Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
![]() +44 1300 500008
+44 1300 500008
ISSN: 2572-3103
![]() +44 1300 500008
+44 1300 500008
Research Article - (2015) Volume 3, Issue 1
A unique protocol using a gelatin-based embedding technique allows long-term preservation of sea ice microalgae and phytoplankton cells. The high quality preservation of the cells and their optical properties for over two decades was confirmed after a re-examination of samples collected and prepared during 1987 at McMurdo Station, Antarctica and during 1990 at the California Bight near Los Angeles. Samples stored frozen until 2011 demonstrated the long-term preservation of the cellular structure, as well as their spectral absorption and fluorescence properties. This protocol makes it possible to assemble archives of sea ice microalgae and phytoplankton cells for environmental studies.
<Keywords: Microalgae; Absorption; Fluorescence; Chlorophyll
The proper cell preservation protocol is essential to maintain the cell structure and optical properties that existed at the time of sampling. Current preservation protocols for microalgae and phytoplankton cells typically provide utilitarian, but sub-optimal outcomes. Often, microscopic analysis of stored microalgae encounters difficulties because of poor sample preservation during storage. The advantages and disadvantages of different preservation techniques can be found throughout the literature [1-6]. Most of the phytoplankton preservation techniques, either alter the color of the cells, or chemically degrade the chlorophyll pigments [7,8].
Microphotometric analyses are highly accurate for the determination of the spectral absorption, fluorescence (excitationemission) and reflectance of individual phytoplankton cells and detrital particulates. In addition, they provide microalgae taxonomic identification, chlorophyll concentration per cell, photo-adaptive state and particle geometrical cross-sectional area [9-12]. These determinations are time intensive and difficult to perform in the field, therefore a technique capable of preserving cells without altering their optical properties during analysis and storage was developed. Preliminary research indicated that natural and cultured cells remained unchanged after samples have been transferred into a gelatin-base mixture and stored frozen at -20°C for over a year [9]. However, the full preservation capacity of this technique in relation to the cell integrity and optical properties for cells stored over two decades were only recently verified. For this particular study, the small number that could be revisited did not allow for a strict comparison to revisit the exact same cells from which the spectra analysis were originally determined. Therefore, some of the spectra comparison of previously analyzed samples to recently analyzed post-storage samples may not correspond to the same individual cells, but only the same assemblage of samples. A thorough statistical analysis was not performed due to the small number of samples that was possible to revisit. However, a visual comparison of the magnitude and shape of the absorption efficiency factor Qa (λ) and fluorescence excitation spectra of the cells before and after two decades of storage indicates that the variability within the spectra does not deviate beyond the range of the natural variability commonly observed in field samples [9,12,13]. The strength and validity of our observations focused on the cell absorption efficiency factor Qa (λ) for chlorophyll a, with peaks at 435-440 nm and 675 nm. For the fluorescence excitation spectra the emission is at 683 nm. They all represent undisputable internal markers of the photosynthetic pigments’ preservation. In all the revisited samples no changes were observed in the spectral absorption efficiency and the fluorescence excitation spectra of these internal markers.
Sample preparation
The technique is fully described in Iturriaga et al. and Iturriaga and Bower [14]. Briefly, the method consists of concentrating water samples onto Nuclepore filters (pore size of 0.4 μm) by filtration at low vacuum pressure (<5 mm Hg). Cells and particulates collected on the upper face of the filter are transferred onto a microscope slide coated with a gelatin mixture (5% gelatin, 0.2% sodium azide in 30% v/v glycerol/ water solution). The filter is then removed and the transferred sample is coated with a drop of the liquefied gelatin mixture and a cover slip placed on top. Samples can be stored for decades when kept frozen (-20°C), facilitating the collection of a large number of samples for later analyses. Cell damage on transferred samples is mainly due to filtration at high vacuum pressure. However, the cells’ conditions at the time the samples were taken can be another cause; senescent cells at different levels of integrity are present, as observed in hundreds of individually analyzed samples [10,12]. The high spectral resolution and low signal to noise ratio of the microphotometer system enables the determination of the absorption efficiency factor Qa(λ) and the fluorescence spectra (excitation or emission), with a resolution comparable to bench top instruments. From the time this method was developed no change in the optical properties has been observed in samples taken repeatedly in out of the freezer for spectral analyses measurements.
