Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2016) Volume 5, Issue 3
A synthetic analog of ceramide (AD2725) elevated ceramide levels in MDA-MB-435 breast cancer cells inducing apoptosis and resulted in cell death. In animal studies, using a model of xenograft breast cancer cells in nude mice, treatment with AD2725 resulted in a significant reduction of the tumor volume and weight, and significantly prolonged survival of the treated mice. The analog also significantly elevated INF gamma and IL-12 cytokine levels and reduced the level of IL-10 in the plasma. These data indicate that use of suitable synthetic analogs that most probably elevate the ceramide levels may have beneficial anti-cancer therapeutic effects.
<Keywords: Ceramide; Sphingolipids; Apoptosis; Cytokine; Breast cancer
Apoptosis is recognized as an important mechanism by which cytotoxic agents induce tumor cell death [1,2]. One mechanism is using ceramide, as an intracellular, pro-apoptotic signaling molecule [3-5]. It is now clear that ceramide plays a central role in both apoptotic and mitogenic pathways [6,7]. Thus, generation of ceramide in cells initiates apoptosis and cell death while reduction of intracellular ceramide or increasing sphingosine-1-phosphate levels leads to drug resistance [8-12]. Ceramide is often generated in response to chemotherapeutic agents or radiation via hydrolysis of sphingomyelin [11,13]. Although the mechanism remains unclear, ceramide causes a disruption of mitochondria, leading to cell death by apoptosis [11,14-17]. Ceramide also reduces the vascular network that feeds the tumor [18]. In this study we used a synthetic ceramide-analog AD2725 to investigate the effects of chemotherapy while achieving an elevation of ceramide in the tumor cells. This study demonstrates that using a non-natural ceramide-like analog caused an elevation of intracellular ceramide level in MDA-MB-435 breast cancer cells, leading to their death by apoptosis and significantly prolonged the survival of tumor bearing mice.
AD2646: (2R,3R)-2-(N-tetradecylamino)-3-(4-nitrophenyl)-1,3- propandiol; AD2673: (2R,3R)-2-amino-3-(4-tetradecanoylaminophenyl)- 1,3-propandiol, were synthesized by Dagan et al. [19] AD2725: (2S,3R)-2-(N-tetradecylamino)-3-phenyl-3-propanol, was synthesized as follows: One mille mole (2S,3R)-2-amino-3-phenyl-3-propanol was reacted with tetradecanal (1.2 mmol) in methanol/0.05 N acetic acid, 9:1 for 15 min and sodium cyanoborohydride (NaCNBH3, 2 mmol) was added in portions during 1 h. The mixture was stirred overnight at room temperature, evaporated to dryness and the residue dissolved in dichloromethane/methanol, 2:1. Dilute HCl was added and following overtaxing, the upper phase was removed. The lower phase was washed two more times with dilute HCl and dried by adding MgSO4. The solvent was collected, evaporated to dryness and the product was purified by column chromatography using increasing concentrations of methanol in dichloromethane. Product is 550 μmol (55% yields). AD2725 structure see Figure 1A.

Figure 1: Structures of AD2725, effects of the analogs AD2725, AD2646 and AD2673 on viability of MDA-MB-435cells and induction of cell apoptosis. A: The chemical structures of the non-natural ceramide analogs AD2725. B: Effects of AD2725 on viability of tumor cell MDA-MB-435. MDA-MB-435 Cells (10^ 5/well) were plated overnight in serum free medium, and then were incubated with increasing concentrations of AD AD2725, AD2646 and AD2673 for 24 hours. The cells were collected and the number of viable cells determined, cell damage/death was determined by trypan blue staining as described in Materials and Methods. C and D: Inductions of apoptosis were measured by Caspase 3 activity and DNA laddering. MDA-MB-435 cells were incubated for 3 hours with 20 µM of AD2725. The cells were collected and apoptosis was analyzed by quantification of Caspase 3 activity (C): MDA-MB-435cells were incubated with AD2646 or AD2725 for 24 hours with 20 µM of the analogs; DNA was extracted and treated as described in Materials and Methods (D): Lane1: Control without analog; Lane2: with AD2725 and Lane3: AD2646 treatment.
Reagents
All chemical and solvents used were of analytical grade. Chemicals were purchased from Sigma Chemicals (St Louis, MO, USA). Thin layer chromatography plates were purchased from Whatman (Clifton, NJ, USA). Culture media and supplements were purchased from Beth Haemek, Israel.
