Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 3
Thiocyanate is a ubiquitous metabolite in man and animals consuming plants containing cyanogenic glycosides and thioglycosides (glucionates) while iodine is present in the earth crust since its origin and is an essential constituent of thyroid hormone requires in trace amount. Iodine is essential for thyroid hormone synthesis while thiocyanate prevents the synthesis of thyroid hormone. As a result the physiological rather functional status of thyroid is very much dependent on the balance between these ions because of their similar ionic volume and charges and competition at different steps in thyroid hormone biosynthesis. Both iodine and thiocyanate enter in the body / thyroid gland through food and water. Thiocyanate in relatively higher concentration regulate the uptake, efflux, organification of iodide, thyroid peroxidase activity and biosynthesis of thyroid hormone. In addition the retaining capacity of iodide in the thyroid gland and body also depends on thiocyanate concentration or in other words the excretion of iodine is related with thiocyanate concentration. In the semi-arid region of earth, the consumption of cyanogenic food (thiocyanate precursor) is relatively high and many regions are environmentally iodine deficient therefore the people are at the risk of iodine deficiency disorders (IDD).
<Thiocyanate is a ubiquitous metabolite in man and animals consuming plants containing cyanogenic glycosides and thioglycosides (glucionates) while iodine is present in the earth crust since its origin and is an essential constituent of thyroid hormone requires in trace amount. Iodine is essential for thyroid hormone synthesis while thiocyanate prevents the synthesis of thyroid hormone. As a result the physiological rather functional status of thyroid is very much dependent on the balance between these ions because of their similar ionic volume and charges and competition at different steps in thyroid hormone biosynthesis. Both iodine and thiocyanate enter in the body / thyroid gland through food and water. Thiocyanate in relatively higher concentration regulate the uptake, efflux, organification of iodide, thyroid peroxidase activity and biosynthesis of thyroid hormone. In addition the retaining capacity of iodide in the thyroid gland and body also depends on thiocyanate concentration or in other words the excretion of iodine is related with thiocyanate concentration. In the semi-arid region of earth, the consumption of cyanogenic food (thiocyanate precursor) is relatively high and many regions are environmentally iodine deficient therefore the people are at the risk of iodine deficiency disorders (IDD). The pregnant and lactating women and the women of childbearing age group are the most vulnerable group of IDD because the neuronal development of the fetus and neonate are greatly affected even in mid to moderate iodine deficiency. This article reviews the sources of thiocyanate and iodine in food and thyroid gland physiology in relation to thiocyanate and iodine based on experimental and epidemiological evidences.
From available literature along with our observations. thyroid gland morphology, iodide uptake, iodide influx, iodide organification, activity of thyroid peroxidase, thyroid hormone synthesis and the excretion of iodine in relation to thiocyanate concentration including thiocyanate metabolism have been discussed based on experimental and epidemiological evidences from available literature along with our observations.
In the semi-arid region of earth, the consumption of cyanogenic food is the cause for the development of goiter and associated iodine deficiency disorders (IDD). The pregnant and lactating women and the women of childbearing age group are the most vulnerable group for IDD because the neuronal development of the fetus and neonate are greatly affected even in mid to moderate iodine deficiency.
In thyroid gland iodine is an indispensable constituent for the synthesis of the thyroid hormone, thyroxine (T4) and triiodothyronine (T3) which are essential for normal growth, physical and mental development in man and animals. The most familiar effect of iodine deficiency is goitre (enlargement of thyroid gland) with a number of physiological disorders on the foetus, neonate, the child, adolescent and the adult in the whole population collectively termed as iodine deficiency disorders (IDD). The role of iodine deficiency as an environmental determination in the development of endemic goitre is established. However many agents in the environment interfere with thyroid gland morphology and function acting directly on the gland or indirectly by altering the regulatory mechanism of thyroid gland. The uptake and utilization of iodine, by the thyroid gland is impaired by the pseudo halide thiocyanate (SCN-). Thiocyanate is formed from cyanogenic substances. It is metabolized in thyroid gland. The role of thiocyanate ion in the homeostasis of thyroid is a provocative issue where IDD persists inspite of adequate iodine intake and consumption of cyanogenic plant food is relatively high. The article reviews the physiology of the thyroid gland in relation to iodine and thiocyanate metabolism.
