentration increases in leaves of this combination under salt stress (in comparison with control condition) and follows that of 110R rootstock whose value is greater than the own rooted Superior Seedless. Paradoxically, in stem and roots, potassium concentrations under salt stress trend downward (Figure 5). These results may suggest and confirm the osmoregulatory role played by potassium within leaf cells. Increase in leaves potassium concentration was only registered for this combination, thus explaining its adaptation and its tolerance to salt stress.
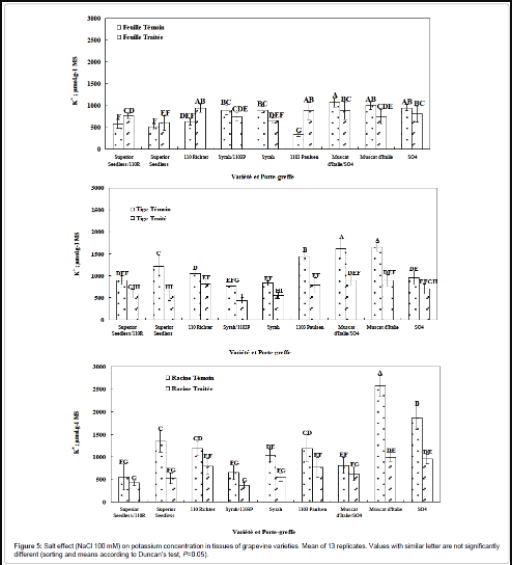
Figure 5: Salt effect (NaCl 100 mM) on potassium concentration in tissues of grapevine varieties. Mean of 13 replicates. Values with similar letter are not significantly different (sorting and means according to Duncan’s test, P=0.05).
Syrah/1103P combination: For this combination, potassium levels decreased under salt stress in all parts of the plant (Figure 5). Although the 1103P rootstock increases its potassium concentration in leaves, once associated with Syrah, he was unable to transmit his capacity of potassium accumulation in syrah leaves. The 1103P rootstock could not improve the behavior of the Syrah variety following grafting.
Muscat d’Italie/SO4 combination: We also found for this combination a decrease in potassium levels in all parts of the plant under salt stress (Figure 5), indicating lack of grafting effect, and where both rootstock and scion displayed the same behavior and potassium concentrations. Grafting has not changed the levels of potassium in the aerial part under control condition, but has resulted in their decrease within roots in comparison with both own rooted Muscat d’Italie and SO4 rootstock.
NaCl effect on calcium accumulation: Concentrations of foliar calcium increase under salt stress for all grafting combinations while within the other organs, no significant change were observed. Cellular signaling via calcium cations remains an important factor in response to salt stress. Under control condition, calcium contents are not modified following grafting excepting for Superior Seedless/110R combination.
NaCl effect on magnesium accumulation: Following salt stress and for all combinations, magnesium concentrations increase, especially in the aerial part of the plant (stem and leaves), indicating that grafting improves the capacity of magnesium accumulation, but there is no change under control condition.
K/Na ratio
In all organs and for all combinations, K/Na ratio shows a decline under salt stress. This ratio was not improved by grafting, it increases in 1103P leaves. We have already seen that grafting improves sodium concentrations towards a better salinity tolerance, but has no influence on potassium concentrations excepted for Superior Seedless/110R combination leaves, but insufficiently to modify the K/Na ratio.
