Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2012) Volume 1, Issue 1
In order to isolate xylanolytic microbial strains, screening and isolation was done using agricultural waste and decaying biomass. The enzyme super-secreter Aspergillus flavus MTCC 9390 was selected for optimized production of xylanase. Various process variables were optimized using conventional ‘one-variable-at-a-time’ approach which involves varying a single independent variable and maintaining others at a constant level. All culture conditional variables had profound influence on enzyme production and 15-30% increase was brought by nitrogen source only. A synergistic five-fold increase in xylanase production was achieved when an inoculums size of 2 x 106 spores/ mL was incubated in modified Czapek Dox-A for 6 days at pH 6.0 and temperature 45ºC under static conditions in submerged fermentation.
Keywords: Aspergillus flavus, Xylanase, Medium optimization
The complex heterogenous polysaccharide after cellulose in cereals, hardwood and fruit is xylan. These hemicelluloses interact with pectin polysaccharides and the aromatic polymer lignin, and integrate with the cellulose fibrils, creating a rigid structure which strengthens the cell wall. They also form covalent cross-links, which are thought to be involved in limiting cell growth and reducing cell wall biodegradability. Structurally, xylan consists of a β-1, 4-linked D-xylose backbone and can be substituted by different side groups such as L-arabinose, D-galactose, acetyl, feruloyl, p-coumaroyl, and D-glucuronic acid residues [1].
The biodegradation of the xylan backbone requires two classes of enzymes. Endoxylanases (EC 3.2.1.8) are able to cleave the xylan backbone into smaller oligosaccharides, which can then be degraded further to xylose by β-xylosidase (EC 3.2.1.37). Endoxylanases differ in their specificity towards the xylan polymer. Some enzymes cut randomly between unsubstituted xylose residues, whereas the activity of other endoxylanases strongly depends on the substituents on the xylose residues neighboring the attacked residues.
Production of xylanases from microorganisms has been reported to be both growth associated [2] and non-growth associated [3]. The production potential of xylanase has been found to be influenced by microbe type and its strain [4], nutrient type and concentration, growth conditions [5]. For example eubacteria and archaebacteria produce xylanase having higher temperature optima and better thermostability than those of fungi but the yield of the enzyme produced by these bacteria is comparatively lower than that produced by fungi [6].
Our group is actively engaged in the studies on industrially important microbial proteins [7-12]. The present manuscript, in continuation, describes the optimization of xylanase production from a soil isolate Aspergillus flavus MTCC 9390 using submerged fermentation.
Microorganism and inoculum
Isolation was carried out from soil samples collected from different fruit and vegetable markets of Hisar (Haryana), India. Based on the ability of xylanolysis and potential of xylanase production, a total of nine isolates were screened. Finally one fungal isolate was selected as the potential producer of xylanase which was identified at the Institute of Microbial Technology (IMTECH), Chandigarh, India as Aspergillus flavus. It had been included to the collection at IMTECH with accession number MTCC 9390. The inoculum was prepared in potato dextrose medium (PDM) containing xylan as the carbon source, by harvesting spores from 120 h old cultures grown at 30°C.
Fermentation parameters
Submerged fermentation (SmF) was carried out with 50mL broth of Czapek Dox Modified; CDM-A medium supplemented with organic nitrogen and trace metals having the following composition (gL-1 ) KH2PO4,1.0; NaNO3,2; KCl, 0.5; yeast extract, 0.5; peptone, 0.5; MgSO4.7H2 O, 0.5; FeSO4, 0.01; ZnSO4, 0.001; CuSO4, 0.0005 and xylan, 1gm. in 250mL Erlenmeyer flasks with inoculum size of 1x105 spores/ mL and incubated at 30°C for 8 days under static conditions in a BOD incubator (Remi, India). After 8 days, culture broths were centrifuged at 3000 × g for 10min and supernatant was assayed for extracellular xylanase activity.