Cell line
Breast cancer MDA-MB-435 cells were cultured at 37ºC in an incubator with 5% CO2 in air in RPMI-1640 medium, supplemented with 10% fetal calf serum, 1% penicillin-streptomycin, 1% glutamate and 4.5 g/L glucose. Once the cells grew to about 80% confluence, the medium was removed, fresh medium was added and incubation was continued.
Mice
Female CD1 nude mice (5-6 weeks old) weighing 20 g were purchased from the Harlan Laboratories, Israel. The animals were fed Purina chow and acidified water (pH 2.7) ad libitum, and maintained in an SPF animal facility at 21ºC with a 12 h cycling of light. The animal studies were approved by The Ethics Committee of the Hebrew University-Hadassah School of Medicine.
Determining of cellular protein concentration and cell number
Cells were harvested, washed with PBS, trypsinized, suspended in PBS and sonicated for 10 seconds with a Microson XL probe sonicator (Misonix) at 40% output power. For quantifying the protein, a sample (5 μl) was taken, interacted with Bradford reagent (250 μl) and the absorbance quantified at 595 nm. A culture sample was mixed with an equal volume of trypan blue, applied to a hemocytometer, and the numbers of live (unstained) and damaged or dead (blue) cells were determined.
Cell viability analysis
104 cells per 100 μl medium were seeded in each well in a 96-well plate, incubated overnight and varying concentrations of ceramide analogs in 100 μl medium containing 2 μl DMSO were added to the cultures. (The controls contained the same concentration of DMSO). An MTT solution was added and incubation was continued for 4 hours. The stop-solution [89% Isopropanol, 1% HCl (37%), 10% TtritonX-100] was then added and cell viability was determined at 570 nm.
Apoptosis determined by DNA fragmentation
1 × 106 MDA-MB-435 cells in 5 ml medium were seeded in a small flask and incubated for 24 h. The next day the medium was removed, new medium and 20 μM of AD2725 or AD2646 were added and incubation was continued for 24 h. On the third day, cells were harvested by trypsinization and the genomic DNA was extracted using the Sigma DNA extraction kit. Samples of DNA were then electrophoresed on a 1.5% agarose gel at 50 volts in TAE electrophoresis buffer for 2 hours. The DNA was detected by a UV-illuminator and pictures were taken.
Apoptosis determined by measuring activation of Caspase 3
Caspase-3 activity was assayed by Z-DEVD-R110 cleavage, using the EnzChek™ Caspase-3 Assay Kit #2 (Molecular Probes). Following incubation with the respective analogs, cells were collected, precipitated and washed with PBS. For each sample, 106 cells were lysed by incubating with 50 μl lysis buffer on ice for 30 min and cell debris was precipitated in a micro-centrifuge for 5 min at 5,000 rpm. The supernatants were incubated with 50 μM of the conjugated substrate Z-DEVD-R110 in a 100 μl reaction volume at room temperature for 30 min and the fluorescence of the released free R110 was measured at an excitation at 485 nm and emission at 535 nm using a FL600 microplate fluorescence reader.
Quantification of cellular ceramide
MDA-MB-435 cells were incubated for 3 hours in the absence or presence of the ceramide analog AD2725 and AD2646 (20 μM) and 15 μM pyrene decanoic acid (P10), as a solution in 1 μl DMSO per ml reaction mixture. The cells were trypsinized, collected and washed twice with PBS. 2 ml PBS were added and the cell suspension was sonicated in a Microson XL probe sonicator (Misonix) for 10 seconds. A sample was taken for quantifying the cell protein using the Bradford procedure. To the rest were added 2 ml of dichloromethane-methanol, 1:1, stirred and centrifuged. The lower phase was collected and the upper one retreated with 1 ml of dichloromethane-methanol, 3:1. The two dichloromethane phases were combined and the solvent was evaporated to dryness under a stream of air. The residue was dissolved in dichloromethane-methanol, 1:1 and applied to the concentrating zone of a TLC silica gel plate having markers of P10 and P10-ceramide. The latter was developed in a mixture of dichloromethane-methanolammonia, 95:5:0.5. The P10-ceramide spots (viewed with an ultraviolet lamp) were scraped, dichloromethane-methanol, 1:1, was added, stirred sonicated in bath sonicator and centrifuged. The fluorescence of P10-ceremide was recorded in a spectrofluorometer (Perkin-Elmer LS5) using an excitation at 345 nm as an emission at 377 nm.
In vivo studies
CD1-Nude mice (5-6 week old) were irradiated with a total body irradiation (TBI) of 400 cGy and one day later injected with MDA-MB- 435 cells.
Four types of experiments were performed:
1) Breast cancer cells (4 × 106 cells in 0.1 ml medium) were injected intradermally into the left side of the flank and the next day the mice were injected i.p. with the analog AD2725.
