Drug Designing: Open Access
Open Access
ISSN: 2169-0138
ISSN: 2169-0138
Research Article - (2015) Volume 4, Issue 1
Alfalfa mosaic virus (AMV) infects over 600 plant species in 70 families (experimental and natural hosts). Coat proteins are important materials for making structure-function correlations with biologically active peptides on the physicochemical and immunochemical levels and also are good models for observing the evolutional changes in protein molecules. Antigenic peptides at position 1-MSSSQKKAGGKAGKPTKRSQN-21; 151-PTHAGMQNQNF-161; 22-YAALRKAQLPKPPALKVPVVKPT-44 of Alfalfa mosaic virus coat protein are most suitable for subunit vaccine development because with single epitope, the immune response can be generated in large population. In this research, we used PSSM and SVM algorithms for the prediction of MHC class I & II binding peptide, antigenicity, Solvent accessibility, polar and nonpolar residue to analyse the regions that are likely exposed on the surface of proteins which are potentially antigenic that allows potential drug targets to identify active sites against infection as well as to design synthetic peptide vaccine.
<Keywords: Alfalfa mosaic virus, Antigenic peptides, MHC-Binders, TapPred, PSSM, SVM, Nonamers
Alfalfa mosaic virus with bacilliform particles of different lengths, the largest usually c. 60 nm; in which four species of single-stranded RNA of messenger polarity are separately packaged. The three largest RNA species comprise the genome; the fourth is a sub-genomic messenger for the coat protein. The three genome RNA species and either the fourth RNA or the coat protein are needed for infectivity. Alfalfa mosaic virus (AMV) infects over 600 plant species in 70 families (experimental and natural hosts). Some hosts are potato (Solanum tuberosum), pea (Pisum sativum), tomato (Lycopersicon esculentum), tobacco (Nicotiana tabacum), bluebeard (Caryopteris incana). The virus is readily sap-transmissible, is seed-transmissible in some hosts and is transmitted in the non-persistent manner by aphids to a very wide range of host plants [1]. AMV infection symptoms are vary from wilting, mottles, white flecks, ringspots, malformation like dwarfing, mosaics to necrosis depending on the virus strain, host variety, stage of growth at infection and environmental conditions. Symptoms of infection can persist or disappear quickly. Alfalfa mosaic virus can be detected in each part of the host plant. The virions are mainly found in the cytoplasm and chloroplast of the infected plant as inclusion bodies.
Alfalfa mosaic virus (AMV) and ilarvirus RNAs are infectious only in the presence of the viral coat protein. To understand the coat protein’s function is important for defining viral replication mechanisms. In vitro replication experiments shows the conformational switch model states that AMV coat protein blocks minus-strand RNA synthesis, while another research states that coat protein present in an inoculum is required to permit minus-strand synthesis [2,3]. Alfalfa mosaic virus causes various mosaics, mottles and malformations in lucerne (alfalfa; Medicago sativa) but is often symptomless in this host, especially during summer, and is most prevalent in old crops. AMV causes calico and tuber necrosis in potato. Various symptoms in tobacco and garden lupin, yellow fleck in Caryopteris incana, white mottle in Philadelphus sp., mosaic in Malva parviflora and Viburnum opulus. It causes of mosaic in red and white clover, celery, celeriac and lettuce, of yellow mosaic in cowpea (Vigna unguiculata), mung bean (V. radiata), bean (Phaseolus vulgaris), and chilli pepper (Capsicum annuum), of necrosis and stunting in pea (Pisum sativum), of severe necrosis in tomato, and of wilting in chickpea (Cicer arietinum). It found naturally in many wild and cultivated species [4-6].
Coat proteins are important biologically active peptides on the physicochemical and immunochemical levels and also are good models for observing the evolutional changes in protein molecules. Antigenic peptides from Alfalfa mosaic virus are most suitable for the development of subunit vaccine because a single toxin subunit can generate sufficient immune response. Major histocompatibility complex (MHC) molecules are cell surface proteins that binds to the peptides derived from host or antigenic proteins, and present them at the cell surface for recognition by cells. Cell recognition is a fundamental mechanism of the immune system by which the host identifies and responds to foreign antigens [7,8]. There are two types of MHC molecule and are extremely polymorphic. MHC class I molecules present peptides from proteins synthesized within the cell, whereas, MHC class II molecule present peptides derived from endocytosed extracellular proteins. Identification of MHC-binding peptides and epitopes helps improve our understanding of specificity of immune responses [9-13].
Virus transmission
The virus is transmitted by thrips, which have a wide range of hosts. The virus survives in these hosts and acts as a source of inoculums for the vector. The thrips are carried by wind. The population of vectors increases rapidly from January-March and August-September Kharif and hence the crop suffers a heavy loss in both the seasons. A prolonged dry spell favours the multiplication of thrips and spread of the virus.
Strategy
The phenotype of the resistant transgenic plants includes fewer centres of initial virus infection, a delay in symptom development, and low virus accumulation. Protoplasts from virus resistant transgenic plants are also resistant, suggesting that the protection is largely operational at the cellular level. Transgenic plants expressing nucleocapsid protein are protected against infection by virus particles but are susceptible to viral RNA, indicating that the protection may primarily involve an inhibition of virus uncoating. This approach is based on the phenomenon of cross-protection [14] hereby a plant infected with a mild strain of virus is protected against a more severe strain of the same virus. Proteins of soybean mosaic virus are necessary for its production in or on all food commodities. An exemption from the requirement of a tolerance is established for residues of the biological plant pesticide.