Bioavailability
of Iodine and Thiocyanate
Iodine
There is a cycle of iodine in nature. Most iodine is present in oceans. It was present during the primordial development of earth, but large amounts were carried by wind, rivers and floods into the sea. Iodine occurs in the deeper layers of the soil and is found in oil-well effluents. Water from deep wells can provide major source of iodine. In general, the older and explored soil surface the more likely it is to be leached of iodine [1].
The dietary source of iodine is the food crops grown in the region and drinking water. Meat, fish and dairy product are also the main source of iodine. In sea fish and seaweeds contain high amounts of iodine. Supplementations of iodine through salt, water, bread are the additional sources of iodine specially in iodine deficient area.
Thiocyanate
Cyanide in trace amount is almost ubiquitous in plant kingdom and occurs mainly in the form of cyanogenic glucosides and glucosinolates (thioglucosides); both are nitrogen containing secondary metabolites share a number of common features. They derive biogenetically from amino acids and occur as glycosides which are stored in vacuoles. They function as prefabricated defense compounds that are activated by the action of a β-glucosidase in case of emergency, releasing the deterent: toxic cyanide from cyanogens or isothiocyanates from glucosinolates [2].
When the cyanogenic plants are wounded by herbivores and other organisms, the cellular compartments are broken down and the cyanogenic glucosides come in contact with an active β-glucosidase having broad specificity, which hydrolyses them to yield 2-hydroxynitrile (cyanohydrin) that is further cleaved into the corresponding aldehyde or ketone and HCN by a hydroxynitrile lyase.
HCN is highly toxic for animals and microorganism due to its inhibition on enzymes cytochrome oxidases (respiratory chain) and its binding to other enzymes containing heavy metal ions. The lethal dose of HCN in man is 0.5-3.5 mg/kg after oral administration and death of animals or man reported after the consumption of plants with cyanogenic glycosides, whose concentrations can be upto 500 mg HCN/100 g seeds. Normally 50-100 mg HCN/100g seeds and 30-200 mg/100 g leaves have been reported [3].
Animals can rapidly detoxify small amounts of HCN by rhodanese. A number of herbivores can tolerate HCN at rest in lower concentrations [4]. Cyanogens are active and potent chemical defense compounds. HCN is toxin for plants which synthesize them. To prevent autotoxicity, a detoxification pathway exists - HCN combines with L-cysteine to yield 3-cyanoalanine by β-cyanoalanine synthase, cyanoalanine is hydrolyzed by β-cyanoalanine hydrolase to L-aspargine. β-cyanoalanine synthase occurs in all plants but likely to be more in strongly cyanogenic species [2] shown in (Figure 1).
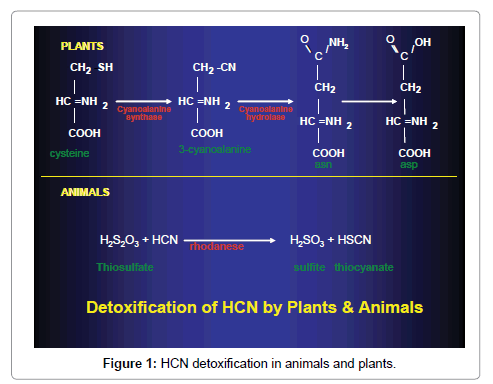
Figure 1: HCN detoxification in animals and plants.
Glucosinolate
Glucosinolates are similar to cyanogens in many respects, but they contain sulphur as an additional atom. Under hydrolysis, glucosinolates liberate D-glucose, sulphate and an unstable aglycone, which may form isothiocyanate (common name mustard oil) as main product under certain conditions, or a thiocyanate, a nitrile or cyano-epithioalkane.
All plants which sequester glucosinolates also possess thioglucoside glucohydrolases (commonly known as myrosinase) that can hydrolyze glucosinolates to D-glucose and an aglycone, spontaneously rearranging to isothiocyanate. These hydrolases are stored in the cell wall, in endoplasmic reticulum, Golgi vesicles and mitochondria.
When the tissues are wounded or disintegrated, the enzyme and its substrate come together liberating the pungent and repellant isothiocyanate. Depending on the environmental condition, enzymes and other compounds, present, the aglycone can rearrange to isothiocyanates as the most common product, or to nitriles, thiocyanates or cyano-epithioalkanes or oxazolidine-2-thiones (Figure 2).