For maximum production of xylanase, various culture conditions viz., carbon and nitrogen source, type of medium, inoculum size, pH, temperature and incubation period were optimized by conventional ‘one variable at a time’ approach which involves varying a single independent variable at a time while maintaining the others at a constant level. Fungal isolate was grown in different media, containing 0.1% xylan, at pH 6.0 for 192 h. Medium giving maximum growth was further used to standardize the inoculum size by inoculating different concentrations (1-5x106 spores/mL) of inoculums. Similarly, the temperature and time of incubation were optimized by growing the fungal isolate at different temperatures (25-50°C) for different time periods (72-192 hrs). Different organic viz., yeast extract (YE), beef extract (BE), peptone (PPT), tryptone (TPT), casein hydrolysate (CH), cotton leaf residue (CLR), soybean residue (SR) and corn powder (CP) were supplemented separately to a final concentration of 0.3 % (w/v) to study the microbial growth and xylanase activity. Glucose, fructose, galactose, arabinose, lactose, sucrose and maltose at a final concentration of 0.1% while wheat straw (WS), oat bran (OB) and birch-wood xylan (BW) were used as the carbon source at a final concentration of 1.0%.
Process variables
Effect of carbon and nitrogen sources and their concentrations, type of medium, inoculum size, pH, incubation temperature and time period, was examined in SmF. Statistical analysis was done to determine the effect of variables on production of xylanase using software ‘Statistical Package for Agricultural Scientists’ OPSTAT.
Enzyme isolation and assay
After 144 hrs of growth, the fungal broth was filtered through 4 layers of muslin cloth and centrifuged at 3000 × g for 15 min at 4°C in a refrigerated centrifuge. The supernatant taken as enzyme extract containing extracellular xylanase was used to assay the enzyme activity. Briefly, 1 mL of 1% xylan solution (in 0.05M, pH 6.0 sodium citrate buffer) was mixed with 0.1 mL enzyme solution and incubated for 15 min at 60°C. The reaction was stopped by addition of 1 mL of DNS reagent. The mixture is heated for 5 min at 100°C (boiling water bath) and then cooled in cold water. Absorbance of samples was measured at 540 nm against the substrate blank. A standard curve of xylose ranging from 0-1000 μg/mL was prepared and then quantified the released xylose in the samples using standard curve. One unit of xylanase activity is defined as the amount of enzyme liberating 1μmole of xylose equivalent under the experimental conditions in 1 min.
In recent years, considerable attention has been paid to the use of micro-organisms in industrial fermentation processes, especially enzyme production. There has been a surge of interest in the production of xylanolytic fungi for their use in food and fruit processing industry. Microbial cultures, regardless of the nature of the end product and the type of bioprocess have certain specific requirements for their growth which have to be optimized for their maximum production. The results on the optimization of culture conditions for the maximum production of xylanase by A. flavus MTCC 9390 are presented here.
Identification of xylanase producing microorganism
Out of twenty six fungal isolates, only few isolates showing clear zone (Figure 9) on agar plates containing xylan were selected as xylanase producers. These isolates were further screened based on diameter of clearing zone formation and the maximum production of xylanase on PD (potato dextrose)/ME (malt extract) media with supplement of xylan and used for further studies for optimizing culture conditions to get maximum production of xylanase. The selected fungal isolate in the present investigations had been identified at Institute of Microbial Technology (IMTECH), Chandigarh as Aspergillus flavus and was included to their collection at the centre with accession number MTCC 9390.
Optimization of culture conditions
Medium: To obtain maximum xylanase production, fungal isolate was grown on different media apart from PDA/ME media viz. Xylanase cultivation medium having xylan as a sole carbon/energy source (XCM), Czapek-Dox medium (CDM) and its modified versions (CDM- A, B, C) with 0.1% xylan under SmF. Each medium was inoculated with 1 mL inoculum containing 1x106 spores at 30°C for 8 days under static condition in BOD incubator and enzyme production was determined. Maximum activity was observed in modified Czapek Dox medium-A (CDM-A), which was used for further experiments (Figure 1). Xylanase cultivation medium which contained Mg2+ ions only showed 73% activity as compared to that in Czapek-Dox medium which contained both Fe2+ and Mg2+ ions. Further, there are reports [13-15] which suggest that organic source of nitrogen support the maximum enzyme production. Keeping this in view, the Czapek-Dox medium was modified with respect to trace metal ions and nitrogen source. Sodium nitrate was replaced with yeast extract and peptone (replaced the inorganic nitrogen source with the organic one) and Zn2+ and Cu2+ ions were added in various proportions in the modified Czepek-Dox media to supplement the medium with trace metals. The results shown in Figure 2 depicted that modified Czapek-Dox media-A (CDM-A) which contained metal ions in highest concentration (0.1%) and yeast extract and peptone (0.05% each) along with sodium nitrate (0.2%) showed the maximum xylanase production (17.09 UmL-1). The possible explanation for improvement in growth and enzyme activity at lower concentration of organic nitrogen could be that higher amount of organic nitrogen sources produce more protease activity which may degrade xylanase [16].