MHC class binding peptides
The new paradigm in vaccine design is emerging, following essential discoveries in immunology and development of new MHC Class-I binding peptides prediction tools [15]. MHC molecules are cell surface glycoproteins, which take active part in host immune reactions. The involvement of MHC class-I in response to almost all antigens and the variable length of interacting peptides make the study of MHC Class I molecules very interesting. MHC molecules have been well characterized in terms of their role in immune reactions. They bind to some of the peptide fragments generated after proteolytic cleavage of antigen [16]. This binding acts like red flags for antigen specific and to generate immune response against the parent antigen. So a small fragment of antigen can induce immune response against whole antigen. Coat protien peptides are most suitable for subunit vaccine development because with single epitope, the immune response can be generated in large population. MHCpeptide complexes will be translocated on the surface of antigen presenting cells (APCs). This theme is implemented in designing subunit and synthetic peptide vaccines [22]. One of the important problems in subunit vaccine design is to search antigenic regions in an antigen [17] that can stimulate T cells called T-cell epitopes. In literature, fortunately, a large amount of data about such peptides is available. Pastly and presently, a number of databases have been developed to provide comprehensive information related to T-cell epitopes [18-21].
Database searching
The antigenic protein sequence of coat protein of Alfalfa mosaic virus was retrieved from GenBank, UniProt databases are initially the most important [AAA46297] [22-28].
Prediction of antigenicity
Prediction of antigenicity program predicts those segments from coat protein that are likely to be antigenic by eliciting response. In this research work antigenic epitopes of coat protein of Alfalfa mosaic virus are determined by using the Gomase-Kale Method, Hopp and Woods, Welling, Parker, Bepipred , Kolaskar and Tongaonkar antigenicity methods [29-35].
Predict protein-protein binding sites
Profisis method (ISIS) is a machine learning-based method that identifies interacting residues from sequence alone. Although the method is developed using transient protein–protein interfaces from complexes of experimentally known 3D structures, it never explicitly uses 3D information. Instead, we combine predicted structural features with evolutionary information [36,37].
Prediction of MHC binding peptide
The major histocompatibility complex (MHC) peptide binding of coat protein of Alfalfa mosaic virus is predicted using neural networks trained on C terminals of known epitopes. Rankpep predicts peptide binders to MHC-I ligands whose C-terminal end is likely to be the result of proteosomal cleavage using Position Specific Scoring Matrices (PSSMs). Support Vector Machine (SVM) based method for prediction of promiscuous MHC class II binding peptides from protein sequence; SVM has been trained on the binary input of single amino acid sequence [38-42].
Prediction of antigenic peptides by cascade SVM based TAPPred method
In the present study, we predict cascade SVM based several TAP binders which was based on the sequence and the features of amino acids [43]. We found the MHCI binding regions, the binding affinity of coat protein of Alfalfa mosaic virus.
Solvent accessible regions
We also predict solvent accessible regions of proteins having highest probability that a given protein region lies on the surface of a protein Surface Accessibility, backbone or chain flexibility by Emini et al. [44] and Karplus and Schulz [45]. By using different scale we predict the hydrophobic and hydrophilic characteristics of amino acids that are rich in charged and polar residues i.e. Gomase-Kale method, Sweet et al., Kyte & Doolittle, Abraham & Leo, Bull and Breese, Guy, Miyazawa, et al., Roseman, Wolfenden et al., Wilson et al., Cowan, Chothia [46-57].
Coat protein of Alfalfa mosaic virus contain a long residue with 221 amino acids [AAA46297].
MSSSQKKAGGKAGKPTKRSQNYAALRKAQLPKPPALKVPVVKPTNTILPQT GCVWQSLGTPLSLSSFNGLGARFLYSFLKDFVGPRILEEDLIYRMVFSITPSHAGTF CLTDDVTTEDGRAVAHGNPMQEFPHGAFHANEKFGFELVFTAPTHAGMQNQN FKHSYAVALCLDFDAQPEGSKNPSFRFNEVWVERKAFPRAGPLRSLITVGLFDEADDLDRH
Prediction of antigenic peptides
Predict protein-protein binding sites are strongest predictions of the method reached over 90% accuracy in a cross-validation experiment. Our results suggest that despite the significant diversity in the nature of protein–protein interactions, they all share common basic principles and that these principles are identifiable from sequence alone (Figure 1).
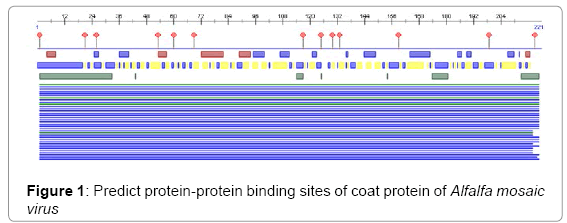
Figure 1: Predict protein-protein binding sites of coat protein of Alfalfa mosaic virus
In this study, we found the antigenic determinants by finding the area of greatest local hydrophilicity. The Hopp-Woods scale of hydrophilicity prediction result data found high in 165-173 [MIN: -2.633, MAX: 1.767] in a protein, assuming that the antigenic determinants would be exposed on the surface of the protein and thus would be located in hydrophilic regions (Figure 2). Welling antigenicity plot gives value as the log of the quotient between percentage in a sample of known antigenic regions and percentage in average proteins and prediction result data found high in position 13-41[MIN: -1.248, MAX: 0.873] (Figure 3). We also study Hydrophobicity plot of HPLC / Parker Hydrophilicity prediction result data found 112-DDVTTED-118, Score: 6.357 (maximum) [Average: 1.376, Minimum: -4.571, Maximum: 6.357] (Figure 4), BepiPred predicts the location of linear B-cell epitopes Result found that 1-MSSSQKKAGGKAGKPTKRSQN-21; 151-PTHAGMQNQNF-161 (Maximum), [Average:0.227 Minimum:-1.937 Maximum:2.305 Threshold: 0.350] (Figure 5, Table 1), Kolaskar and Tongaonkar antigenicity methods (Figure 6, Table 2) predicted peptides result found i.e., 165-YAVALCL-171, Score :1.226 (maximum) [Average: 1.029 Minimum: 0.907 Maximum: 1.226 Threshold: 1.000] and the predicted antigenic fragments can bind to MHC molecule is the first bottlenecks in vaccine design.