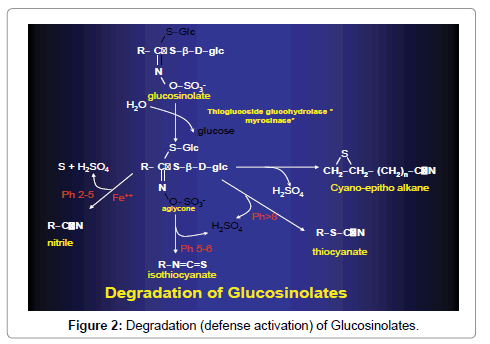
Figure 2: Degradation (defense activation) of Glucosinolates.
A number of isothiocyanates are liophilic, volatile with a pungent smell and taste, while others are not volatile and pungent smelling, but have otherwise similar properties. Isothiocyanates can penetrate through biomembranes and they have many physiological actions.
Glucosinolates are thus considered as preformed defense chemicals which are activated during emergency. They have a wide range of activities and important specially in plant-harbivore but also in plantplant and plant-microbe interactions.
Thyroid morphology/histology
The morphological profile of thyroid cells can be altered by dietary iodine. A low iodine diet causes distinctive functional alteration in thyroid cells. Some of the effects are direct result of iodine deficiency and the others are the secondary by elevated serum TSH level. In contrast high doses of iodine cause various responses depending on the dose of iodine given, the duration of the experiment and the route of administration [5].
Feeding of thiocyanate in rats at relatively high dose showed increased weight and abnormal histology of the thyroid. The histological features of the thyroid of animals from iodine non-supplemented thiocyanate added groups (-KI+SCN) indicated hypo functioning of the thyroid with a marked decrease in colloid containing follicles and significant proliferation of new follicles with indistinct lumen (hyperplasia) in contrast to the iodine supplemented control animals. The thyroid of rats deprived of KI (-KI) also showed a decreased in mature follicles and mild follicular hyperplasia. Colloid content of the follicles was however unaltered [6].
Cruciferous plants viz., cabbage, cauliflower, mustard (seeds and leaves), turnip, radish, brussel sprout, sprouts of Brassica family, bamboo shoot and cassava from non-Brassica family constitute a major portion of human diet contain naturally occurring goitrogenic substances or thiocyanate precursors (glucosinolates and cyanogenic glucoside). Extreme differences in the goitrogen content of these plants belong to same family and same taxonomy owing to their genetic and ecological backgrounds have also been reported [7]. Besides, the goitrogenic / anti-thyroid potential of the plant foods not only depend on their relative concentrations of the goitrogenic constituents as found in fresh plants but as also on their processing as foods [8]. The histological status of thyroid after prolonged feeding of cyanogenic plant foods e.g., fresh and cooked radish, turnip, cassava, bamboo shoot etc. by replacing 1/3 portion of the diet with and without iodine supplementation was investigated [9]. The thyroid follicles were lined by cuboidal cells with distinct nucleus showing hypertrophy and hyperplasia filled with less homogeneous colloid; some follicles were invaded by epithelial cell. Increase in the number of comparatively small follicles was one of the characteristic features. On the contrary, in the control rat, thyroid follicles were almost equal in size, lined by follicular cells filled with colloid. Alterations were found in the thyroid structure between the fresh and cooked cyanogenic plant fed rats. Variation in the number and size of follicular cells and colloid content was observed in KI- supplemented and non- supplemented plant fed group of rats. In addition, colloid stained more with eosin in the experimental group (-KI) as compared to control and KI- supplemented (+KI) group of rat for the variation of concentration of iodine [10,11].
Consumption of excess cyanogenic plants in relation to iodine is considered as an etiological factor for the persistence of iodine deficiency disorders in many regions. Moreover, in spite of salt iodization, residual goiter still persists [12-14].
Therefore moderate intake of iodine could be sufficient to meet the requirement but may not ensure normal thyroid in the presence of goitrogens like thiocyanate that come through cyanogenic plant which contributes a major portion of human diet of the people living in semiarid region of the world.
