Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2013) Volume 1, Issue 3
The study of food webs is a very important topic in order to understand the dynamics and stability of communities. We studied the food webs structure and energy balance of some tide pools from a rocky shore of Mar del Plata (Argentina). Food webs were drawn, and connectivity, proportion of different trophic species, and the link-richness regression slope were determined. Photosynthesis and respiration rates of several species were also measured. All the topological measures were calculated with and without sediments/detritus as potential energy sources. The proportion of basal species and the connectivity were greater when detritus were included; proportion of intermediate species lowed significantly with detritus inclusion. There were no differences between seasons, but sampling sites did show differences. Proportion of top species and connectivity did not show differences in any case. Some discrepancies were found with respect to the theoretical expected values of the food web properties: the proportion of basal species was significantly higher and the intermediate species proportion significantly smaller than the theoretical ones. Slope of the L-S regression was near 2 and the fit was improved when detritus was included. Mesograzers showed higher respiratory rates in spring and green algae higher photosynthetic rates in autumn. Considering the entire intertidal zone, anemone and mussels were the most important oxygen consumers. The intertidal zone works as a source of production with P/R ratio greater than one.
<Keywords: Intertidal community; Food web; Connectivity; Detritus; Sediments
The structure of food webs not only describes the feeding relationships between organisms in a given community; moreover, it can reveal dynamic properties of the community like resilience, nutrient recycling and resistance. Two main approaches have been used in field studies of food webs: the topological approach and the energetic one. The first refers to the topological regularities in the food web [1-5] and involves some statistical “laws” or invariants [1,6]. The second approach considers energy fluxes and dissipation at each node of the web [7].
Regularities in food webs topology constitute the basement of the classic “food web theory”. Many authors have critiqued these theories arguing that such generalizations probably are artifacts due to the poor quality of the data sets. Most analyses showed that properties varied with scale, the scale dependent hypothesis being more successfully predictive and precise than the older paradigm of scale invariance [8-12].
Topological and energetic analyses, help ecologists to understand some properties of communities and ecosystems; for instance, it is known that interaction strength seems to be related to web stability and connectivity [5,13,14]. May [13] has proved that highly connected or poorly connected webs are not globally stable, and there is a rank of intermediate connectivity inside which food webs are persistent.
In general, detritus and sediments were not considered as equivalent to trophic species in the pioneer works [15]; however, Pimm [16] have included detritus as one “species” and classified food webs as “Lotka- Volterra systems” when basal species are plants, and “donor-controlled systems” when detritus and sediments are the base of webs. In such donor-controlled systems, the control of population densities is from the type bottom-up.
Food webs on marine littoral shores can be considered as a mix of both types of systems because they have primary producers (macroalgae) and organic detritus that maintain a rich community of consumers [17], particularly in tide pools. Several organisms colonize these pools and interact with each other in a complex way [18]. Most of them are benthic sessile species, although occasional mobile species become trapped or seek refuge inside the pools at low tides; these organisms range from plankton to larger forms like nektonic macroscopic mollusks and crustaceans [17-19].
In Argentina, rocky shores have been studied showing some predator-prey relationships [20-22] and the effects of physical conditions [22-23] on individual species and community structure. Tide pools have been little studied, although they are of great ecological interest to develop scientific research on trophic dynamics. We describe the trophic structure of tide pool in the framework of food webs theory and report the metabolic rates of some species in these systems, analyzing the relationship between the structure of webs and energy fluxes in relation to explanatory environmental variables. We tested whether the inclusion of detritus as a trophic species increased the connectivity values and allowed us to detect new competitive and regulatory relationships.
Study sites
The study was carried out on the Mar del Plata coastline (38°02´S, 57°31´O, Figure 1). In this region, the intertidal zone is mainly composed of soft sediments (mainly sand), however, there are occasional rocky outcrops of different magnitude. There are tide pools with diverse morphological characteristics (shape, depth and volume) and located at different heights across the intertidal zone.
Figure 1: Location of the study sites Cabo Corrientes and Playa Chica in intertidal rocky shores in Mar del Plata (Argentina).