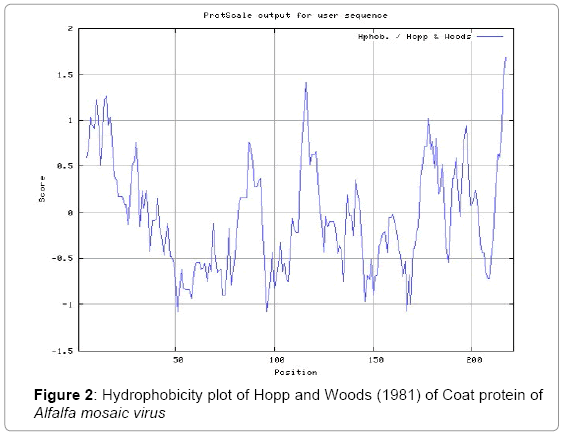
Figure 2: Hydrophobicity plot of Hopp and Woods (1981) of Coat protein of Alfalfa mosaic virus
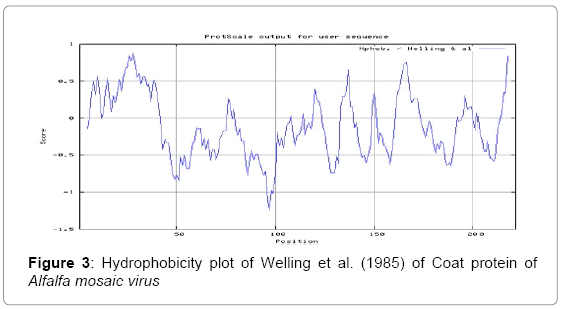
Figure 3: Hydrophobicity plot of Welling et al. (1985) of Coat protein of Alfalfa mosaic virus
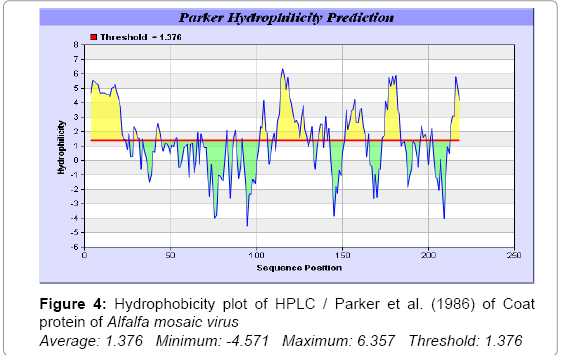
Figure 4: Hydrophobicity plot of HPLC / Parker et al. (1986) of Coat protein of Alfalfa mosaic virus Average: 1.376 Minimum: -4.571 Maximum: 6.357 Threshold: 1.376
| No. | Start Position | End Position | Peptide | Peptide Length |
|---|---|---|---|---|
| 1 | 1 | 21 | MSSSQKKAGGKAGKPTKRSQN | 21 |
| 2 | 29 | 37 | QLPKPPALK | 9 |
| 3 | 41 | 49 | VKPTNTILP | 9 |
| 4 | 102 | 105 | PSHA | 4 |
| 5 | 113 | 133 | DVTTEDGRAVAHGNPMQEFPH | 21 |
| 6 | 136 | 137 | FH | 2 |
| 7 | 151 | 161 | PTHAGMQNQNF | 11 |
| 8 | 174 | 185 | DAQPEGSKNPSF | 12 |
| 9 | 196 | 202 | AFPRAGP | 7 |
Table 1: Bepipred linear epitope predicted epitopes
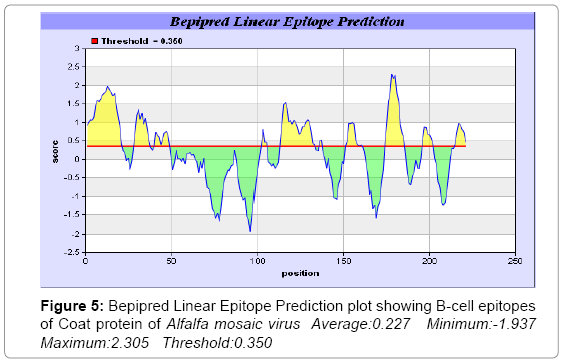
Figure 5: Bepipred Linear Epitope Prediction plot showing B-cell epitopes of Coat protein of Alfalfa mosaic virus Average:0.227 Minimum:-1.937 Maximum:2.305 Threshold:0.350
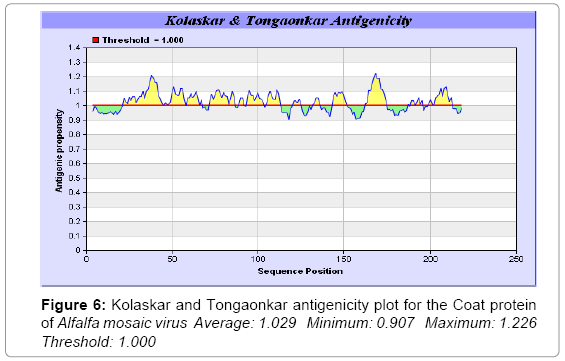
Figure 6: Kolaskar and Tongaonkar antigenicity plot for the Coat protein of Alfalfa mosaic virus Average: 1.029 Minimum: 0.907 Maximum: 1.226 Threshold: 1.000
| No. | Start Position | End Position | Peptide | Peptide Length |
|---|---|---|---|---|
| 1 | 22 | 44 | YAALRKAQLPKPPALKVPVVKPT | 23 |
| 2 | 46 | 67 | TILPQTGCVWQSLGTPLSLSSF | 22 |
| 3 | 72 | 86 | ARFLYSFLKDFVGPR | 15 |
| 4 | 94 | 103 | YRMVFSITPS | 10 |
| 5 | 105 | 113 | AGTFCLTDD | 9 |
| 6 | 144 | 151 | FELVFTAP | 8 |
| 7 | 162 | 174 | KHSYAVALCLDFD | 13 |
| 8 | 189 | 195 | EVWVERK | 7 |
| 9 | 199 | 212 | RAGPLRSLITVGLF | 14 |
Table 2: Kolaskar and Tongaonkar antigenicity Predicted peptides
Solvent accessible regions
We also predict solvent accessible regions in proteins; different measurement was performed for the prediction of antigenic activity, surface region of peptides. Emini et al. [44] (Figure 7,Table 3) predicts the highest probability i.e. found 14-KPTKRS-19, Score: 5.164 (maximum) (Average: 1.000 Minimum: 0.061 Maximum: 5.164 Threshold: 1.000), that a given protein region lies on the surface of a protein and are used to identify antigenic determinants on the surface of proteins. Karplus and Schulz (Figure 8) high score is found i.e. 2-SSSQKKA-8, Score : 1.127 (maximum) Average: 1.000 Minimum: 0.898 Maximum: 1.127 Threshold: 1.000. The Chou and Fasman scale which is commonly used to predict beta turns and position is i.e. 177-PEGSKNP-183, Score :1.334 (maximum). Predict backbone or chain flexibility on the basis of the known temperature B factors of the a-carbons. The hydrophobicity and hydrophilic characteristics of amino acids is determined by using different scales that are rich in charged and polar residues i.e. Sweet et al., Kyte & Doolittle, Abraham & Leo, Bull and Breese, Guy, Miyazawa et al., Roseman, Wolfenden et al., Wilson et al., Cowan, Chothia, Chou- Fasman, Manavalan et al., [46-59] shows hydrophobicity prediction result data found high in position (Figure 8-21).