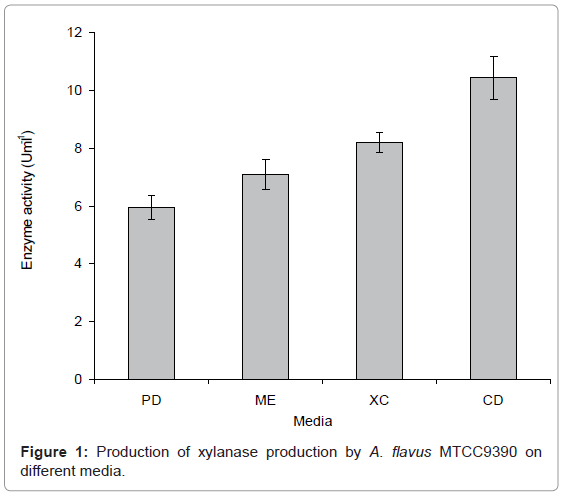
Figure 1: Production of xylanase production by A. flavus MTCC9390 on different media.

Figure 2: Effect of modified Czapek-Dox media on xylanase production by A. flavus MTCC9390.
Yeast extract and peptone has been used and recommended by many workers [2,12] as nitrogen source. Among inorganic and complex nitrogen sources, yeast extract supported the maximum enzyme production [14]. Though all related inorganic nitrogen sources supported the enzyme production but not as efficiently as yeast extract. Inorganic nitrogen such as NaNO3 (0.5%) has been reported to be the best in stimulating xylanase production by Cohliobolus sativus [13].
Inoculum concentration/size: Each 250 mL Erlenmeyer flask containing 50 mL modified CDM-A (pH 6.0) was inoculated with different sizes of working inoculum ranging from 1-5 mL taken from spore stock prepared in Tween-80 (approx.1-5x106 spores/mL) in a BOD incubator at 30°C for 8 days to determine the optimum inoculum size for xylanase production. Since 2 mL inoculum gave the maximum activity of xylanase, this size was used for further optimization process (Figure 3). Xylanase production from A. niger USM A1I and A. japonicus was reported to be maximum using 1x105 spores/mL and 1x107 spores/mL [17,18]. During the cultivation of A. foetidus maximum xylanase activity occurred when inoculum had a concentration of 1.5x108 spores/mL [19].

Figure 3: Effect of inoculum size (spore concentation) on xylanase production by A. flavus MTCC9390.
pH: To obtain maximum xylanase production by A. flavus MTCC 9390, each Erlenmeyer flask containing 50 mL modified CDM-A with pH ranging from 4.0-7.0 was incubated at 30°C with 2 mL inoculum for 8 days. After 8 days of incubation, xylanase activity was determined. Growth medium having pH 6.0 gave the maximum activity and was used for further studies (Figure 4). On either side of the optimum pH (pH 6.0) of the medium, the enzyme production decreased. The initial pH has been observed to influence the transport of enzymes across the cell membrane [13]. The growth and enzyme production by this organism has been reported to be adversely affected at alkaline pH. Similar results has been reported in A. terreus where lowest (pH 3.0 or 10.0), moderate (pH 4.0) and maximum xylanase activities (pH 6.0) were observed [20].
