Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 5
Aging is associated with metabolic changes and the development of diseases, due to the reduction of antioxidant defenses and alteration in apoptosis process. The lithium is used to treat bipolar disorder and neuroprotector; it also can be used as an antioxidant, but its function in aging has not been elucidated. The alterations in apoptosis could be associated with development oxidative stress in several tissues, although this information is unclear to aged rats. Disorders in apoptosis process and/or oxidative stress development impair the health of aged rats. The association between apoptosis with hepatic metabolism changes and oxidative stress disorder in aging is also unclear. Therefore, the objective of this work was to determine the effects of lithium on the lipid profile in serum and oxidative stress as well as apoptosis in the liver of aged rats. Twenty female Wistar rats were divided into 4 groups (n=5): the C group, with 3- month-old rats that received only water; the L group, with 3- month-old rats that received a lithium solution in drinking water; the S group, with 22- month-old rats that received water; and the SL group, with 22- month-old rats that received lithium in drinking water. The total experimental period was 30 days. All animals received standard diet ad libitum. The lithium dose took was according to neuroprotective dose and carefully monitored daily. The dyslipidemia studies were made through TC, HDL, LDL, VLDL, TG and glucose in serum. Oxidative stress analyses involved experiments with LH, TAS and ATP-SA. The apoptotic proteins analyses were made by western blot. Oxidative stress and apoptotic proteins were studied in the liver rats. The results showed that the lithium treatment reduced energy intake, aqueous solution intake and palatability in both, young and aged rats. This treatment increased TG and VLDL in the serum of animals in both groups. The treatment promoted hyperglycemia in young group, and in aged group, induced LDL enhanced as well as decreased HDL. Lithium induced enhanced Sirt 1 in livers of young and aged rats. TAS were higher in young group submitted lithium treatment. The livers of aged rats supplemented with lithium exhibited raised ATP- SA and Bax. In conclusion, the lithium induced dyslipidemia and hyperglycemia in the young, but this treatment also acted as a possible antioxidant agent, associated with Sirt 1 enhanced to protect this tissue of damages. In the aged rats, lithium promoted dyslipidemia and could induce cell death. Therefore, the supplementation could exert toxic effects in the livers of aged rats.
<Keywords: Aging; Lithium; Mitochondria; Apoptosis; Liver; Dyslipidemia; Hyperglycemia
C: Control Group; L: Young Group Treated With Lithium; SC: Senescent Group; SL: Senescent Group Treated With Lithium; ATP (SA): Atp Synthetase Activity; GSK-3: Glycogen Synthase Kinase 3; TC: Total Cholesterol; HDL: High Density Lipoprotein Cholesterol; VLDL: Very Low Density Lipoprotein Cholesterol; LDL: Low Density Lipoprotein Cholesterol; TG: Triacylglycerol; LH: Lipid Hydroperoxide; PDH: Pyruvate Dehydrogenase; TAS: Total Antioxidant Substances; ROS: Reactive Oxygen Species
The aging process promotes decline in cellular function and the accumulation of damages to macromolecules and organelles [1]. The liver diseases incidence increases during aging, while decreased the ability to resist injuries changes [2]. This fact is associated with a reduction in organs functions [3] due mainly oxidative damage and oxidative stress development [4]. Oxidative stress had been associated with damages, such as cells degeneration, membrane lipid peroxidation or LH higher, cell death by apoptosis pathway p53, Bax, Bcl2 concentration changes. Sirt 1 (nicotinamide adenine dinucleotide (NAD+ )-dependent class III histone deacetylases) affected oxidative stress in aged animals. Aged animals showed less active of these molecules in the liver, heart and lung [5].
The mitochondria have been involved in the production of ROS and development of oxidative stress. These organelles represents the main producer of ROS and presented the higher impaired by ROS during aging [6]. Dysfunction of the mitochondria affects the liver and central nervous system striatum functions [7], reduced ATP production and inducing apoptosis or cell death [8]. The liver function changes could be responsible for the metabolic alterations in this tissue, which probably modify the lipid profile (represented by TG and lipoproteins concentrations presents in blood), being associated with triggering diseases during aging. The lipid profile alteration could induce dyslipidemia condition; this fact is associated with development cardiovascular diseases [9].