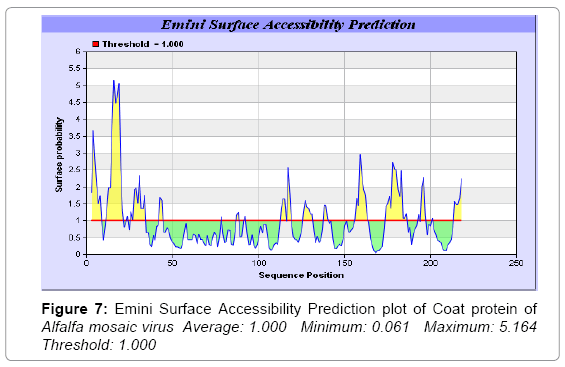
Figure 7: Emini Surface Accessibility Prediction plot of Coat protein of Alfalfa mosaic virus Average: 1.000 Minimum: 0.061 Maximum: 5.164 Threshold: 1.000
| No. | Start Position | End Position | Peptide | Peptide Length |
|---|---|---|---|---|
| 1 | 3 | 8 | SSQKKA | 6 |
| 2 | 12 | 21 | AGKPTKRSQN | 10 |
| 3 | 26 | 34 | RKAQLPKPP | 9 |
| 4 | 126 | 131 | NPMQEF | 6 |
| 5 | 156 | 163 | MQNQNFKH | 8 |
| 6 | 174 | 186 | DAQPEGSKNPSFR | 13 |
Table 3: Emini surface accessibility predicted peptides
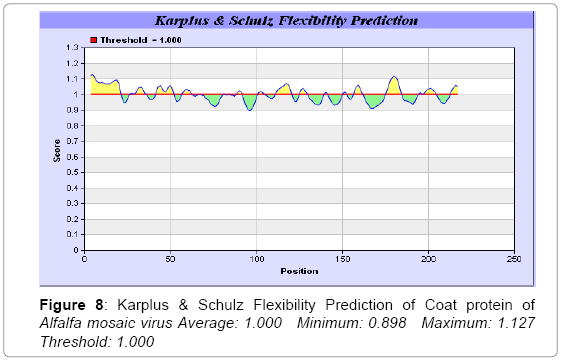
Figure 8: Karplus & Schulz Flexibility Prediction of Coat protein of Alfalfa mosaic virus Average: 1.000 Minimum: 0.898 Maximum: 1.127 Threshold: 1.000
Figure 9: Hydrophobicity plot of Sweet et al. (1983) of Coat protein of Alfalfa mosaic virus
Figure 10: Kyte & Doolittle hydrophobicity plot of Coat protein of Alfalfa mosaic virus
Figure 11: Abraham & Leo hydrophobicity plot of Coat protein of Alfalfa mosaic virus
Figure 12: Bull & Breese use surface tension to measure hydrophobicity and also uses negative values to describe the hydrophobicity of Coat protein of Alfalfa mosaic virus
Figure 13: Hydrophobicity plot of Miyazawa et al. (1985) of Coat protein of Alfalfa mosaic virus
Figure 14: Hydrophobicity plot of Guy (1988) of Coat protein of Alfalfa mosaic virus
Figure 15: Hydrophobicity plot of Wolfenden et al.(1981) of Coat protein of Alfalfa mosaic virus
Figure 16: Hydrophobicity plot of Roseman M.A.. (1988) of Coat protein of Alfalfa mosaic virus
Figure 17: Hydrophobicity/HPLC plot of Wilson & al (1981) of Coat protein of Alfalfa mosaic virus
Figure 18: Hydrophobicity/HPLC pH 3.4/ plot of Cowan (1990) of Coat protein of Alfalfa mosaic virus
Figure 19: Hydrophobicity plot of Chothia (1976) of Coat protein of Alfalfa mosaic virus
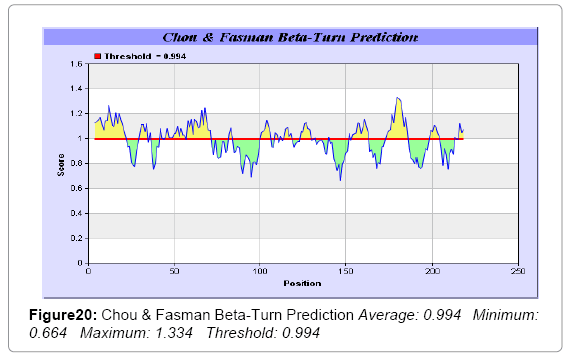
Figure 20: Chou & Fasman Beta-Turn Prediction Average: 0.994 Minimum: 0.664 Maximum: 1.334 Threshold: 0.994
Figure 21:Hydrophobicity plot of Manavalan et al of Coat protein of Alfalfa mosaic virus
Prediction of MHC binding peptide
We found binding of peptides to a number of different alleles using position specific scoring matrix. Coat protein of Alfalfa mosaic virus sequence is 221 residues long, having 8mer_H2_Db, 9mer_H2_Db, 10mer_H2_Db, 11mer_H2_Db MHC I binders. MHC molecules are cell surface proteins, which actively participate in host immune reactions and involvement of MHC-I and MHC-II in response to almost all antigens. We have predicted MHC-I peptide binders of coat protein of Alfalfa mosaic viruswas tested with on a set of 3 different alleles i.e. H2-Db (mouse) 8mer, H2-Db (mouse) 9mer, H2-Db (mouse) 10mer (Tables 4-7) and MHC-II peptide binders for I_Ab.p, I_Ad.p alleles highlighted in red represent predicted binders (Table 8). Here RANKPEP report PSSM-specific binding threshold and is obtained by scoring all the antigenic peptide sequences included in the alignment from which a profile is derived, and is defined as the score value that includes 85% of the peptides within the set [60-62]. Peptides whose score is above the binding threshold will appear highlighted in red and peptides produced by the cleavage prediction model are highlighted in violet. We also use a cascade SVM based TAPPred method which found 8 High affinity TAP Transporter peptide regions which represents predicted TAP binders residues which occur at N and C termini from coat protein of Alfalfa mosaic virus (Table 9).