2) Breast cancer cells (4 × 106 cells in 0.1 ml medium) were injected intradermally into the left side of the flank and one week later the mice were injected i.p. with the analog AD2725.
3) Breast cancer cells (5 × 106 cells in 0.2 ml medium) were injected intravenously into the lateral tail vein and two hours later the mice were injected i.p. with the analog AD2725.
4) Breast cancer cells (5 × 106 cells in 0.2 ml medium) were injected intravenously into the lateral tail vein and the mice were given the analog AD2725 in the drinking water, starting day 0.
The tumor size was measured twice per week and the tumor volume was calculated from the formula: length × width2 × π/6. Mice injected with saline or drunk regular water were used as controls. At the end of experiment, mice were sacrificed with CO2; the tumors on the skin were removed and weighed. All organs of the mice were tested for presence of metastases.
Cytokine assay
The levels of cytokines in the plasma were assayed by ELISA assay. ELISA reagents were purchased as Opt. EIA Cytokine ELISA sets from BD Biosciences (San Diego, CA, USA) and the respective cytokines were analyzed according to the manufacturer’s protocol as previously described [20].
Statistical analysis
Most results are based on experiments run in triplicates at least twice, except where indicated. Statistical analyses were performed by Student's t-test (except where indicated) and the results were considered statistically significant when p<0.05.
Cytotoxic effects of the sphingolipid analogs
MDA-MB-435 cells (0.1 × 106 cells/ml, 100μl/well) were grown overnight in RPMI-1640 without FCS in 96-well plates, then incubated for 24 hours with increasing concentrations of three ceramide analogs AD2646, AD2673 and AD2725. The viability of cells were determined by the MTT assay, all the synthetic drugs exhibited cytotoxicity toward breast cancer cell line MDA-MB-435 based on a dose respond manner (Figure 1B), the estimated IC50 of AD2725 was 12.5 μM. Trypan blue staining results further showed that all the 3 synthetic analogs of ceramide severely damaged the breast cancer cells. AD2725 as a ceramide analog was used for a further study in vivo (average of 2 experiments, each one in triplicates).
Induction of apoptosis by the analogs
Caspase-3 analysis: MDA-MB-435 cells (1 × 106 cells/ml) were incubated for 3 hours with 20 μM AD2725, the cells were collected and induction of apoptosis was analyzed by the Caspase-3 activity procedure. As shown in Figure 1C, AD-2725 increased the Caspase-3 activity 4-5 fold relative to the control in which the cells were grown without the analogs (Figure 1C).
DNA laddering: In a small flask 2 × 105 of MDA-MB-435 cells/ ml were seeded, 20 μM each of AD2725 and AD2646 were added and incubation for 24 hours. Cells were trypsinized and harvested; the genomic DNA was extracted, quantified and electrophoresed. Figure 1D shows the DNA electrophoresis on 1.5% agarose gel. DNA laddering was seen after 24 hours analogs treatment and without in control. In parallel experiments (not shown), the DNA laddering was not appeared after 6 hours when the concentration of the analogs was 20 μM, but when the concentration of AD2725 raised to 80 μM, considerable DNA fragments laddering was observed after 5 hours and was strengthened after 7 hours.
Quantification of cellular ceramide: As shown in HL-60 cell with ceramide analogs by Dagan et al. [19], incubation of MDA-MB-435 cells with AD2725 showed elevated cellular ceramide (Figure 2). This was measured by the conversion of pyrene-dodecanoic acid (P12) to pyrene dodecanoylsphingosine (P12-ceramide). In the current study, 6 × 106 MDA-MB-435 cells were incubated with 50 μM of P12 for 3 hours in the absence or presence of 20 μM of AD2646 or AD2725. The lipids were extracted, applied to a thin layer chromatography plate, and was developed in a mixture of chloroform–methanol–ammonia 95:5:0.5 (by volume). The P12-ceramide spots were visualized with a UV-lamp, and then were scraped, the lipid was extracted and the fluorescence was quantified in a spectrofluorometer., In Figure 2, the values of ceramides are presented as the fluorescent units divided by the cellular protein, indicating that AD2646 resulted in a 2.3 fold increase and AD2725 in a 2.6 fold P12-ceramide increased compared to the control.
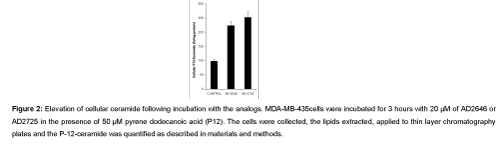
Figure 2: Elevation of cellular ceramide following incubation with the analogs.MDA-MB-435cells were incubated for 3 hours with 20 µM of AD2646 or AD2725 in the presence of 50 µM pyrene dodecanoic acid (P12). The cells were collected, the lipids extracted, applied to thin layer chromatography plates and the P-12-ceramide was quantified as described in materials and methods.