Studies concerning the relationship between grape varieties, rootstocks and environment have, in viticulture, special importance due to crop yield impact and plant adaptation [17]. Our work intends to study interactions between rootstock and scion in different grafting combinations under salt stress condition. Development and growth of the whole plant, physiological parameters, as well as plant mineral content were addressed. Assumptions have been suggested concerning the influence and modalities of action of the rootstock on scion’s behavior and performance. In grafting combination, the rootstock interacts with the scion to promote the developmental characteristics of the whole plant [18]. From a physiological point of view, the rootstock is the link between the soil and the plant and therefore it is responsible for mineral and water uptake and transport towards the scion. Rootstock is also known to confer certain vigor to the scion, resulting from an interaction between rootstock and Scion [19]. This conferred vigor can be greater or lower than that of the own rooted scion [18]. The determinism of the vigor conferred by the rootstock is not clearly understood and the underlying mechanisms involved are still under investigation. Generally, the rootstock/scion relationship is mainly focused on the rootstock side, as responsible and main factor of the resultant vigor of grafting; while our results highlight the influence of the scion on the phenotype and the final behavior of the combination. Although grapevines are classified as moderately sensitive to salinity according to ref. [20], a great variability of behaviours was observed among rootstocks and varieties, in field or greenhouse. Globally, cultivated species V. vinifera are less tolerant to salinity than those used as rootstocks [7,21,22]. Sultana grapevine growing on its own root system, or else grafted to Dogridge, 1613, Harmony and Ramsey rootstocks, and irrigated with 75 mM solutions of chloride salts under glasshouse conditions for three consecutive growing seasons, displayed growth reduction that was less pronounced with Vitis champini rootstocks [23]. All used rootstocks, with the exception of Dogridge, allowed to reduce the foliar concentrations of chloride below that of self-rooted Sultana. Rootstocks lowered sodium, particularly in petioles and raised potassium concentrations in all plant parts [23]. In the same way, grafting Cardinal and Sultana varieties on Dogridge, 1163-3 and Salt Creek rootstocks led to a reduction in foliar concentrations of chloride in comparison with own rooted varieties [2]. On the other hand, grafting of Shiraz variety on Ramsey, 1103P and 140R rootstocks helped to improve the levels of potassium [24]. Several authors [6,22,24] particularly recommend the use of Ramsey and 1103P rootstocks, due to their conferred vigor, with Sultana variety grown under salinity conditions. Rootstocks like Ramsey, 140R and 1103P grafted with Syrah have shown lower chloride concentrations in berry juice than own rooted plants; however, some rootstocks like K51-40 gave higher concentrations of chloride and sodium in berry juice when irrigated with saline water (2.1 dS/m). Chloride exclusion ability of Ramsey and 1103P rootstock decreased upon several seasons of irrigation with saline water (2, 1 ds/m), whereas that of 140R remains constant [25]; confirming our results concerning the skills and Cl- excluding potentiality of this last rootstock. Several authors highlighted the influence of rootstock on scion photosynthetic capacity [26-28]. For example, grafting Pinot noir variety onto the SO4 rootstock led to a reduction in leaf chlorophyll content as well as a decrease in the levels of mineral elements P, K, Ca, Mg and Fe [29]. The Chardonnay variety once grafted on the SO4 displayed a bigger reduction of photosynthetic activity, stomatal conductance, and chlorophyll levels than that on the 1103P [26,30] related also that these effects were specific to the rootstock/scion combination. In grapevine, the relationship between drought tolerance and conferred vigor is not strongly demonstrated; however, since cell growth is among the most sensitive processes to the plant water status, it is not likely to disconnect the vigor from water status of cell [31]. In ref. [32] and Lider did not found any influence of rootstock on the leaf content of calcium and magnesium of plants under normal condition, and our work confirms these results; although ref. [33] found slight modification of the leaf levels of minerals under the influence of rootstock. Each rootstock own specific capacities of ion absorption and distribution and variable affinities with mineral elements [34]. However, most often, measurements have been taken on the petiole or limb. When comparing Cabernet Sauvignon grafted onto 11 rootstocks, same authors did not find any link between vigor and mineral nutrition and therefore no effects of dilution by the vigor (=most vigorous plants absorb more minerals) were seen. It is now clearly established that there are exchanges of non-trophic substances between the root system and aerial part [18]. These exchanges take place via the xylem or phloem. The supply of growth regulators such as cytokinins and abscisic acid by the root system may be involved in several mechanisms governing relations between roots and aerial part of the plant [35]. In ref. [36] observed an effect of rootstock on the xylem sap content of cytokinins within Sultana variety. In this context, 110R rootstock better behaves