Microphotometric analysis
Absorption efficiency factor Qa(λ): The spectral absorption of single cells was determined with a photomicroscope system (Zeiss Universal, equipped with a type 03 photometer interfaced to a 150 W, Xenophot, Tungsten-Halogen light source and a scanning monochromator Type H-10, Instruments S.A.). The system is operated by a personal computer equipped with a data acquisition board that permits continuous recording (~100 measurements per nm) of the spectra from 400-750 nm. The spectral radiant fluxes for the sample (Is(λ)) and the blank (I0(λ)) allow the determination of the absorption efficiency factor of the cell or particle [9,10]. The absorption efficiency Qa(λ) is defined as the ratio of the energy absorbed by the cell to the energy impingent upon its geometrical cross-sectional area [15-17].
Qa(λ)=Is(λ)/I0(λ)
Knowing this ratio it is possible to determine the phytoplankton cell absorption (Acell (λ)) on a single cell by multiplying the cell geometric cross-sectional area (G) by its Qa(λ).
Acell (λ)=G •Qa(λ)
Fluorescence properties
Determinations of fluorescence excitation spectra were performed using the same illumination system described for the determination of Qa(λ).The contribution by scattered light is eliminated by placing a monochromator, or cut-on and cut-off filters, in combination with a narrow band-pass filter of the emission wavelength selected (e.g. 683 nm for chlorophyll) at the emission site. This therefore, assures that no other wavelengths, except that of the cell fluorescence, reaches the detector [13,14].
The embedding medium consists of a mixture of gelatin, glycerol and azide (NaN3). All these compounds have special properties for the preservation of the cell structure, intracellular pigments and cell storage under freezing. Gelatin is a heterogeneous mixture of water-soluble proteins of high average molecular weights and used in multiple applications, including coating cell culture plates to improve cell attachment, addition to PCR to help stabilize Taq DNA polymerase, as a blocking reagent in Western blotting, ELISA assays, immuno-histochemistry and several industrial and pharmaceutical uses. Cryo-protectants such as glycerol improve cell preservation by minimizing water content and/or supporting the vitrification occurrence by protecting the cell constitutive macromolecules. This is particularly relevant during the freeze-thawing that constitutes a double stress for the cells [18-20]. Azide inhibits cell respiration and photosynthesis; it is also a strong antioxidant [21-24]. Furthermore, it’s a selective and instantaneous inhibitor of mitochondrial metabolic activity [25]. The combined properties of the gelatin, glycerol and azide into a gel embedding mixture appear to have a synergistic effect and thus provide optimal preservation of the microalgae cell structure and photo pigments, and counters the effects of multiple freezing and thawing of the samples between analyses. The refractive index of the gelatin mixture is 1.036, a mid-point of the refractive index range of cells or detritus, therefore minimizing the light scattering of the cells or detritus. The gelatin mixture does not affect either the absorption, or the fluorescence properties of the cell pigments within the visible spectrum [9].
Cell response to light regimes
The solar irradiance spectral distribution in the McMurdo Sound coastal region undergoes rapid changes from the surface of the congelation ice to the benthos surface to a depth of 25-30 m, where the light field is near the limits for photosynthesis. Despite the low light levels found at those depths, an abundant and diverse population of diatoms can be found on the benthos surface [12,24]. Examples of such light adaptation can be shown by comparing the optical properties of cells collected at the ice platelet layer, such as those of Pleurosigma sp., collected at the EITN16 Station, at the ice edge, in the McMurdo Sound and benthic species, e.g.Trachyneis aspera, at the Cinder Cone Station, Cape Armitage, are described in this section.
The preservation of a Pleurosigma sp., collected and prepared during 1987 and kept stored frozen until 2011, is shown in Figure 1. The spectral absorption efficiency and fluorescence excitation spectra performed on this cell are shown in Figure 2.