| MHC:I Allele Mouse | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|
| 8mer_H2_Db | 183 | SKN | PSFRFNEV | WVE | 977.1 | 15.274 | 29.10% |
| 8mer_H2_Db | 48 | NTI | LPQTGCVW | QSL | 862.04 | 12.589 | 23.98% |
| 8mer_H2_Db | 115 | DDV | TTEDGRAV | AHG | 829.86 | 7.115 | 13.55% |
| 8mer_H2_Db | 15 | AGK | PTKRSQNY | AAL | 975.07 | 6.269 | 11.94% |
| 8mer_H2_Db | 180 | PEG | SKNPSFRF | NEV | 964.1 | 6.076 | 11.57% |
| 8mer_H2_Db | 202 | RAG | PLRSLITV | GLF | 880.1 | 5.516 | 10.51% |
| 8mer_H2_Db | 124 | AVA | HGNPMQEF | PHG | 941.03 | 4.134 | 7.88% |
| 8mer_H2_Db | 57 | VWQ | SLGTPLSL | SSF | 768.91 | 3.424 | 6.52% |
| 8mer_H2_Db | 34 | PKP | PALKVPVV | KPT | 804.04 | 2.857 | 5.44% |
| 8mer_H2_Db | 18 | PTK | RSQNYAAL | RKA | 904 | 2.673 | 5.09% |
| 8mer_H2_Db | 103 | ITP | SHAGTFCL | TDD | 816.93 | 2.282 | 4.35% |
| 8mer_H2_Db | 139 | FHA | NEKFGFEL | VFT | 965.08 | 1.594 | 3.04% |
| 8mer_H2_Db | 166 | HSY | AVALCLDF | DAQ | 833.02 | 1.347 | 2.57% |
| 8mer_H2_Db | 69 | SFN | GLGARFLY | SFL | 878.05 | 1.216 | 2.32% |
| 8mer_H2_Db | 10 | KAG | GKAGKPTK | RSQ | 767.91 | 1.015 | 1.93% |
| 8mer_H2_Db | 199 | AFP | RAGPLRSL | ITV | 851.03 | 0.795 | 1.51% |
| 8mer_H2_Db | 29 | RKA | QLPKPPAL | KVP | 845.06 | 0.452 | 0.86% |
Table 4: Promiscuous 8mer_H2_Db (Mouse) MHC I ligands, having C-terminal ends are proteosomal cleavage sites of coat protein of Alfalfa mosaic virus
| MHC:I Allele Mouse | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|
| 9mer_H2_Db | 17 | KPT | KRSQNYAAL | RKA | 1032.17 | 10.272 | 20.40% |
| 9mer_H2_Db | 199 | AFP | RAGPLRSLI | TVG | 964.19 | 9.692 | 19.24% |
| 9mer_H2_Db | 102 | SIT | PSHAGTFCL | TDD | 914.05 | 7.3 | 14.49% |
| 9mer_H2_Db | 67 | LSS | FNGLGARFL | YSF | 976.15 | 6.405 | 12.72% |
| 9mer_H2_Db | 184 | KNP | SFRFNEVWV | ERK | 1142.32 | 3.937 | 7.82% |
| 9mer_H2_Db | 134 | FPH | GAFHANEKF | GFE | 1002.1 | 3.723 | 7.39% |
| 9mer_H2_Db | 28 | LRK | AQLPKPPAL | KVP | 916.14 | 3.591 | 7.13% |
| 9mer_H2_Db | 50 | ILP | QTGCVWQSL | GTP | 980.13 | 2.371 | 4.71% |
| 9mer_H2_Db | 123 | RAV | AHGNPMQEF | PHG | 1012.11 | 1.777 | 3.53% |
| 9mer_H2_Db | 71 | NGL | GARFLYSFL | KDF | 1055.26 | 1.467 | 2.91% |
| 9mer_H2_Db | 74 | GAR | FLYSFLKDF | VGP | 1161.38 | 1.307 | 2.60% |
| 9mer_H2_Db | 161 | NQN | FKHSYAVAL | CLD | 1017.2 | 0.639 | 1.27% |
Table 5: Promiscuous 9mer_H2_Db (Mouse) MHC I ligands, having C-terminal ends are proteosomal cleavage sites of coat protein of Alfalfa mosaic virus
| MHC:I Allele Mouse | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|
| 10mer_H2_Db | 27 | ALR | KAQLPKPPAL | KVP | 1044.31 | 16.664 | 28.31% |
| 10mer_H2_Db | 209 | LIT | VGLFDEADDL | DRH | 1075.15 | 12.405 | 21.08% |