Tides are semidiurnal with amplitudes not exceeding 1 m. Shore water temperature varies between 9°C and 20°C; salinity between 32.6 and 34.6% [23].
Data collection and analysis
Two sample sites were chosen (Figure 1): Cabo Corrientes (CC) and Playa Chica (PC). Three representative pools were sampled at Cabo Corrientes (CC1, CC2 and CC3) and two at Playa Chica (PC1 and PC2).
Samples were taken on unclouded days at low tide level periods, two times in the year: Autumn (May, 2007) and Spring (November, 2007). Species were surveyed and samples of algae and their associated fauna were taken from each tide pool. Samples were analyzed under stereoscopic microscope and all organisms were identified to the finest possible taxonomic level. The cover of species was quantified by means of a 1600 cm2 grid divided in 4 cm2 cells.
Morphology of pools was characterized by the measurement of the horizontal minimum and maximum diameter and the maximum depth. Using these data, the floor surface and the volume of each pool were computed using a spherical cap approximation. In the case of very irregular pools, we used an average of several diameters. The pond volumes varied between 20 to 424 liters.
Food webs were drawn for each tide pool combining published information about the organisms’ eating habits and diets with field observations [24-31]. Phytoplankton and zooplankton (not sampled in this work) were included in food webs in order to avoid underestimation of connectivity. From the drawn webs, the following parameters were calculated: connectivity [15] as C=Creal/Cpossible, where Cpossible=S(S-1)/2, ratios of basal (B), intermediate (I) and top (T) species, slope of the number of observed links (L) versus richness (S).
All calculations were made in two ways: including and excluding detritus/sediment as basal “species”. This method allowed us to evaluate the properties proposed by Pimm [16] for donor-controlled systems.
Connectivities and ratios of B, I and T species were compared using non parametric two-way Kruskal-Wallis test [32] using as factors the sampling season (Autumn/Spring) and the inclusion or not of the sediments and detritus as a trophic species (Yes S-D/No S-D).
Metabolic measures
Photosynthetic and respiratory rates were measured by closed respirometry [33]. Organisms were incubated for one hour in closed bottles of size according to the body size of each species and changes in oxygen concentrations were measured using a Hanna HI 9142 device (rank 0.0 to 19.9 mg/l; accuracy ±0.1 mg/l). In the case of algae, two successive measures were made: one on dark (only respiration) and one on light (photosynthesis and respiration) in order to calculate the gross photosynthesis. In laboratory, individuals were dried until constant weight in order to calculate the specific metabolic rates (per unit of biomass). Using cover and abundance information and the floor surface of each pool, respiration and photosynthesis per square meter were calculated for the whole intertidal zone.
For all the sessile species sampled, cover area (cm2) and percentage of cover were measured in Autumn and Spring. These data were further used to calculate the contribution of each species to the total metabolism of the intertidal zone. The cover dominance varies between green algae, red algae and mussels depending on the wave conditions and the location of the pool across the lower-higher tide axis. The main algae were Corallina officinalis, Polysiphonia fucoides, Chaetomorpha aerea and Ulva rigida. Only in Spring, was a high cover of Enteromorpha sp. found.
The mussels Brachidontes rodriguezii and Mytilus sp. were the main sessile invertebrate species found.
Richness and food web topology
Of the 24 taxa found at the studied tide pools, ten were classified as basal species (producers), ten were intermediate species (mainly crustaceans and mollusks), and four were top species (mainly anemones and polychaetes).
Food webs were compiled for the five tide pools at the two sampling periods (Figure 2) including the 24 taxa plus sediment and organic detritus as trophic species (nodes). Based on these drawings, the food webs parameters were calculated and compared (Figure 3); Kruskal- Wallis test showed that proportions of B and I species, and connectivity were significantly different depending on the inclusion of detritus or not (p<0.05). Connectivity and proportion of B species increased and proportion of I species decreased when detritus was included. There were no significant differences between seasons for any topological parameter.