than the 1103P, supporting thus our results. Also, it has been shown that proteins and nucleic acids could translocate between different parts of a grafted plant via the phloem sap [37]. Several signaling molecules, moving through phloem stream, could be involved in gene expression regulation. The involvement of RNA trafficking in phloem-mediated signaling has been implicated by the recent discovery that posttranscriptional gene silencing (PTGS) can operate on a whole-plant level [38]. Grafting studies provided unequivocal proof that the propagation of PTGS within the plant represents a non-cell/organ-autonomous event. A sequence-specific signal for nitrate/nitrite reductase was shown to move from a silenced rootstock to non-silenced scion where it induced silencing of only this specific gene [39]. Among all genes that may be involved in the regulation of growth process and water status of plants, those encoding aquaporins and ion carriers might be particularly interesting [18,40]. Indeed, their study would help elucidating at the molecular level the mechanisms of adaptation of grafted grapes through their water status and mineral nutrition. The works of ref. [25] confirm our findings concerning the fact that 1103P rootstock improves the scion behavior (especially Sultana) with regard to salinity. Experiments with Sultana variety grafted on different rootstocks (Ramsey, 1103P, J17-69 and 4 other hybrids) growing under salt stress (0.40, 1.75 and 3.50 dS/m NaCl) led to a reduction in chloride foliar concentrations, by behaving as excluder in comparison to the own rooted Sultana; and where the 1103P rootstock was the best chloride excluder (Walker et al.). Thus, these authors concluded that the strategy of ions exclusion as well as the conferred vigor by the rootstock represent major factors for salinity adaptation. The grafting of the Pinot Blanc variety on the 140R reduces ferric chlorosis on calcareous soils unlike the use of 101-14 (V. riparia × V. rupestris) rootstock, proving that 140R would have a high capacity of iron uptake [41]. Using Ramsey rootstock with several varieties (Muscat d’Alexandrie, Chardonnay, Syrah, Cabernet Sauvignon and Riesling), ref. [7,21] found a positive correlation between potassium content in berries and grafting, some combinations displaying better affinities than others, such as Syrah grafted on Ramsey. This is consistent with our results, especially with the matter that grafting of a sensitive variety such as Shiraz is always beneficial, but also that there are affinities between scion and rootstock to be considered for grafting success. In ref. [17], it highlighted a significant interaction between rootstocks and varieties concerning yields and sugar accumulation in berry. Under same conditions, a rootstock is able to behave differently, for a same feature, according to the variety with which it is associated. Indeed, Ramsey rootstock is sometimes able to reduce root concentrations of chlorides with Syrah and Sultana scions, and sometimes able to increase these concentrations with the Ruby Cabernet scion [42]. Moreover, according to ref. [43], the study of water stress adaptation should be performed on grafted plants to take into account the mechanisms of rootstock/scion interactions. Ref. [44] considers that the mechanism of drought adaptation induced by the rootstock to the combination would be a better water uptake capacity through the root system. However, the determinism of drought tolerance of a rootstock/scion combination is not yet completely understood. According to Paul and Delas (1982), leaf mineral content in the scion genotype may be considered as the result of two physiological properties:
- The uptake ability of the root system, which is characteristic of the rootstock genotype,
- The accumulation capacity of the leaf blade and the transit ability of the petiole, which are characteristic of the scion. These latter may be more or less stable, while the uptake ability of the rootstock is varying. However, the uptake ability of a rootstock may be influenced by the accumulation capacity and the transit ability of scion (Paul and Delas, 1982). Ref. [19] have shown that growth under salt stress of the own rooted Sultana variety exceeded those of Sultana grafted onto 110R, 41B, 140R, 1103P and SO4 rootstocks. These results are in contradiction with what we obtained with Muscat d’Italie and Syrah varieties, but are similar to those obtained by the Superior Seedless, confirming thus the importance of affinity between rootstock and scion. Grafting therefore not always improves the behavior of the scion to salinity, but it also depends on the combinations and affinity between rootstock and scion. Although, the levels of sodium and chloride have been reduced due to the use of all rootstocks, 41B then the 1103P being those which allow the highest accumulation of chloride in tissues ; 140R, being the one who allows the highest accumulation of sodium. The Sultana variety grafted on SO4 displays the lowest sodium concentrations in the roots as well as the lowest photosynthetic activity [45]. All tested rootstock resulted in Na+ and Cl- exclusion [45], particularly V. berlandieri genotypes and their descendants unless the hybrid has a V. vinifera parent [5], and which explains the weak exclusion capacities of 41B (V. berlandieri × V. vinifera). So, despite the high contents of sodium and chloride, own rooted Sultana behaves better with regard to salinity than when it is grafted on 110R, 41B, 140R, 1103P and SO4 rootstocks [45].