Studies about substances with antioxidant capacity are important to prevent damages in the aged organisms and regulate the apoptotic process. Lithium has been described by acts as a neuroprotective agent due its capacity to modulate inflammation, mitochondrial function, and oxidative stress, thus preventing neurodegenerative diseases [10]. However, the neuroprotective role of lithium [11] was not associated with studies involving cell death, hepatic and serum metabolism in aging rats. The lithium is a monovalent cation that has been used for more than 60 years to treat bipolar disorder and depressive disorders [12]. This drug was reported to protect against Alzheimer’s disease development after chronic treatment [13].
Lithium treatment either has beneficial or can have harmful effects, depending on the dose used, duration of treatment and age of the patient [14]. Chronic treatment with lithium carbonate and aged animals were used in this work. Lithium treatment in aged rat’s livers could be prevented metabolic changes and oxidative stress as well as improving ATP production. The improvement of metabolic changes possibly may prevent the apoptosis process, avoiding diseases development in this organ. This work attempts to answer the following questions about alterations in lipid profile, oxidative stress and ATP production, associated with apoptosis process after lithium treatment in the liver, during the aging.
Animals
This work used 20 female Wistar 2BAW rats from the Federal University of Sao Paulo. This work was subjected to approval by the Ethical Committee for the Conduct of Animal Studies for this University (protocol 0311/12), and all the experiments and procedures were performed in accordance with the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health. Male Wistar rats show oxidative damage to mitochondrial DNA being 4-fold superior than in female. Therefore female rats have higher protection against oxidative damage, and consequently live longer than male rats [15]. The animals were divided into two large groups (n=10) of young and aged rats, and then subdivided into four groups (n=5). The initial body weight of the rats was 206 ± 10.49 g in the young group, and 218.46 ± 28.72 g in the aging group. The young groups aged 3 months were subdivided into the C groups that received drinking water, and the L groups that received lithium (1.05 g/L) solution in drinking water ad libitum. Animals aged 22 months were also subdivided into two groups: the SC received the same solution as the C group, and the SL group received lithium as well as the L group. All animals were fed standard chow diet, and it was kept individually housed in polypropylene cages. The animals were maintained in controlled clean-air rooms, with a temperature of 22 ± 3°C a 12- h light/ dark cycle, and a relative humidity of 60 ± 5%. The total experiment period was 30 days.
The daily lithium intake was 0.068 ± 0.007 and 0.057± 0.09 g/kg/ day in young and aging rats, respectively (L and SL). The lithium dose was determinate according to the neuroprotective dose described by Valdés and coworkers [16], and the dilution was made in water. The administrated lithium dose (g/Kg/day) was carefully evaluated, and calculated from amount of aqueous solution ingestion [17,18]. The drinking solutions and food intake were determinate as the difference between what was given and the leftover. The food consumption and drinking solutions were measured daily at the same time (9:00-11:00 h), and the body weight was determined once a week (9:00-11:00 h) [19]. The nutrition parameters, such as energy intake and palatability, were analyzed in this work [17-19].
Experimental procedures
All animals were sacrificed by decapitation; the blood was collected into a centrifuge tube; it was centrifuged at 1400 g for 10 minutes to acquire the serum. The serum was use to the lipid profile analysis through assays included glucose, TC, TG, LDL, VLDL, HDL concentrations. After finished the blood collected, samples (100 mg each) of livers from the rats were separated, placed in liquid nitrogen and stored in a -80°C freezer until Western blot analysis. Also were collected samples (200 mg each) that were used in the biochemical analysis of oxidative stress through LH, TAS and ATP-SA methods.