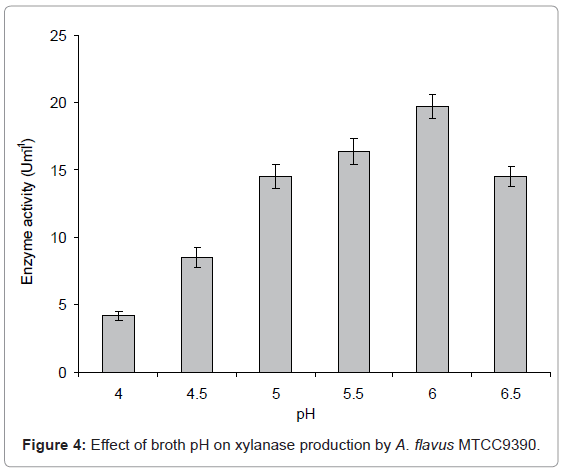
Figure 4: Effect of broth pH on xylanase production by A. flavus MTCC9390.
The effect of initial culture pH on xylanase production in P. thermophila has been reported and found maximum activity when the initial pH was adjusted to 7.0 [21]. High level of xylanase production by this strain was observed in the range of pH 5.0-8.0. Aspergillus sp. RSP- 6 was active in xylanase production over a broad range of pH from 2.0- 6.0 with maximum production at pH 3.0 [14]. The enzyme production was drastically reduced at neutral pH and no enzyme production was noticed in alkaline medium of pH 8.0. A pH of around 5.0, in general, has been observed to be optimum for xylanase production [1,19,22,23].
Incubation period: To study the effect of incubation period, each 250 mL flask containing 50 mL modified CDM-A (pH 6.0) was incubated at 30°C with 2x106 spore inoculum. After 72 hrs of incubation, the samples were harvested at regular interval of 24 hrs up to 8 days to determine xylanase activity in the supernatant of each sample. Maximum xylanase production was observed when incubated for 6 days (Figure 5). After 6 days, a decline in the activity was observed reaching a value of 16.38 UmL-1 on 8th day of incubation could be due to the release of xylanase from bound substrate or from the autolysis of fungal cells. Similar results were reported in P. oxalicum which produced maximum xylanase production after 6 days [15]. Maximum xylanolytic activity in A. terreus was observed after 4 days at 35°C and remained at that level until 6.5 days when the activity started to decrease [14]. In F. solani, enzyme production started after 24h of inoculation but showed maximum production on 6th day of incubation period at 30°C [22]. Maximum xylanase production by Aspergillus sp. RSP-6 occurred on 5th day of incubation and further increase in fermentation time resulted in reduction of activity [14].
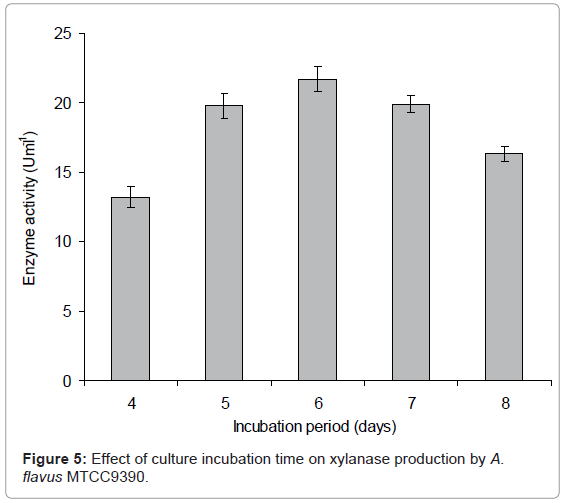
Figure 5: Effect of culture incubation time on xylanase production by A. flavus MTCC9390.
Incubation temperature: To determine the optimum temperature for maximum xylanase production by A. flavus MTCC 9390, each Erlenmeyer flask containing 50 mL modified CDM-A (pH 6.0) was inoculated with 2x106 spores and incubated for 8 days at different temperatures viz., 25, 30, 35, 40, 45 and 50°C in BOD incubator. Xylanase activity determined for 8 days of incubation was found to be maximum at 45°C (Figure 6). Further optimization was done by growing the culture at 45°C. Although the physiological changes induced by high temperatures during enzyme production is not very clear, it has been suggested that at high temperatures, microorganisms may synthesize reduced number of proteins that are probably essential for growth and other physiological processes including enzyme production [24]. By applying the temperature shift during laboratory cultivation, hydrolytic activity could almost be doubled, whereas the xylanolytic production was three fold higher in comparison to cultivation at a constant temperature of 28°C [25]. The ambient temperature of 28 ± 3°C was found to be suitable for maximum production of xylanase in A. niger [17].