| 10mer_H2_Db | 181 | EGS | KNPSFRFNEV | WVE | 1219.37 | 9.238 | 15.70% |
| 10mer_H2_Db | 162 | QNF | KHSYAVALCL | DFD | 1086.32 | 7.505 | 12.75% |
| 10mer_H2_Db | 45 | KPT | NTILPQTGCV | WQS | 1027.19 | 5.087 | 8.64% |
| 10mer_H2_Db | 156 | HAG | MQNQNFKHSY | AVA | 1278.4 | 3.317 | 5.64% |
| 10mer_H2_Db | 122 | GRA | VAHGNPMQEF | PHG | 1111.24 | 2.966 | 5.04% |
| 10mer_H2_Db | 17 | KPT | KRSQNYAALR | KAQ | 1188.36 | 2.696 | 4.58% |
| 10mer_H2_Db | 39 | LKV | PVVKPTNTIL | PQT | 1063.29 | 1.449 | 2.46% |
| 10mer_H2_Db | 9 | KKA | GGKAGKPTKR | SQN | 981.15 | 1.237 | 2.10% |
| 10mer_H2_Db | 198 | KAF | PRAGPLRSLI | TVG | 1061.31 | 1.04 | 1.77% |
| 10mer_H2_Db | 135 | PHG | AFHANEKFGF | ELV | 1149.28 | 0.791 | 1.34% |
| 10mer_H2_Db | 160 | QNQ | NFKHSYAVAL | CLD | 1131.3 | 0.78 | 1.33% |
| 10mer_H2_Db | 74 | GAR | FLYSFLKDFV | GPR | 1260.51 | 0.645 | 1.10% |
| 10mer_H2_Db | 101 | FSI | TPSHAGTFCL | TDD | 1015.15 | 0.442 | 0.75% |
| 10mer_H2_Db | 8 | QKK | AGGKAGKPTK | RSQ | 896.04 | 0.219 | 0.37% |
Table 6: Promiscuous 10mer_H2_Db (Mouse) MHC I ligands, having C:terminal ends are proteosomal cleavage sites of coat protein of Alfalfa mosaic virus
| MHC:I Allele Mouse | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|
| 11mer_H2_Db | 17 | KPT | KRSQNYAALRK | AQL | 1316.53 | 11.716 | 14.74% |
| 11mer_H2_Db | 100 | VFS | ITPSHAGTFCL | TDD | 1128.31 | 6.124 | 7.70% |
| 11mer_H2_Db | 64 | PLS | LSSFNGLGARF | LYS | 1150.31 | 4.796 | 6.03% |
| 11mer_H2_Db | 134 | FPH | GAFHANEKFGF | ELV | 1206.33 | 2.131 | 2.68% |
| 11mer_H2_Db | 69 | SFN | GLGARFLYSFL | KDF | 1225.47 | 0.713 | 0.90% |
| 11mer_H2_Db | 136 | HGA | FHANEKFGFEL | VFT | 1320.48 | 0.278 | 0.35% |
| 11mer_H2_Db | 161 | NQN | FKHSYAVALCL | DFD | 1233.5 | 0.067 | 0.08% |
Table 7: Promiscuous 11mer_H2_Db (Mouse) MHC I ligands, having C:terminal ends are proteosomal cleavage sites of coat protein of Alfalfa mosaic virus
| MHC:II Allele | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|
| MHC:II I_Ab | 94 | DLI | YRMVFSITP | SHA | 1095.33 | 13.779 | 38.67% |
| MHC:II I_Ab | 120 | EDG | RAVAHGNPM | QEF | 934.08 | 12.843 | 36.04% |
| MHC:II I_Ab | 148 | ELV | FTAPTHAGM | QNQ | 914.04 | 11.612 | 32.59% |
| MHC:II I_Ab | 197 | RKA | FPRAGPLRS | LIT | 982.17 | 10.747 | 30.16% |
| MHC:II I_Ab | 165 | KHS | YAVALCLDF | DAQ | 996.2 | 10.614 | 29.79% |
| MHC:II I_Ab | 174 | LDF | DAQPEGSKN | PSF | 926.94 | 10.54 | 29.58% |
| MHC:II I_Ab | 146 | GFE | LVFTAPTHA | GMQ | 938.09 | 9.898 | 27.78% |
| MHC:II I_Ab | 30 | KAQ | LPKPPALKV | PVV | 944.23 | 9.818 | 27.55% |
| MHC:II I_Ad | 118 | TTE | DGRAVAHGN | PMQ | 877.91 | 12.73 | 23.95% |
| MHC:II I_Ad | 96 | IYR | MVFSITPSH | AGT | 1000.18 | 10.768 | 20.26% |
| MHC:II I_Ad | 161 | NQN | FKHSYAVAL | CLD | 1017.2 | 10.357 | 19.49% |
| MHC:II I_Ad | 98 | RMV | FSITPSHAG | TFC | 897.99 | 9.948 | 18.72% |
| MHC:II I_Ad | 68 | SSF | NGLGARFLY | SFL | 992.15 | 8.966 | 16.87% |
Table 8: Prediction of I_Ab & I_Ad MHCII ligands all rows highlighted in red represent predicted binders.
| Peptide Rank | Start Position | Sequence | Score | Predicted Affinity |
|---|---|---|---|---|