The samples of liver were homogenized in 5 mL of cold 0.1 M phosphate sodium buffer (pH 7.4). The homogenate was acquired through a motor driven Tefflon glass Potter and centrifuged at 10.000g for 15 minutes. After centrifugation, the supernatant was used to analyze lipid hydroperoxide (LH) and TAS. The resulting-pellets were suspended and used in the ATP-SA method.
Biochemical analysis in the serum
The following substances present in the serum as glucose, TG, TC, LDL and HDL concentration were analyzed with enzymatic methods (test kit Labtest Diagnostics Incorporated, Minas Gerais, Brazil). The VLDL concentration was analyzed in serum by Friedwald [20] method. The analyses were performed in the Pharmacia Biotech spectrophotometer at a controlled temperature within the cuvette chamber (UV/ Visible Ultropec 5000 with Swift II applications software to computer system control, 974212, Cambrigde, England, UK).
Hepatic oxidative stress analysis
LH was measured in the liver through the hydroperoxide-mediated oxidation of Fe2+, with 100 μL of sample and 900 μL of a reaction mixture containing 250, l M FeSO4, 2 5mM H2SO4, 100 μM xylenol orange and 4 mM BHT in 90% (v/v) methanol. This analyze involved the oxidation of Fe2+ by lipid hydroperoxides presents in the liver samples at low pH in the presence of the Fe3+-complexing dye xylenol orange [o-cresolsulfonphthalein-3',3"-bis(methyliminodiacetic acid sodium salt)]. This method used the ferrous oxidation Fe3+-complexing xylenol orange in a study of nanomolar lipid hydroperoxyde formation during transition metal-catalyzed glucose oxidation [21]. Antioxidant capacity of the liver, or total antioxidant substances (TAS), were analyzed through assay kit (test kit Randox Laboratories Ltd., Crumlin, Co., Antrim, UK).
The resulting-pellets homogenate resulted of liver samples were suspended at 1ml of 0.1 mol/l phosphate buffer, pH 7.4 containing 250 mmol/l mannitol, 70 mmol/l sucrose, and 1 mmol/l EDTA Adenosine triphosphate (ATP)-synthetase activity or ATP-SA (ATPase, E.C. 2.7.1.20.) [9]. ATP-SA assay is used to analyze mitochondrial dysfunction. Alteration in ATP-SA could harm ATP synthesis and induce ROS production [22]. The analyses were made-in the microplate reader (Dcom Micro-injection Automatic, Flex Station 3, Molecular Devices Microplate, California, USA) with the software SoftMax Pro (Molecular Devices, California, USA).
Western blot analyses
Western blot analyses were used to determined apoptotic proteins concentration in the livers. The tissue samples were lysed to release the proteins. Liver samples of 100 mg were homogenized in ultra Turrax equipment in the lysis buffer containing 50 mM Tris (pH7.5), 150 mM NaCl, 1 mM EDTA, 0.05% deoxicolate, 1% NP-40, 0.1% SDS (detergent sodium dodecyl sulfate). The total protein concentration was determined through Bradford method [23]. The next step involved the reduction and denaturation of livers samples, adding SDS and beta-mercaptoethanol, and submitted this samples to heat (95°C) for 5 minutes. After, the samples were applied to the gel of polyacrylamide (8-15%) followed by SDS-PAGE protocol (sodium dodecyl sulfatepolyacrylamide). The electrophoreses was made with running buffer containing 100 mM Tris, 768 mM Glycine at 100 V with 60 mA for 2 h, and the proteins were separated according to size. The protein’s transfer process to PVDF membranes (Millipore, Billerica, MA) was made in a transfer buffer containing 0.15 M Tris and 1.2 M Glycine; the electrophoresis in this case was made at 100V with 60 mA for 30 minutes. The membranes were blocked in 5% non-fat milk in TBS-T buffer (20 mM Tris, 500 mM NaCl and 0.2% Tween) for 1 h at 4°C to prevent non-specific background binding of primary and/or secondary antibodies in these membranes.