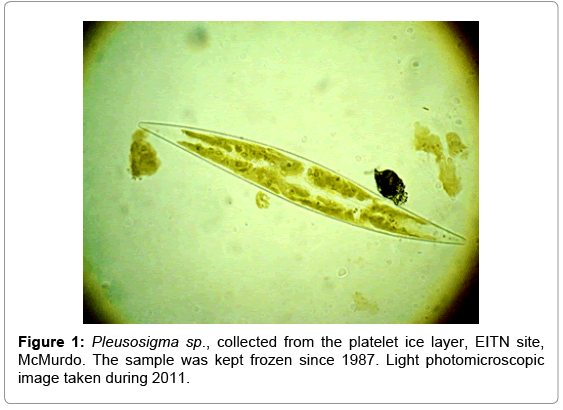
Figure 1: Pleusosigma sp., collected from the platelet ice layer, EITN site, McMurdo. The sample was kept frozen since 1987. Light photomicroscopic image taken during 2011.
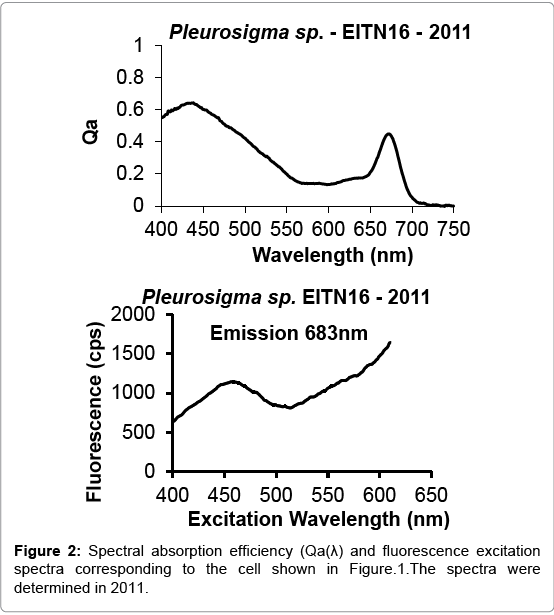
Figure 2: Spectral absorption efficiency (Qa(λ) and fluorescence excitation spectra corresponding to the cell shown in Figure.1.The spectra were determined in 2011.
The particulate matter observed around the cell shown above, illustrates detrital aggregates, which are present in all samples. Broken cells show their chloroplasts protruding, which is not the case observed in Figure 1, in which all the chloroplasts are symmetrically aligned along the cell axis. The refractive index of the medium approximates the refractive index of the cell, making it difficult to observe the cell wall under light transmission.
No major differences were found between the cell spectrum determined during 2011 shown in Figure 2 and the spectra determined in 1987 shown in Figure 3. Microphotometric analysis demonstrated that the intra-sample variability of Qa(λ) and fluorescence excitations spectra is naturally occurring, as exhibited in contiguous cells of the Nitschia kergelensis chain cell sample (Figure 4).

Figure 4: Chain of benthic Nitzschia kergelensis collected at the surface of the benthos at the IETN site. The sample was prepared in 1987. Light photomicroscopic image taken in 2011.
The spectral absorption efficiency Qa (λ) and fluorescence excitation spectra of microalga found at the platelet layer, such as of Pleurosigma sp., distinctively display an absorption maximum between 435-440 nm and a fluorescence excitation maxima between 440-460 nm shown in Figures 2 and 3. The photo adaptation observed in benthic cells is also displayed in their absorption and fluorescence excitation spectra.
Benthic microalgae
The benthic microalgae Trachyneis aspera shown in Figure 5 was preserved soon after collection in 1978 and all revisited samples seem to show no alteration of their cells structure and oil inclusions. One of the main characteristic of benthic microalgae, such as Trachyneis aspera, is their high Qa(λ) values and shape of the fluorescence excitation spectra, due to their photoacclimation to the very low and spectrally altered irradiance that reaches the surface of the sediments where they grow. The light field reaching the benthos is narrowed towards the green wavelengths (500 nm-550 nm), due to the absorption of the blue and red regions by the microalgae pigments present in the ice platelet layer and by the congelation ice respectively. The high Qa(λ) present in benthic microalgae is a consequence of their pigment packaging. While their fluorescence excitation spectra are indicative of the pigment region contributing to the transfer of energy to PS II [12]. This photoacclimation is expressed in the shape of the absorption and fluorescence excitation spectra found on Trachyneis aspera cells shown in Figures 6 and 7.