| 1 | 72 | ARFLYSFLK | 8.259 | High |
| 2 | 92 | LIYRMVFSI | 7.788 | High |
| 3 | 123 | AHGNPMQEF | 7.263 | High |
| 4 | 159 | QNFKHSYAV | 6.942 | High |
| 5 | 179 | GSKNPSFRF | 6.507 | High |
| 6 | 203 | LRSLITVGL | 6.337 | High |
| 7 | 88 | LEEDLIYRM | 6.3 | High |
| 8 | 195 | KAFPRAGPL | 6.027 | High |
Table 9: cascade SVM based High affinity TAP Binders of coat protein of Alfalfa mosaic virus
In this study, we found the antigenic determinants by finding the area of greatest local hydrophilicity. Hopp and Woods hydrophobicity scale is used to identify of potentially antigenic sites in proteins. Hydrophilicity Prediction result data found high in amino acid position at 165-173 [MIN: -2.633, MAX: 1.767] in a protein this scale is basically a hydrophilic index where apolar residues have been assigned negative values. The Window size of 5-7 is good for finding hydrophilic regions, greater than 0 values are consider as hydrophilic which is consider as antigenic. Welling used information on the relative occurrence of amino acids in antigenic regions to make a scale which is useful for prediction of antigenic regions and the predicted result data found high in sequence position 68-70. Welling antigenicity plot gives value as the log of the quotient between percentage in a sample of known antigenic regions and percentage in average proteins and prediction result data found high in position 13-41[MIN: -1.248, MAX: 0.873]. We also study Hydrophobicity plot of HPLC / Parker Hydrophilicity prediction result data found 112-DDVTTED-118, Score: 6.357 (maximum) [Average: 1.376, Minimum: -4.571, Maximum: 6.357]. BepiPred predicts the location of linear B-cell epitopes at position 1-MSSSQKKAGGKAGKPTKRSQN-21; 151-PTHAGMQNQNF-161 (Maximum), [Average:0.227 Minimum:-1.937 Maximum:2.305 Threshold: 0.350]. There are 3 antigenic determinant sequences is found by Kolaskar and Tongaonkar antigenicity scales the results show highest pick at position 165-YAVALCL-171, Score :1.226 (maximum) [Average: 1.029 Minimum: 0.907 Maximum: 1.226 Threshold: 1.000]. Result of determined antigenic sites on proteins has revealed that the hydrophobic residues if they occur on the surface of a protein are more likely to be a part of antigenic sites. This method can predict antigenic determinants with about 75% accuracy and also gives the information of surface accessibility and flexibility. Further this region form beta sheet which show high antigenic response than helical region of this peptide and shows highly antigenicity.
We predict solvent accessibility by using Emini et al. [44] the result found the highest probability i.e. found 5.164 Maximum in 14-KPTKRS-19, that a given protein region lies on the surface of a protein and are used to identify antigenic determinants on the surface of proteins. This algorithm also used to identify the antigenic determinants on the surface of proteins and Karplus and Schulz predict backbone or chain flexibility on the basis of the known temperature B factors of the a-carbons here we found the result with high score is i.e. 1.127 maximum in 2-SSSQKKA-8.