The membranes were incubated with primary antibodies to studies of proteins involving apoptosis. These antibodies were made with diluted 1:200 to p53 and Bax antibodies. The diluted antibody to antiapoptotic protein Bcl2 was 1: 200 as well as anti-aging protein Sirt 1. All membranes were incubated with primary antibodies overnight at 4°C. These membranes were washed to 4 times in TBS-T by 5 minutes each, and incubated with HRP-linked anti rabbit IgG, diluted 1:10000 for 2 hrs at 4°C. The membranes were exposed to ECL-plus reagent chemiluminescence detection (GE Healthcare, Waukesha, WI) and placed into gel documentation equipment which included the analysis system by software Gel Capture, Gel Quant (DNR Bio- Imaging Systems Ltd, Jerusalem, Israel). The antibodies for Bcl2 and Sirt 1 were from Santa Cruz (Dallas, Texas, USA). The p53, Bax antibodies and secondary antibody were from Cell Signaling (Danvers, Massachusetts, USA).
Statistical analyses
The analyses were expressed as the average ± the standard deviation (SD) in the tables and figures. Statistical comparisons were carried out by two-way analysis of variance (ANOVA), and Turkey’s post-roc test, with significant set as p<0.05 (SigmaStat software for Windows, Jandel Corporation, San Rafael, CA, USA).
Alteration in nutritional and biochemical parameters
Aged rats (SC group) presented a decreased energy intake and palatability, compared to the C group, and enhanced parameters in relation to the L group. Lithium supplementation in aged animals reduced food consumption and aqueous solution intake compared to the C group, in addition to showing reduced energy intake and palatability in comparison to animals without supplementation. Young animals treated with lithium showed a reduction in energy intake, aqueous solution intake and palatability in comparison with the control group, similar to the aged rats treated with this cation. There was not any significant change in the final body weight between the groups (Table 1). These results were important to determine comparisons between the young and aged rats. The nutritional parameters provide results to understanding the use of energy by animals and possible lipid profile or glucose disorders.
| Groups | ||||
|---|---|---|---|---|
| C | L | SC | SL | |
| Final body weight (g) | 225 ± 10 | 201 ± 11.4 | 207.5 ± 38.6 | 207.5 ± 33 |
| Food consumption (g/day) | 17.6 ± 2.1 | 14.4 ± 3.0 | 14.5 ± 1.4 | 10.6 ± 2.1a |
| Energy intake (Kcal/day) | 150 ± 11.9 | 71.3 ± 7.4a | 117.26 ± 9.0ab | 66.9 ± 6.9ac |
| Aqueous solution intake (mL/day) | 27.2 ± 5.4 | 14.8 ± 5.2a | 21.3 ± 6.4 | 12.2 ± 1.3ac |
| Palatability (%) | 57.28 ± 7.39 | 25.92 ± 2.69a | 42.64 ± 3.26ab | 24.33 ± 2.53ac |
Values were given as the mean ± standard deviation of the mean. a Statistically significant compared to the C group at p<0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05
Table 1: Final body weight, food consumption, energy intake, aqueous solution intake and palatability in young control rats (C), young rats treated with lithium (L), aged rats (SC) and aged rats treated with lithium (SL).
Senescent rats, the SC group, did not show alterations in TG and VLDL; but, did show reduced glucose, cholesterol and LDL, compared to the young group. The SL group presented normalization glucose, LDL and VLDL in comparison with the C group. Lithium supplementation in the aged group raised TG, VLDL and LDL, as well as reduced HDL compared to the senescent animals without treatment. The lithium enhanced glucose, TG and VDL, as well as reduced LDL and cholesterol without altering HDL in the serum of the young animals (Table 2). These results indicated that lithium induced hyperglycemia in the young group and dyslipidemia in young and aged rats, which could be associated with oxidative stress parameters.