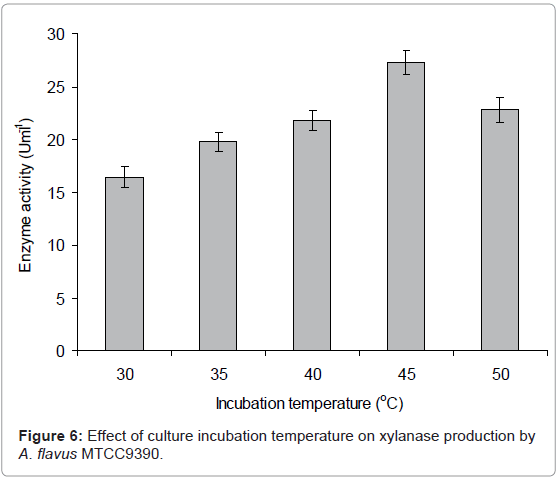
Figure 6: Effect of culture incubation temperature on xylanase production by A. flavus MTCC9390.
The best temperatures for xylanase production by A. japonicum have been reported to be 25°C [18]. With cultivation temperature lower and higher than the optimum, decline in xylanase activity has been reported [17,22]. A slightly higher temperature of 45°C and 50°C has been reported to be optimum for xylanase production by P. oxalicum and T. aurantiacus, respectively [15,26].
Carbon source: Different carbon sources were used to determine their effect on xylanase production. Each 250 mL Erlenmeyer flask containing 50 mL modified CDM-A (pH 6.0) contained either of these carbon sources viz. various commercially available sugars, wheat straw, oat bran, birch-wood xylan at a concentration of 0.1-1% each. The medium was inoculated with 2 mL inoculum and incubated at 45°C for 6 days and xylanase production was determined. Birch-wood xylan and oat bran at a concentration of 1% showed almost similar xylanase production which was 27.28 and 25.54 UmL-1, respectively (Figure 7). Though, commercial xylan also supported the xylanase production but not as efficiently as wheat straw. None of the monosaccharides and disaccharides tested viz. glucose, fructose, galactose, arabinose, maltose, lactose and sucrose was found suitable for xylanase production. Since no xylanase activity could be observed in the absence of xylan in the culture medium, it could be suggested that growing of A.flavus in xylan enriched media exerted some effect on enzyme secretion.
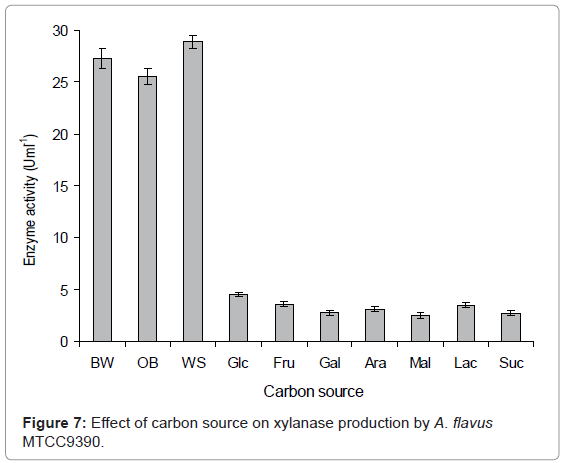
Figure 7: Effect of carbon source on xylanase production by A. flavus MTCC9390.
Induction of the synthesis of xylan-degrading enzymes by xylanolytic organisms cultured with xylan as carbon source is well documented [4]. However, xylanase production in Aspergillus sp. RSP-6 to be constitutive in nature and none of the monosaccharide or disaccharide improved the xylanase production compared to palm fiber as carbon source [14]. In contrast, xylanase activity in A. pullulans Y-2311-1 was induced by xylose and xylan [27]. Induction of xylanase activity in T. fusca and P. bryantii by glucose, cellobiose, water soluble xylan and acid-ethanol soluble xylan has been reported [28,29]. Suppression of xylanase synthesis by readily metabolizable sugars such as glucose and/or xylose has been reported in Streptomyces sp. [30].