Effect of AD2725 on the growth of MDA-MB-435 Xenografts: CD1-Nude mice were irradiated with 400 cGy TBI and following one day, each mouse was injected i.d. into one flank with 4 × 106 MDA-MB- 435 cells in a volume of 0.1ml. After one day the mice were injected i.p. with 5 mg/kg of AD2725 in 10% CE saline daily. A control group (n=8) of mice was similarly treated with saline. As seen in (Figure 3A) after 25 days of daily injections, the tumors in AD2725 injected mice were significantly smaller than that of the controls. In another experiment the mice were injected i.d. with 4 × 106 MDA-MB-435 cells and when the tumors were palpated after a week, the ceramide analog AD2725 was injected i.p. for another 6 weeks. Also in these experiments there was a significant inhibition of the tumor growth, measured up to 25-50 days (Figure 3B).
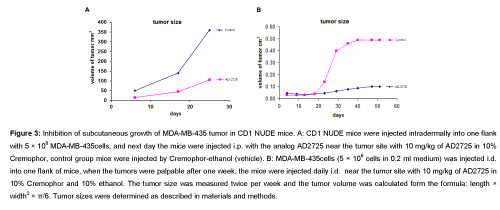
Figure 3: Inhibition of subcutaneous growth of MDA-MB-435 tumor in CD1 NUDE mice. A:CD1 NUDE mice were injected intradermally into one flank with 5 × 106 MDA-MB-435cells, and next day the mice were injected i.p. with the analog AD2725 near the tumor site with 10 mg/kg of AD2725 in 10% Cremophor, control group mice were injected by Cremophor-ethanol (vehicle). B:MDA-MB-435cells (5 × 106 cells in 0.2ml medium) was injected i.d. into one flank of mice, when the tumors were palpable after one week; the mice were injected daily i.d. near the tumor site with 10 mg/kg of AD2725 in 10% Cremophor and 10% ethanol. The tumor size was measured twice per week and the tumor volume was calculated form the formula: length × width2×π/6.Tumor sizes were determined as described in materials and methods.
Survival of nude mice injected i.v. with human breast cancer cells and treated with AD2725: CD1-Nude mice, 6 mice in each group, were irradiated with 400 cGy, and one day latter injected intravenously with 5 × 106 MDA-MB-435 cells and AD2725 (10 mg/kg) was administrated by intraperitoneal injections after two hours, and then continue 60 injections, starting day 0, 6 days per week. Figure 4A shows that all the treated mice by AD2725 survived till 80 days while the control mice started to die on 43 days and 2 mice of 6 died till 80 days. In another experiment, 8 mice in each group were injected with 5 × 106 breast cancer cells i.v. and the ceramide analog AD2725 was administered in the drinking water (10 mg/kg, starting day 0). The mice were followed up for 100 days. Figure 4B shows that mice treated with AD2725 in daily drink did not lose weight and survived more than 100 days. On the other hand 5 of the 8 mice in the control group died (Figure 4B).
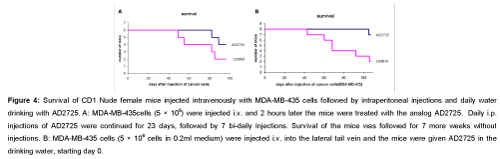
Figure 4: Survival of CD1 Nude female mice injected intravenously with MDA-MB- 435 cells followed by intraperitoneal injections and daily water drinking with AD2725. A: MDA-MB-435cells (5 × 106) were injected i.v. and 2 hours later the mice were treated with the analog AD2725. Daily i.p. injections of AD2725 were continued for 23 days, followed by 7 bi-daily injections. Survival of the mice was followed for 7 more weeks without injections. B: MDA-MB-435 cells (5 × 106 cells in 0.2ml medium) were injected i.v. into the lateral tail vein and the mice were given AD2725 in the drinking water, starting day 0.