| Groups | ||||
| C | L | SC | SL | |
| Glucose (mg/dL) | 118.8 ± 9.54 | 150.1 ± 8.58a | 76.05 ± 8.10ab | 95.48 ± 16.83b |
| Triacylglycerol (mg/dL) | 69.59 ± 9.57 | 166.91 ± 9.41a | 41.54 ± 8.19b | 106.97 ± 25.67abc |
| Cholesterol (mg/dL) | 125.63 ± 13.78 | 63.41 ± 9.99a | 94.46 ± 9.06ab | 109.12 ± 12.68b |
| VLDL-(mg/dL) | 13.92 ± 1.91 | 33.38 ± 1.88a | 8.31 ± 1.64b | 19.20 ± 4.46bc |
| LDL (mg/dL) | 44.73 ± 4.81 | 23.45 ± 8.48a | 22.14 ± 8.9a | 49.97 ± 11.86bc |
| HDL (mg/dL) | 61.51 ± 6.12 | 48.26 ± 3.90 | 54.54 ± 9.35 | 34.96 ± 5.28ac |
Values were given as the mean ± standard deviation of the mean. a Statistically significant compared to the C group at p<0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05.
Table 2: Glucose, triacylglycerol, cholesterol, VLDL-cholesterol (VLDL), LDLcholesterol(LDL), HDL –cholesterol (HDL) in serum of young control rats (C),young rats treated with lithium (L), aged rats (SC) and aged rats treated with lithium(SL).
Aged rats treated with lithium presented higher LH liver levels and decreased TAS in relation to the L group. LH liver concentration was enhanced in the SL group compared to the L group; however, the L group showed raised TAS in the liver of young rats (Table 3). The increase of LH liver concentration could be associated with possible development of oxidative stress while TAS represents protection against oxidative stress.
| Groups | ||||
| C | L | SC | SL | |
| LH in liver (nmol/g tissue) | 789.76 ± 111.81 | 574.36 ± 61.79 | 898.16 ± 64.48b | 953.08 ± 150.68b |
| TAS in liver (%) | 6.47 ± 0.68 | 9.05 ± 0.59a | 6.32 ± 1.08b | 6.20 ± 0.74b |
Values were given as the mean ± standard deviation of the mean. a Statistically significant compared to the C group at p < 0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05.
Table 3: Lipid hydroperoxide (LH) and total antioxidant substances (TAS) in livers of young control rats (C), young rats treated with lithium (L), aged rats (SC) and aged rats treated with lithium (SL).
Livers of aged animals did not exhibit alterations in ATP- SA; however, lithium enhanced ATP- SA in SL group in comparison with the other groups (Figure 1). The ATP-SA studies are important to analyze potential mitochondrial dysfunction and its association with oxidative stress development.
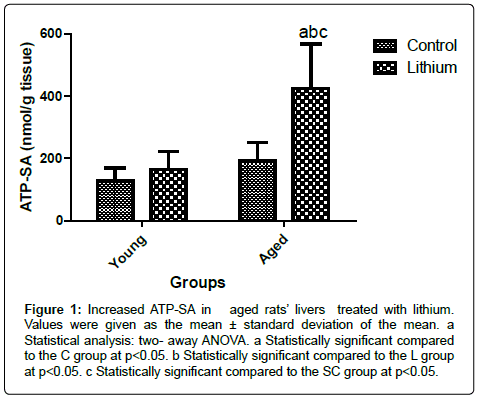
Figure 1: Increased ATP-SA in aged rats’ livers treated with lithium. Values were given as the mean ± standard deviation of the mean. a Statistical analysis: two- away ANOVA. a Statistically significant compared to the C group at p<0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05.
p53, Bax and Bcl2 in the liver
The liver of aged rats showed a reduction in p53 protein levels compared to the young groups (Figure 2). However, Bax protein was enhanced in the hepatic tissue of aged animals (Figure 3). The alterations in the Bax and p53 levels in the liver of aged rats were attributed to the aging effects, independent of the lithium supplementation. Lithium did not alter the liver of the young group in relation to the control group (Figure 3). Bcl 2 levels were decreased in the livers of aged animals compared to the C group. The treatment reduced Bcl2 concentration in both, aged and young livers of rats (Figure 4).