Iodide uptake
Thiocyanate has been considered as a possible cause of thyroid disorders because it inhibits iodide transport [15-17]. The effect of thiocyanate on iodide transport in different TSH concentrations were studied in porcine thyroid cultured cells and found that SCN concentration >10 μmol/ L inhibited iodide uptake in a dose response manner regardless of TSH concentration. It has also been reported that follicles without preincubation and with 24hr preincubation by thiocyanate showed identical inhibition [18].
cAMP production and Na+K+ - ATPase activity
cAMP production and Na+K+- ATPase activity were measured in thyroid follicular cells to investigate the mechanism of inhibition of iodide uptake by thiocyanate. The presence of 200μmol/L thiocyanate for 24hr did not inhibit TSH mediated cAMP production in the cultured thyroid follicular activities of Na+K+- ATPase in the control and the thiocyanate group were almost same [18].
All these results indicate that inhibition of iodide uptake by thiocyanate is independent of TSH concentration and that thiocyanate does not affect cAMP generation or Na+K+- ATPase activity. Then the question arises how thiocyanate inhibits iodide transport. [19] have described thyroid iodide translocator, a Na dependent iodide transport protein different from Na+K+- ATPase, in the phospholipids vesicle of plasma membrane; and speculated that thiocyanate inhibits the system [17]. Their model is also convenient to explain competitive inhibition of iodide transport as thiocyanate and iodide are common substrates for the iodide transporting protein. Competitive inhibition is important to understand the action of thiocyanate in vivo because iodine deficiency may facilitate action of thiocyanate or an excessive iodine intake may diminish the action of thiocyanate in thyroid gland.
Metabolism of thiocyanate in thyroid
The concentration of thiocyanate in thyroid is fairly constant. There is no concentration gradient for thiocyanate between the thyroid and serum, the ratio (T/S) varying from 0.29 - 0.34.Thiocyanate ion is metabolized by the thyroid of rat. Sulphate is the major sulfur product; iodine is a potent inhibitor of the metabolism of thiocyanate. On the contrary there is a maximal inhibition of the concentration of iodide by the thyroid at time when the serum level of thiocyanate ranged from 180 – 286 μmol/ml. It would appear that there is narrow ranged between the level of thiocyanate in the serum of rat and the level at which there is an inhibition of the concentration of iodide by the thyroid. Thus the role thiocyanate ion if any in the homeostasis of thyroid was found important [20].
Monovalent anions with a molecular size corresponding to that of iodine viz. SCN are concentrated in the thyroid and inhibits the normal metabolism of iodine [21]. Thiocyanate not only interferes with the uptake of iodide but also on the iodination of thyroglobulin. Thiocyanate is therefore, a potent antithyroid substance and may cause hypothyroidism at high concentrations [22]. However, short term administration of thiocyanate, serum levels of thiocyanate < 18mg/ L did not suppress thyroid function. Because thiocyanate is largely exerted through kidney and can therefore, be accumulated in the body, results from short – and long term – experiments are however, not fully comparable [22].
Iodide efflux
Thiocyanate has been reported to increase iodide efflux [23] and in vivo model thiocyanate is more potent for iodide efflux than for inhibition of iodide transport. In culture thyroid follicles thiocyanate at 10μmol/L or greater increased iodide efflux from the thyroid follicles. The degrees of iodide efflux by 10μmol/L and 100μmol/L thiocyanate were almost the same. Even the increment of iodide efflux was minor at 200μmol/L in comparison with that of 10μmol/L thiocyanate [18].
Iodide organification
Thiocyanate decreases iodine organification in a dose response manner. The decrease in iodine organification occurred in parallel to the decrease in iodide uptake. To study whether thiocyanate has an independent inhibitory action on iodine organification, thyroid follicles were incubated with the mixture of Na 125I and NaI for 2 hrs to load iodide; then 50 μmol/L thiocyanate was added. After the addition of thiocyanate, iodide uptake becomes plateau and iodine organification decreased significantly indicating that thiocyanate inhibits iodine organification independent of iodide uptake [18].