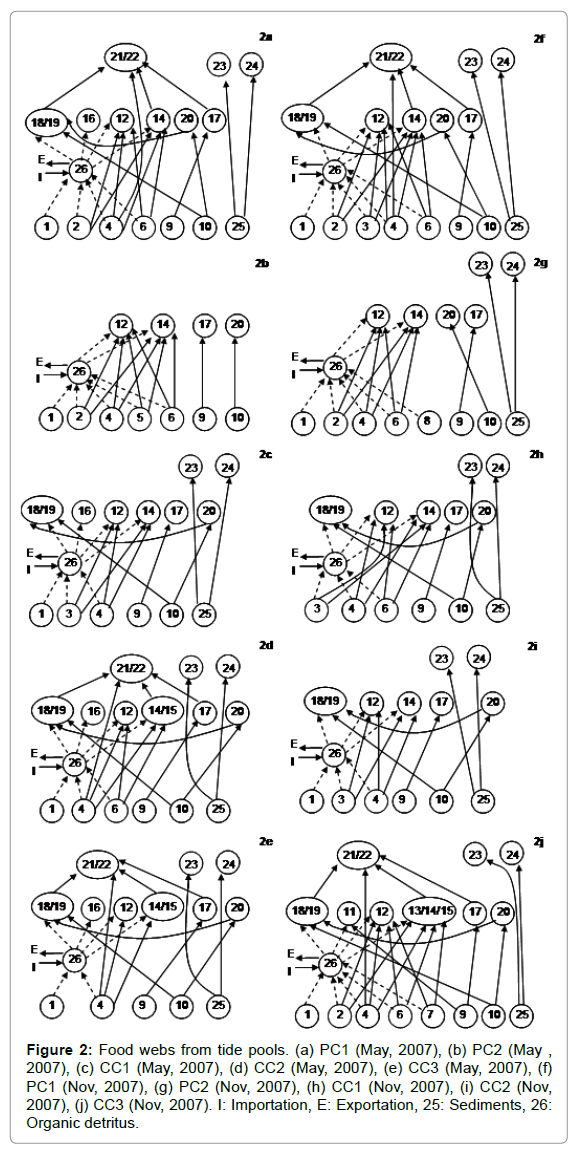
Figure 2: Food webs from tide pools. (a) PC1 (May, 2007), (b) PC2 (May , 2007), (c) CC1 (May, 2007), (d) CC2 (May, 2007), (e) CC3 (May, 2007), (f) PC1 (Nov, 2007), (g) PC2 (Nov, 2007), (h) CC1 (Nov, 2007), (i) CC2 (Nov, 2007), (j) CC3 (Nov, 2007). I: Importation, E: Exportation, 25: Sediments, 26: Organic detritus.
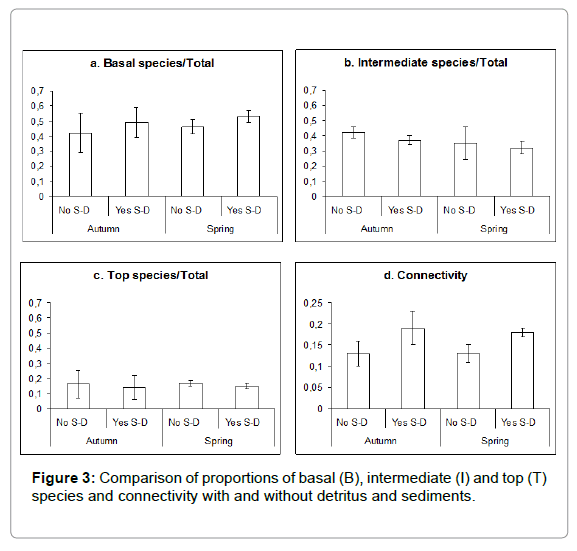
Figure 3: Comparison of proportions of basal (B), intermediate (I) and top (T) species and connectivity with and without detritus and sediments.