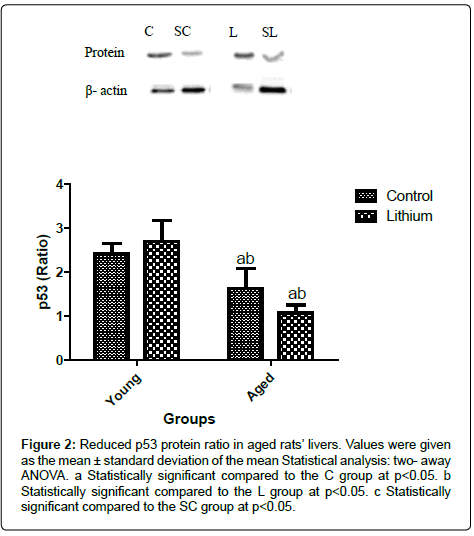
Figure 2: Reduced p53 protein ratio in aged rats’ livers. Values were given as the mean ± standard deviation of the mean Statistical analysis: two- away ANOVA. a Statistically significant compared to the C group at p<0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05.
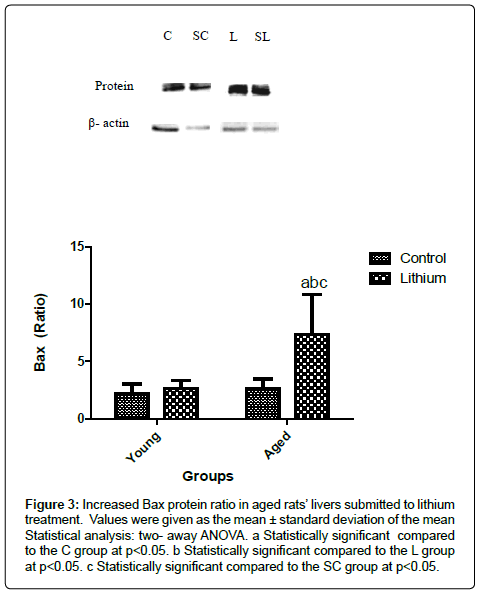
Figure 3: Increased Bax protein ratio in aged rats’ livers submitted to lithium treatment. Values were given as the mean ± standard deviation of the mean Statistical analysis: two- away ANOVA. a Statistically significant compared to the C group at p<0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05.
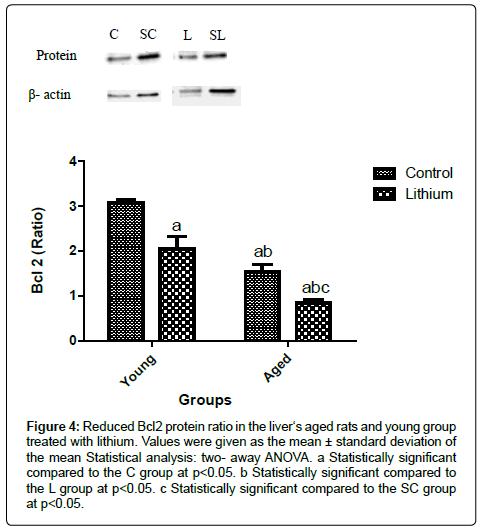
Figure 4: Reduced Bcl2 protein ratio in the liver‘s aged rats and young group treated with lithium. Values were given as the mean ± standard deviation of the mean Statistical analysis: two- away ANOVA. a Statistically significant compared to the C group at p<0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05.
The mitochondrial dysfunction modified LH or TAS, and it could induce cell death by apoptosis. Apoptosis in hepatic tissue, represented by increased in p53 or Bax, and Bcl2 reduction ratio, could be associated with lipid profile disorders. Aged rats` livers did not present alterations in Sirt 1 in relation to the C group (Figure 5). However the lithium enhanced Sirt 1 protein in the liver (Figures 5). Sirt 1 is known as antiaging protein, but its accumulation in the aged case indicates lower activity of this protein, and it could affect ROS enhanced or induce apoptosis in cells.