Thyroid peroxidase (TPO)
Cyanogenic constituents affect hormone synthesis in thyroid gland either by inhibiting iodide uptake or interfering the activity of thyroid peroxidase (TPO) i.e., by inhibiting the organification of iodide (I- leads to I2) or iodination of tyrosine in thyroglobulin followed by coupling reaction [24-28]. The goitrogen content of a number of cyanogenic plant foods of Indian origin and their in vitro anti thyroid activity in raw, boiled and cooked extract s with and without extra iodide have been studied by Chandra et al, (2004). The relative anti TPO potency of the studied plants and PTU equivalence was also determined by estimating the amount of plant food or PTU capable of producing 50% inhibition (IC50) of TPO activity. The IC50 was highest in bamboo shoot, followed by cassava, mustard, cauliflower, radish, turnip and cabbage. This observation was confirmed by PTU equivalence of the studied plants
After the feeding of the cyanogenic plants in experimental animals for a prolonged period, the TPO activity of thyroid of the treated animals was reduced markedly [29-42]. As mentioned cyanogenic glucosides are readily converted into active goitrogenic agent thiocyanate by glucosidases and sulphur transferase enzymes present in the plant or in the animal tissue. Thiocyanate or thiocyanate like compounds primarily inhibit iodide concentrating mechanism of the thyroid, however at high concentration thiocyanate (SCN) inhibits the incorporation of iodide into thyroglobulin by competing with iodide at the thyroid peroxidase level [43] and forming insoluble iodinated thyroglobulin in thyroid [44]. High concentration of thiocyanate is also responsible for inhibition of TPO catalyzed oxidation (I-leads to I2) [28] while glucosinolates undergo a rearrangement to form isothiocyanate derivatives [26]. Isothiocyanate reacts spontaneously with amino groups to form thiourea that interferes in thyroid gland with organification of iodide and formation of thyroid hormone and this action cannot be antagonized by the iodide [43].
T3/T4 synthesis
At low doses, thiocyanate was shown to inhibit the uptake of iodide; in addition it also affects the organic binding there by influence in the thyroxine synthesis. The latter effect of thiocyanate cannot be counteracted by excess iodide. Further, thiocyanate causes depression in protein bound thyroxine levels with concomitant increase in free thyroxine levels [45]. The reduced total circulating thyroxine levels could be a consequence of reduced thyroxine synthesis, depression in protein bound thyroxine levels and the utilization of free thyroxine levels by the peripheral tissues. Feeding of cyanogenic foods viz., bamboo shoot [39-41], radish (Raphanus sativus Linn) [34-36], cassava (Manihot esculata crantz) [34-36], maize (Zea mays Linn), cabbage, cauliflower, mastered etc. decreased the serum total T3 and T4 levels significantly. As mentioned, reduced TPO activity may be responsible for decreasing thyroid hormone levels because it regulates the synthesis of thyroid hormone.
Iodine/ thiocyanate ratio
The studies in Zaire have shown that cassava a staple diet in the region has definite antithyroid action in man and animals, resulting in the development of endemic goiter and cretinism. This action is due to the endogenous release of thiocyanate (SCN) from linamarin, a cyanogenic glucoside contained in cassava despite the fact that the cassava is consumed a large scale within tropics, however goiter and cretinism are not found in all population, where staple food is cassava. One possible explanation for the lack of goitrogenic action of cassava in some populations may be that they have a high iodine intake [46].
The development of goiter is critically related to the balance between dietary supplies of iodine and SCN. Under normal conditions, the urinary excretion of iodine (UEI) and thiocyanate (UESCN) or UEI/ UESCN or I/SCN is higher than 7. Endemic goiter develops when it reaches a critical threshold about 3 and becomes hyperendemic, complicated by endemic cretinism when it is lower than 2. The validity of this ratio as an index of the risk of development of goiter has been demonstrated by comparative studies conducted in different regions of Zaire [46].