In terms of salinity tolerance and grafting influence, general rules can be concluded from our work:
-Using two similar genotypes in the grafting combination would not bring significant improvement. Indeed, as shown by the Superior Seedless/110R combination, the best genotype (Superior Seedless, although being a scion) imposes its phenotype on the final behavior of the grapevine plant. This combination remained as tolerant as the best genotype of rootstock and scion and its level of tolerance was not dictated by the lowest genotype of the combination nor by the rootstock.
-Using two genotypes behaving differently under salinity (like Muscat d’Italie, tolerant scion, grafted on SO4, very sensitive rootstock) would improve very well the resulting phenotype of the grafting combination. Thus, although SO4 is a very sensitive rootstock, it manages to improve the behavior of Muscat d’Italie, which is already a tolerant variety. However, in our work because of lack of plant material, this gain of performance has not been proved for a very sensitive scion grafted on a tolerant rootstock; yet a very well-known dogma among grape growers and scientists. In addition, according to the results of the third combination, grafting would be also benefit if the rootstock is slightly more tolerant than the scion. Indeed, although the 1103P rootstock is moderately sensitive it could improve the behavior of the Syrah variety which itself is very sensitive.
Table 1 lists and summarizes the various physiological parameters modified by grafting over scion cultivated on his own roots.
| Relative growth rate | Dry matter production | Leaves number | Total leaf area | Chlorophyll content | Water content | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | S | C | S | C | S | C | S | C | S | C | S | |
| Superior Seedless / 110R | - | - | O | O | O | O | + | O | + | + | O | O |
| Muscat d’Italie / SO4 | ++ | ++ | ++ | ++ | ++ | ++ | + | + | O | O | O | O |
| Syrah / 1103P | - | - | + | + | O | O | O | O | + | + | - | - |
Table 1: Changes on physiological parameters of various grapevine grafting (variety/rootstock) in comparison with the variety grown on its own roots. C: Control; S: Salt stress; O: No change; -: Regression; +: Improvement.
-At the nutritional level, for the three tested combinations, the foliar sodium accumulation capacity (highly elevated in tolerant rootstock) is improved in comparison with the rootstock. However, sodium exclusion within roots, which is a specific strategy of tolerant varieties, is not transmitted to the combination of grafting. A preferential accumulation of sodium in older leaves is observed within positive combinations. Chloride exclusion within roots, specific behavior of tolerant rootstocks, could be transmitted to the Superior Seedless/110R combination by the 110R rootstock, but not to the Muscat d’Italie/ SO4 combination by the SO4 rootstock, since this later is very sensitive. Concerning potassium cation, only the 110R, best of tested rootstocks, allowed to transmit to the scion its strong aptitude of leaf potassium accumulation needed by plant for osmotic adjustment.
-Comparative analysis of the different combinations highlights that rootstocks do not behave the same way depending on whether they are used individually or in combination with scion. Indeed, the more or less sensitive SO4 and 1103P rootstocks (individually) behave better in terms of salinity tolerance once in combination with their respective scions. Similarly, individually tested varieties display a different behavior once they are used with a rootstock. Generally, when they are grafted, their behaviour is improved (like Muscat d’Italie and Syrah) or at least remains the same (like Superior Seedless) under saline constraint.
-Behaviours displayed by the different combinations reveal and highlight the existence of interactions between the different genotypes of rootstock and scion. These interactions are generally favorable to a better behavior of the grafting combination. Moreover, when the rootstock is sensitive (and whatever is the scion, tolerant or sensitive) there’s always a gain of performance within the grafting combination. The more the rootstock is sensitive, the more this gain is bigger
-Our work led to the following salinity tolerance ranking (according to the index of sensitivity) of the grafted varieties:
Superior Seedless/110R>Syrah/1103P>Muscat d’Italie/SO4
This result clearly shows the importance of grafting and scion/ rootstock interaction in grape salinity tolerance acquirement. It demonstrates that a sensitive variety (Syrah), when grafted on a moderately tolerant rootstock (1103P) becomes more tolerant and better behaves under salt stress than a moderately tolerant variety (Muscat d’Italie) grafted on a sensitive rootstock (SO4); meaning that the rootstock could confer its ability of tolerance to a sensitive variety. Moreover, in comparison with a sensitive variety grafted on a moderately tolerant rootstock, a more or less tolerant variety (Muscat d’Italie) loses its tolerance once grafted on a sensitive rootstock.