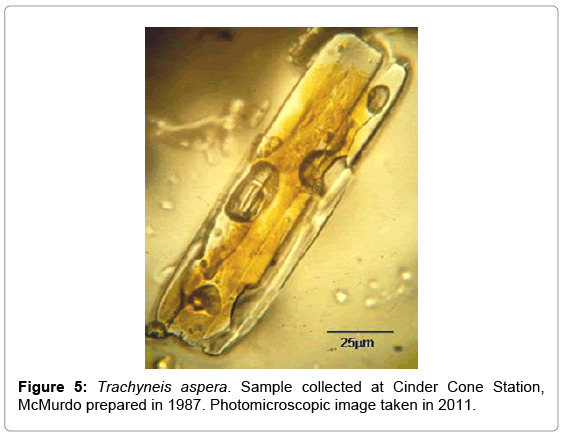
Figure 5: Trachyneis aspera. Sample collected at Cinder Cone Station, McMurdo prepared in 1987. Photomicroscopic image taken in 2011.
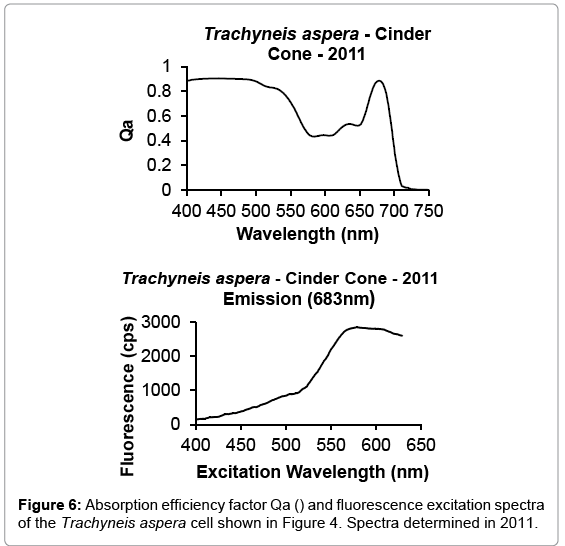
Figure 6: Absorption efficiency factor Qa () and fluorescence excitation spectra of the Trachyneis aspera cell shown in Figure 4. Spectra determined in 2011.
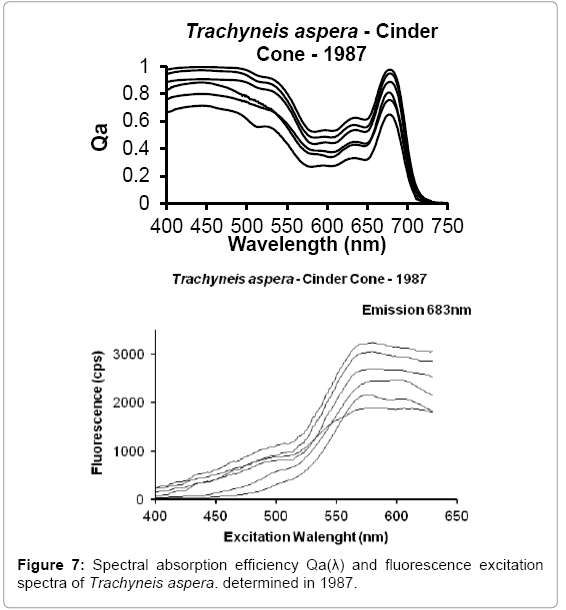
Figure 7: Spectral absorption efficiency Qa(λ) and fluorescence excitation spectra of Trachyneis aspera. determined in 1987.
A comparison of the Qa(λ) and fluorescence excitation spectra of the Trachyneis aspera cell determined during 2011, with spectra determined on other cells of the same assemblage collected and determined during 1987 (Figure 7), indicate that all values are within the range of the intrasample variability observed in field samples.
Fluorescence excitation spectra are highly indicative of the photosynthetic pigments preservation. These spectra also enable the detection of the photoacclimation state of the cells in relation to the spectral light field conditions to which they were exposed before collection. This is illustrated in Figures 2-6. Cells, that were exposed to a broader light field, such as those found at the platelet layer (e.g. Pleurosigma sp.), the excitation maxima was centered between 435-450 nm. On the other hand, benthic cells which were exposed to a highly attenuated and spectrally limited light field (e.g.Trachyneis aspera), the excitation maxima was centered at 575 nm. Similar findings were reported by Robinson et.al. 1995.