Ceramide analog AD2725 effect on cytokine profile: Cytokine concentrations of IL-12, IFN-γ and IL10 were assessed by ELISA in sera from each group of mice. A significant reduction of IL10 was seen in AD2725 treated mice compared to the vehicle control (p<0.05). IFN- and IL-12 were significantly elevated in AD2725 treated mice compared to control (p<0.05) (Table 1).
| In vivo treatment of Mice | Number of Mice | Cytokine evaluated | pg/ml Median | pg/ml Range | |
|---|---|---|---|---|---|
| AD2725 | 4 | Plasma | IL-10 | 10 | 10-660 |
| Vehicle | 4 | Plasma | IL-10 | 372 | 19-820 |
| AD2725 | 4 | Plasma | IL-12 | 217 | 122-3800 |
| Vehicle | 4 | Plasma | IL-12 | 15 | 10-1080 |
| AD2725 | 6 | Plasma | IF N gamma | 115 | 10-240 |
| Vehicle | 7 | Plasma | IF N gamma | 10 | 10-185 |
Table 1: Cytokine levels in the plasma from untreated and AD2725 treated mice.
The accumulation of intracellular ceramide as a result of a) treatment by ceramide analogs in vitro [21], b) modification of endogenous ceramide metabolism [22], c) inhibition of the conversion of ceramide [23] and d) the induction of de novo ceramide synthesis [24] cause cancer cell death through apoptosis. Our results and those of others illuminate the potential therapeutic utility of ceramide to cancer. In this study, we show that the application of an analog of ceramide AD2725 has a cytotoxic effect on MDA-MB-435 breast cancer cells and 5 μM of AD2725 may reduce the number of cancer cells. These results are in agreement with other previous studies showing that ceramide can effectively induce apoptosis in tumor cells [25,26]. Moreover, in this study we show that AD2725 exhibited mitochondrial toxicity (IC50 12.5 μM) leading to cell death, we proposed that the mechanism of cell death is because of the accumulation of ceramide in mitochondria. The mechanism of ceramide induced apoptosis has been well established, the elevated ceramide in mitochondria is responsible for mitochondria dysfunction including loss of electron potential, cytochrome C release and prerequisites for caspase activation [27,28].
In agreement with these data, our results show a significant inhibition of growth of the human breast cancer cells MDA-MB-435 in vivo as xenograft in nude mice. The mice that were injected i.v. with tumor cells and drank water with AD2725 did not lose weight and survived more than 100 days, whereas 60% of the control died within this period. Same as our previously study in TSU-Pr1 prostate cancer model [29], we found mice injected i.v. with MDA-MB-435 cells and treated i.p. with AD2725 showed a remarkable reduction of metastases in internal organs of the survivors, whereas all control mice died with developed metastases tumors (data not show). Measuring the cytokine levels in the plasma revealed a significant reduction in IL-10 level in the AD2725 treated mice and a huge elevation in IL-12 and IFN-γ. IFN-γ plays a crucial role in enhancing tumor cell cytotoxicity and this has been showed in T cells mediated breast cancer (MCF-7) cells killing [30]. Preclinical studies demonstrat that IL-12 promotes specific anti-tumor immunity in several types of tumors [30]. Those results that IL-12 induced production of IFN-γ and suppression of IL-10 are in agreement with ours. Ceramide down-regulates LPS-stimulated production of IL-5, IL-10, and IL-13 [31]; the pro-inflammatory cytokine (eg. TNF--∝) mediated synthesis of ceramide can be inhibited by IL-10 and IL-13 through the activation of PI3-kinase [29]. Elevating Th1 related cytokines and reducing Th2 related IL-10 is responsible for eradication of tumor cells, vaccination with breast cancer cells that were transfected by pcDNA-IL-12 induced a Th1 dominant immune response and resulted in IL-12 secretion and tumor regression in mice [30]. Eguchi et al. [32] suggested that IL-12 stimulates non-specific killing, they showed that transduction of the IL-12 gene to MC38 significantly reduced tumorigenicity in immunocompetent mice. Lee et al. [33] showed that herbal prescription also significantly inhibited liver tumor metastasis and caused a marked increase of Th1 cytokine (IFN-γ) and a decrease of Th2 cytokine (IL-4) stimulated by Concanavalin A. Tsung et al. [34] demonstrated that IL-12 played an essential role in the induction of Th1 response against tumors and showed that IL-12 caused complete regression of 10-days s.c. tumors (4-8 mm) with activation of NO synthase and induction of macrophages. In addition, Park et al. and Cohen et al. [35,36] summarized that healthy women with a family history of breast cancer have shown decreased immune responses, such as low NK cell activity and low Th1-cytokine production.
In light of these results, we suggest that administration of ceramide analog AD2725 elevates ceramide in MDA-MB-435 breast cancer cells leading to apoptosis and growth inhibition of tumor. In addition, we propose that ceramide analogs AD-2725 can react as immune modulators and create a shift of Th2 to Th1. Th1 cells and Th1 associated cytokines can create anti-tumor response which leads to a suppression of tumor growth and induces cell death.
The authors report no conflict of interest.