Proportions of B and I species, considering the ten webs, were tested for fit to a Normal distribution (Kolmogorov-Smirnov test); this allowed comparison of the mean values with the predicted by the food web theory (using the 95% confidence interval). The proportion of B species (mean 0.47) was significantly greater than the theoretically expected value (0.20); moreover, the mean proportion of I species (0.37) was significantly lower than the theoretically expected value (0.50).
Another quantitative descriptor of food webs is the link density [34], that is, the relationship between the number of links and the number of nodes; Figure 4 shows these relationships with and without the inclusion of detritus and sediments. Including sediment and detritus, raised the slope upwards from 1.78 to 2.11, nearest to the theoretical value, and improved fit.
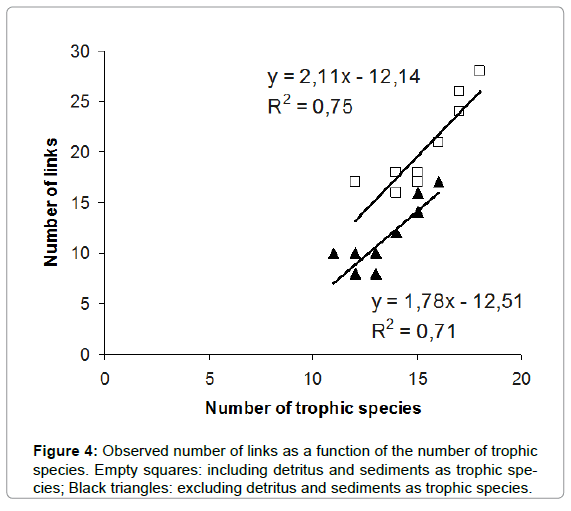
Figure 4: Observed number of links as a function of the number of trophic species. Empty squares: including detritus and sediments as trophic species; Black triangles: excluding detritus and sediments as trophic species.
Metabolic rates
Amphipods and isopods have the highest specific respiratory rates, mainly in spring, whereas the green algae Ulva rigida and Chaetomorpha aerea and the red alga Polysiphonia fucoides have the highest photosynthetic rates, mainly in autumn (Table 1). However, when we consider the abundance of each organism, the values of metabolism projected for pool in the entire intertidal zone are different (Table 2). On a while shore scale, the red alga Corallina officinalis is the most important primary producer, followed by Ulva rigida;and Polysiphonia fucoides; and anemone and mussels are the most important organisms in terms of oxygen consumption in tide pools.
| Organisms | RR (mgC × h-1 × gDW-1) | GPR (mgC × h-1 × gDW-1) | NPR (mgC × h-1 × gDW-1) | |||
|---|---|---|---|---|---|---|
| Autumn | Spring | Autumn | Spring | Autumn | Spring | |
| Corallina officinalis | 0.02 | 0.04 | 0.29 | 0.22 | 0.27 | 0.18 |
| Polysiphonia fucoides | 0* | 0.12 | 0.76 | 0.50 | 0.76 | 0.38 |
| Enteromorpha sp. | - | 0.04 | - | 0.14 | - | 0.10 |
| Ulva rigida | 0* | 0.05 | 3.24 | 0.16 | 3.24 | 0.11 |
| Chaetomorpha aerea | 0* | 0.10 | 3.24 | 0.12 | 3.24 | 0.02 |
| Cladophora sp. | - | 0.04 | - | 0.77 | - | 0.73 |
| Dictyota sp. | - | 0.22 | - | 0.41 | - | 0.19 |
| Idotea baltica (I) ** | 1.19 | - | ||||
| Sphaeroma serratum (I) | - | 4.21 | ||||
| Hyale grandicornis (I) | 1.91 | 3.45 | ||||
| Siphonaria lessoni (I) | 0.03 | 0.37 | ||||
| Mussels*** (I) | 0.06 | 0.09 | ||||
| Bunodosoma zamponi (T) | - | 0.59 | ||||
| Aulactinia sp. (T) | 0.03 | - | ||||
Table 1: Specific metabolic rates (per unit of biomass). RR: Respiration Rate, NPR: Net Photosynthetic Rate, GPR: Gross Photosynthetic Rate.