In Manipur and Tripura of north- east India and Siddhartha nagar in the foot hills of Himalayas, universal salt iodization is in progress and the people consume adequate iodine, they also consume cyanogenic plant foods regularly in relatively considerable amount. Goiter is prevalent in the areas as mentioned and the UIE is almost adequate. In India, mean urinary thiocyanate level from non-endemic population is 0.504 ± 0.19 mg/dl [14]; the SCN levels in all those areas is much more. Therefore the contribution of SCN in the persistence of residual goiter during post salt iodization phase may not be ruled out. However the validity of I/SCN as mentioned (i.e., below 7) is not universal. This is consistent with earlier studies / report [8]; I/SCN ratio was a useful indicator in the epidemiological studies in Zaire that elucidated the goitrogenic effect of SCN from cassava consumption [46]. However the proposed use of a defined threshold of 3 for this ratio for prediction of goiter frequency has several limitations. First it requires that the ratio is not clearly defined. It has been used as both the mean of individual I/ SCN ratio, and as the ratio between mean I/ mean SCN. As there ratios are mathematically different, they yield different results. Secondly, the distribution of individual I/SCN ratio is much skewed and that if any summary statistics is to be used the median and mode are preferable to the mean. A third limitation with the use of this ratio is that very high thiocyanate load will yield serum levels that exceed the kidney threshold. Urinary excretion of SCN is therefore not linearly related to the serum levels that exert the effect on the thyroid. Finally the often pronounced seasonal variation of cyanide exposure from cassava can result in 10 to 15 fold variations of SCN and this must be considered when estimating goitrogenic effect [47].
In the semi-arid region where the agricultural production of cyanogenic plant food and consumption of thiocyanate are more, the balance between the dietary supplies of iodine (I) and thiocyanate (SCN) play important role in the etiopathogenesis of endemic goiter and associated disorders but for the prediction of this ratio is yet to be determined.
The concentration of iodine in urine is the most widely used as biochemical marker of nutritional iodine deficiency as most of the body’s iodine is excreted in the urine, usually over 90%. For surveys it is to collect 40-50 samples from an area and express the iodine as a concentration (μg/dl urine). A median urinary iodine concentration of 10 μg/dl in an area indicates no iodine deficiency [48].
Ingeston of the Brassica vegetables in human causes a rise of thiocyanate ion in blood followed by its appearance in urine. The thiocyanate level was found to drop as soon as the eating of the plants was discontinued [49]. As mentioned the urinary excretion of SCN (after a very high thiocyanate level) is not linearly related to the serum levels that exert the effect on the thyroid [50,51].
Inspite of adequate iodine nutritional status, endemic goiter is prevalent in many regions because the urinary iodine does not always truly reflect the iodine nutritional status in an environment where consumption of food containing thiocyanate precursors is relatively high [31-33]. Excess thiocyanate thus not only inhibit the iodine concentrating mechanism by inhibiting unidirectional clearance iodide from the thyroid gland but also the iodine retaining capacity of thyroid and body is also dependent on the consumption pattern of cyanogenic plant food [34-36].
In the semi-arid region, the cyanogenic plant food is grown and commonly consumed by the people and thus regular exposure of thiocyanate or its precursor is relatively high. It is also higher in cigarette smokers. When the dietary supplies of iodine and thiocyanate reach a critical point, endemic goiter and associated iodine disorders develops. On the other hand, iodine enters in the body through food and water but its availabilities vary on the geographical location. Therefore dietary supplies of iodine and thiocyanate vary from region to region depending on the availability and consumption patterns. The intake of iodine in relation to SCN and vice versa is a determinant for the causation of thyroid disorders viz. iodine deficiency disorders (IDD).
• Iodide itself is goitrogenic when it is presented in excessive in serum. Indiscriminate consumption of iodide salt regularly in environmental iodine sufficient region may be a risk factor for the development of autoimmune thyroid disorders, thyroid carcinoma, iodine- induced hypo and hyper thyroidism in long run. Conversely, intake of high thiocyanate is also goitrogenic if the intake of iodine is not satisfactory.
• Thus the question arises what should the ‘adequate’ or ‘optimum’ level of iodine? Will this remain at uniform level all over the country irrespective of its geographical distribution of iodine and consumption pattern of dietary goitrogens as cyanogenic plant foods.
• Experimental observations revealed that thiocyanate feeding inhibits iodine absorption by the mammary gland as well as by the thyroid. Such an effect on mammary gland conserves the iodine for the lactating animals but lowers iodine content milk for his young. Therefore lactating mothers ingesting thiocyanate ion could possibly cause goiter in her young affecting the development of foetus.
• Mild and moderate iodine deficiency due to thiocyanate overload associated with iodine deficiency affects intelligence, fine motor skills, problem solving capacity etc of the children and thus evaluation of their brain damage is important.