We predicted solvent accessibility of coat protein of Alfalfa mosaic virus for delineating hydrophobic and hydrophilic characteristics of amino acids. Solvent accessibility used to identify active site of functionally important residues in membrane proteins. Solvent accessible surface areas and backbone angles are continuously varying because proteins can move freely in a three-dimensional space. The mobility of protein segments which are located on the surface of a protein due to an entropic energy potential and which seem to correlate well with known antigenic determinants. We also found the hydrophobicity prediction result data found high in position. These scales are a hydrophilic with a polar residues assigned negative value. Because the N- and C- terminal regions of proteins are usually solvent accessible and unstructured, antibodies against those regions recognize the antigenic protein. Gomase method, B-EpiPred Server, Hopp and Woods, Welling, Parker, Kolaskar and Tongaonkar antigenicity scales were designed to predict the locations of antigenic determinants in Alfalfa mosaic virus (coat protein). Coat protein shows beta sheets regions, which are high antigenic response than helical region of this peptide and shows highly antigenicity. We also found the Sweet hydrophobicity, Kyte & Doolittle hydrophobicity, Abraham & Leo , Bull & Breese hydrophobicity, Guy, Miyazawa hydrophobicity, Roseman hydrophobicity, Cowan HPLC pH7.5 hydrophobicity, Rose hydrophobicity, Eisenberg hydrophobicity, Manavalan hydrophobicity, Black hydrophobicity, Fauchere hydrophobicity, Janin hydrophobicity, Rao & Argos hydrophobicity, Wolfenden hydrophobicity, Wilson HPLC hydrophobicity, Cowan HPLC pH3.4, Tanford hydrophobicity, Rf mobility hydrophobicity and Chothia hydrophobicity scales, Theses scales are essentially a hydrophilic index, with a polar residues assigned negative values.
In this study, we found predicted MHC-I peptide binders of toxin protein for 11mer_H2_Db, 10mer_H2_Db, 9mer_H2_Db, 8mer_H2_ Db alleles and I_Ab,I_Ad, for MHC II allele was tasted. The predicted binding affinity is normalized by the 1% fractil. The MHC peptide binding is predicted using neural networks trained on C terminals of known epitopes. In analysis predicted MHC/peptide binding is a logtransformed value related to the IC50 values in nM units. These MHC binding peptides are sufficient for eliciting the desired immune response. Predicted MHC binding regions in an antigen sequence and there are directly associated with immune reactions, in analysis we found the MHCI and MHCII binding regions. We also use a cascade SVM based TAPPred method which found 8 High affinity TAP Transporter peptide regions which represents predicted TAP binders residues which occur at N and C termini from coat protein of Alfalfa mosaic virus. TAP is an important transporter that transports antigenic peptides. TAP binds and translocate selective antigenic peptides for binding to specific MHC molecules. The efficiency of TAP-mediated translocation of antigenic peptides is directly proportional to its TAP binding affinity. Thus, by understanding the nature of peptides, that bind to TAP with high affinity, is important steps in endogenous antigen processing. The correlation coefficient of 0.88 was obtained by using jackknife validation test. In this test, we found the MHCI and MHCII binding regions. Cell immune responses are derived by antigenic epitopes hence their identification is important for design synthetic peptide vaccine. Cell epitopes are recognized by MHCI molecules producing a strong defensive immune response against of coat protein of Alfalfa mosaic virus. Therefore, the prediction of peptide binding to MHCI molecules by appropriate processing of antigen peptides occurs by their binding to the relevant MHC molecules. Because, the C-terminus of MHCI-restricted epitopes results from cleavage by the proteasome and thus, proteasome specifity is important for determing cell epitopes. Consequently, RANKPEP also focus on the prediction of conserved epitopes. C-terminus of MHCI-restricted peptides is generated by the proteasome, and thus RANKPEP also determines whether the C-terminus of the predicted MHCI-peptide binders is the result of proteasomal cleavage. Moreover, these sequences are highlighted in purple in the output results. Proteasomal cleavage predictions are carried out using three optional models obatined applying statistical language models to a set of known epitopes restricted by human MHCI molecules as indicated here.
From the above result and discussion it is concluded that the ability of RANKPEP to predict MHC binding peptides, and thereby potential epitopes, antigenic peptide that binds to MHC molecule are antigenic that means hydrophilic in nature. This means the increase in affinity of MHC binding peptides may result in enhancement of immunogenicity of coat protein of Alfalfa mosaic virus. Hence synthetic peptides will helpful in the designing of peptide vaccine. This approach can help reduce the time and cost of experimentation for determining functional properties of coat protein of Alfalfa mosaic virus. Overall, the results are encouraging, both the ‘sites of action’ and ‘physiological functions’ can be predicted with very high accuracies helping minimize the number of validation experiments.
This method will be applicable in immunodiagnostics, vaccine design for understanding of autoimmune susceptibility. Coat protein of Alfalfa mosaic virus involved multiple antigenic components and useful to protect the host from the nucleocapsid. MHC molecules are cell surface proteins, which take active part in host immune reactions and involvement of MHC class in response to almost all antigens and it give effects on specific sites. Predicted MHC binding regions acts like red flags for antigen specific and generate immune response against the parent antigen. So a small fragment of antigen can induce immune response against whole antigen. The method integrates prediction of peptide MHC class binding; proteosomal C terminal cleavage and TAP transport efficiency. This theme is implemented in designing subunit and peptide vaccines.