Although metabolic measurements were no made of all species present, we can say that roughly the intertidal zone acts as a producer system in spring and autumn because P/R values are very higher than one (Table 2). Notably, the value of P/R ratio is higher in autumn (14.3) than spring (3.4), probably because, in spring, the temperature affects the respiration rates more than the photosynthetic ones, producing lower ratios. However, on a daily basis, considering the photoperiod, values could be similar by the increment of daylight hours in November respect to May.
| Organisms | RR (mgC × h-1 × m-2) | GPR (mgC × h-1 × m-2) | NPR (mgC × h-1 × m-2) | |||
|---|---|---|---|---|---|---|
| Autumn | Spring | Autumn | Spring | Autumn | Spring | |
| Corallina officinalis | 6.92 | 7.41 | 99.08 | 93.30 | 92.16 | 34.99 |
| Polysiphonia fucoides | 0* | 5.28 | 40.32 | 44.62 | 40.32 | 16.73 |
| Enteromorpha sp. | 6.89 | 43.57 | 16.34 | |||
| Ulva rigida | 0* | 13.35 | 83.08† | 84.43 | 83.08† | 31.66 |
| Chaetomorpha aerea | 0* | 9.58 | 4.17 | 1.56 | ||
| Cladophora sp. | 1.48 | 64.90 | 24.34 | |||
| Mussels** (I) | 9.46 | 29.27 | ||||
| Family Actiniidae(T) | 110.80 | 543.24 | ||||
Table 2: Metabolic rates per m2, projected to the total intertidal region. RR: Respiratory Rate, GPR: Gross Photosynthetic Rate, NPR: Net Photosynthetic Rate.
Comparing with the data reported by Cohen [15], the webs of tide pools have a higher proportion of basal species and a lower proportion of intermediate species. Intermediate and top species are mobile and may be reduced by wave disturbance compared to sessile species. It is also possible that mobile organisms like amphipods, isopods, gastropods, echinoderms, or pycnogonids visit pools when the tide is in-tide, in consequence these organisms may be underestimated by our sampling.
Waves could be an important physical factor structuring the system. However, many authors have observed different proportions in higher resolved webs [35].
In spite of the great seasonal variations of environmental variables that take place in these microenvironments [24] and seasonal variations in species distribution in similar environments produced by biotic factors, parameters in our study did not show significant differences by site (PC vs. CC), neither by season (Autumn vs. Spring). This indicates that community structure of intertidal tide pools is relatively robust.
When we compare the fitted slope of total links (L) vs. trophic species (S) with and without detritus/sediments against the data from Cohen, et al. [6] we can observe that the slope is lower than the theoretical value of 2 when we do not include detritus/sediments (slope=1.78; R2=0.71), but when they are included there is an increment in the number of links per node (slope=2.11). This latter value is closer to the proposed by the food web theory suggesting that these communities are working in similar way to those studied by Cohen, et al. [6] but with detritus playing an important role as energy supply.
We found that food web connectivity is higher when detritus are included as equivalent to a trophic species. This indicates the convenience of including it for food web analysis. Detritus are not trophic species in a strict sense, however they should be considered trophic species in an energetic sense [16]; in consequence, if we do not include them in the analysis, we will ignore a source of material to tide pools system that supply detritivorous and sedimentivorous species. Dunne [36] has pointed the importance of detritus inclusion in ecological food webs, emphasizing that it must be carefully thoughtout how to include it in the analysis, because non living organic matter can plays an ambiguous trophic role. Detritus are an important source of energy and must be included in the food web graphs.
All these traits suggest that intertidal food webs can be conceived as donor systems.
When we consider some energetic measurements, like metabolic rates, we found that P/R ratios are greater than 1, showing that tide pools are capable of exporting energy to the rest of the shelf system, probably throughout intermediate and top species of their food webs.
The measurement of metabolic (respiratory and photosynthetic) rates is an important step in the way of turn food web studies from topology to energetics given a more complete picture than the only topological description of food webs.