The use of wheat straw and wheat bran as a carbon source for xylanase production has been reported [13,22,31]. Other agro-residues such as rice straw [26], sugarcane bagasse [32], corncob [12,33,34], oatspelt xylan [15] and Brewer’s spent grain [35] have also been reported as suitable substrates for xylanase production.
Nitrogen source: Effect of different nitrogen sources viz., tryptone, casein hydrolysate, soybean residue, cotton leaf residue and corn powder on xylanase production was studied by replacing yeast extract by various nitrogen sources in modified CDM-A (pH 6.0). Each 250 mL Erlenmeyer flask containing 50 mL medium (pH 6.0) was incubated with 2 mL inoculum, containing 2x106 spores, at 45°C for 6 days under static conditions, after which xylanase production was determined. Corn powder was found to be the best nitrogen source as it gave the maximum xylanase activity (Figure 8). Further to find out its optimum concentration, different concentrations of corn powder were ranging from 0.1–0.5% in modified CDM-A (pH 6.0) containing xylan as carbon source was investigated. At 0.1% concentration, xylanase production was found to be maximum (data not shown). The results obtained during the present investigations are in agreement with those reported [14] where yeast extract has been reported to be the best nitrogen source for xylanase production by Aspergillus sp. RSP-6 and on contrary, nitrogen sources such as peptone, beef extract, soybean meal, peanut meal and corn steep liquor were found to be poor nitrogen sources.
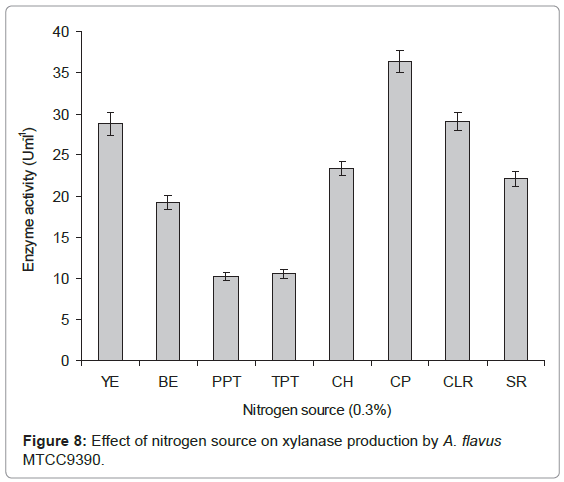
Figure 8: Effect of nitrogen source on xylanase production by A. flavus MTCC9390.

Figure 9: Agar plate showing clear zones of hydrolysis formed by xylanolytic isolates.
Among the other organic nitrogen sources, defatted rapeseed meal induced maximum enzyme production by Streptomyces sp. [36]. Corn steep liquor and soya bean meal produced maximum xylanase level in B. licheniformis A 99 [3]. In contrast to our results, peptone as the best source of organic nitrogen for the production of xylanase from A. niger, F. solani and T. harzianum [13,22,37]. However, when alternative nitrogen sources such as cotton leaf residues and soybean residues were used individually and in combination, lower xylanase activities were observed [37].
The medium optimized at this stage for xylanase production by A. flavus MTCC 9390 was termed as ‘xylanase overproduction medium’ and finally, optimized conditions were: inoculum size of 2 mL with 6 days of incubation in CDM-A medium of pH 6.0 at 45°C.
Isolation and multi-step screening method employed in the study led to the isolation of one efficient strain culture of A. flavus for production of xylanase. Results obtained on optimization of process variables under SmF revealed that wheat straw and corn powder could be effectively used for xylanase production. About five fold enhancement in xylanase production by A. flavus MTCC 9390 was achieved when an inoculum size of 3x106 spores/mL was incubated in modified Czapek-Dox medium-A at pH 6.0 and 45°C for 144 hrs. The low cost of its production and acidophilic nature of enzyme may further broaden the scopes for its use in fruit juice industries.
The authors are grateful to the CSIR-UGC for financial assistance and to the Head, Department of Biochemistry, Chaudhary Charan Singh Haryana Agricultural University for providing all basic facilities and guidance during this investigation.