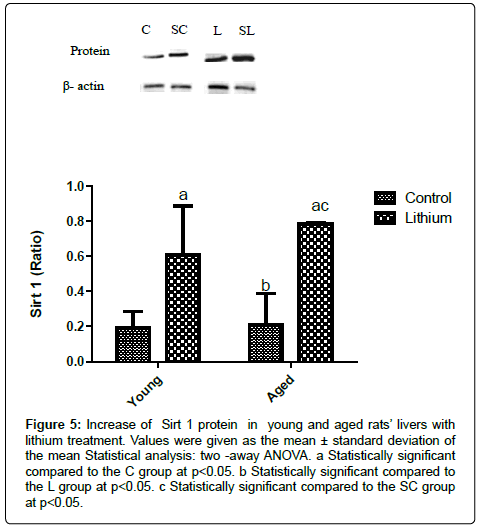
Figure 5: Increase of Sirt 1 protein in young and aged rats’ livers with lithium treatment. Values were given as the mean ± standard deviation of the mean Statistical analysis: two -away ANOVA. a Statistically significant compared to the C group at p<0.05. b Statistically significant compared to the L group at p<0.05. c Statistically significant compared to the SC group at p<0.05.
Aging is associated with the development of metabolic disorders and liver diseases, which were triggered by the decrease of antioxidant defenses and alterations in the apoptotic process [24]. This condition promotes a progressive loss in mitochondrial function in various tissues [25]. The mitochondrial dysfunctions are responsibly by reduction in the antioxidant defenses due an increase in the production of oxidants, such as ROS [3]. However, female’s rats present exhibit major mitochondrial protection when compared to male rats [15].
The choice by aged rats occurred due to decline of hepatic efficiency during aging. Aged rats presented loss of smooth-surfaced endoplasmic reticulum and therefore microsomal monooxygenases hepatics function, which could impair drugs metabolism in aged rats’ livers [26]. Microsomal monooxygenases have been associated with ROS production that, on the other hands, was involved in apoptotic responses [27]. The liver is responsible by several functions, including lipoprotein and VLDL, assembly and secretion, plasma protein synthesis, xenobioticsmetabolism and more [28]. These alterations in the hepatic tissue can harm the lipid metabolism. Lithium has been emerged as a neuroprotective agent [29], but its role as a protector in aged rats’ livers still has not been elucidated.
The lithium in the aged group did not promote significant differences in LH concentration; perhaps, the drug acted as an antioxidant agent [30] in the liver of the SL group. Mitochondria require oxygen to produce energy, and the deficiency or excessive generation of ROS is associated with damage in the organelles that promote aging [31], because more oxygen is used in lipid oxidation. However, lithium could prevent excessive generation of ROS and increase LH in hepatic tissue. This fact emphasizes the increase TAS in the liver of L group, compared with the control group (Table 3); the lithium acted in the same way in the SL group. Moreover, this treatment induced the increase Sirt 1 level in the aged rats’ livers involved in oxidative stress improvement (Figure 5).
It has been shown involvement of Sirt 1 protein in metabolism [32,33] by its acting in ROS reduction [34], and in the prevention of the mitochondrial damages. Sirt 1 has been determined to extend the lifespan of the cell and protect cell against injuries [35]. The raise in Sirt 1 in the liver of rats treated with lithium (L and SL), was followed by the improvement of ATP- SA only in SL group (Figures 1 and 5 respectively).
Sirt 1 has been reported to have a role in DNA repair in the aging process, blocking apoptosis cell death [36]. Nevertheless, the increasing Sirt 1 level induced by lithium was not able to prevent apoptosis in aged or young rats. This phenomenon is due to the decrease in its activity by carbonylation of this enzyme [37], altering its effect. Second Yuan et al. [38], Sirt 1 increased could cause accumulate of this protein in cell, associated with stress response during aging.
The p53 protein is known as a tumor suppressor, able to induce apoptosis by up-regulation of transcription proteins pathway [39]. However, lithium treatment did not produce p53 ratio alteration between studies groups; it could be involved in enhanced Bax ratio [40] as occurred in SL group only (Figures 2 and 3). The enhanced of Bax expression acts transactivation of this Bax in the cytosol, and induces the oligomerization of this Bax at the mitochondrial outer membrane to clearance pro-apoptotic factors [41]. In case of young rats, lithium treatment did not induce alteration of p53 and Bax ratio, but reduced Bcl 2 in a similar way to SL group (Figure 4).
The anti-apoptotic protein Bcl2 is closely associated with oxidative stress due its ability in modulates intracellular ROS concentration [42]. However, in the young rats, lithium drug acted as an antioxidant agent preventing excessive generation of ROS and apoptosis induction in young rats’ livers.
The aging process can be implicated in the decline of renal function and the increase of lithium levels in blood circulation and, therefore, of the lithium elimination. These factors were responsible by increase toxicity of the drug [43], and able to induce apoptotic process development, as discussed previously. Apoptosis could be implicated in the lipid metabolism disorders presents in Table 2. Boren and Bridle [44] described the inhibition of lipogenesis enzymes and enhanced de novo lipid synthesis in consequences of apoptosis activation.
The treatment with lithium promoted alterations in the lipid profile of the SL group. Lithium increased TG, VLDL and LDL in the liver of SL group, compared to the SC group (Table 2). HDL, the lipoprotein responsible by reverse transport cholesterol, was reduced in this group (Table 2). The condition which TG, LDL, and VLDL increased and HDL reduced is known as dyslipidemia, a main risk factor to cardiovascular diseases development [45]; it shows higher incidence in during aging. The level of glucose was reduced in response the diminution of food consumption and energy intake after lithium administration in SL group, although the treatment did not modify animals final body weight (Tables1 and 2). This factor was related to taste aversion conditioned by lithium-induced [46], which was followed by the reduction of aqueous solution consumption, most likely due to a reduction of palatability in both, young and aged rats (Table 1). Another factor connected with the glucose reduction would be pyruvate dehydrogenase (PDH) inhibition. PDH complex act to control the balance between glucose and fatty acid substrate oxidation, with a reduction of glucose serum concentration, resulting in steatosis pathology [47], once pyruvate was used for TG formation, which is released into the blood by liver [48].
The serum of young animals submitted to lithium supplementation presented increase in glucose, TG and VLDL, and showed a reduction of total cholesterol and LDL (Table 2). The enhance of glucose level, induced by lithium in the serum of the L group, probably promoted insulin resistance [49], because this treatment may not be able to inhibit GSK3 in young animals [47]. The insulin resistance is a main disorder of diabetes pathology due ability of lower insulin response by tissues, followed by TG and VLDL enhanced in serum [50]. The TG increase was associated with a possible enhanced of non-sterified fatty acids turnover, which could occur independently of the body fat [51], explaining the absence of alterations in the L group final body (Table 1). Insulin resistance could be responsible by the increase of TG in serum due, in part by VLDL–triglyceride overproduction, secondary to increase triglyceride synthesis in the liver [51]. The higher TG concentration had been associated with large VLDL triacylglycerolrich, which interacts with LDL. VLDL triacylglycerol -rich converts LDL in LDL-oxidized, and this fact reduced the total LDL levels [52]. Oxidized LDL are small and dense, and were not detected in method to LDL cholesterol described in this work. It was associated with a decrease of LDL and TC in young rats treated with lithium (Table 2).
In conclude, the lithium induced dyslipidemia and hyperglycemia in the young, but also acted as a possible antioxidant agent associated with Sirt 1 enhanced, to protect this tissue of damages. In the aged rats, lithium promoted dyslipidemia and could be caused cell death, however this treatment had been enhanced Sirt 1 and ATP-SA. Therefore, the supplementation with lithium may exert toxic effects in the livers of aged rats.
This article was supported by FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo, São Paulo, Brazil).