Intrasample variability
Microphotometric analyses have shown that a wide range of variability within the cells absorption and fluorescence properties can be found in natural phytoplankton cells and sea ice microalgae [10,12]. This variability is characteristic not only within individual species, but for clonal populations as observed in cell chains, where genetically identical contiguous cells of the same species are not only close to one another, but exposed to the same environmental conditions. As a result, a range of variability within the optical absorption and fluorescence spectra is expected to occur within the same species in any sample, which cannot be determined through bulk optical measurements. Benthic diatoms are less exposed to sudden changes of light or nutrients and despite the close proximity between cells in chain cells, the intrasample variability is distinctly displayed in the absorption efficiency Qa(λ) and fluorescence excitation spectra shown in a cell chain of Nitzschia kergelensis obtained from the benthos at the EITN Station, McMurdo (Figures 4 and 8).
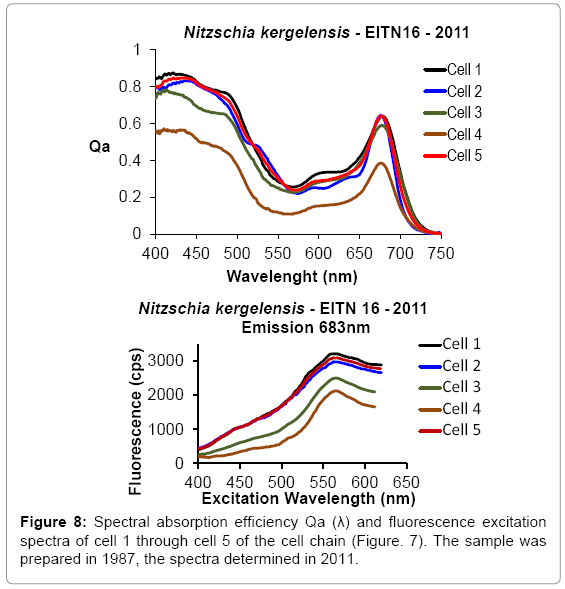
Figure 8: Spectral absorption efficiency Qa (λ) and Spectral-absorption excitation spectra of cell 1 through cell 5 of the cell chain (Figure. 7). The sample was prepared in 1987, the spectra determined in 2011.
Dinoflagellates are particularly difficult to preserve intact for long periods using classical preservation techniques. However, determinations performed on specimens of Lingulodinium sp., prepared using the gelatin-based technique revealed that embedded cells were able to keep their structure and optical properties after being stored frozen for 21 years.
The cell structure illustrated in the lower picture is not deteriorated. The indentations around the cell correspond to either the cingulum, or the sulcus, both present in dinoflagellates. The picture above shows the cell flagella, indicating that delicate dinoflagellate cells such as these are able to maintain their structure using the transfer technique described.
Figures 9-11 represent a comparison of the Qa() absorption and fluorescence excitation spectra of Lingulodinium sp. cells performed in 2011, with spectra originally determined in 1990 on cells of the same assemblage. Both spectra were within the range of variability observed in field samples. The presence of photo protectants, such as carotenoids, overlapping the spectral absorption of photosynthetic pigments in the 400-450 nm region, including chlorophyll a, (maximum absorbance at 435-440 nm), is revealed in the fluorescence excitation spectra.
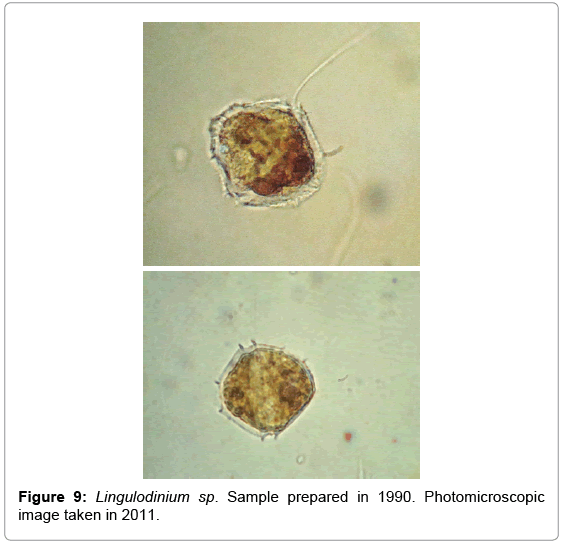
Figure 9: Lingulodinium sp. Sample prepared in 1990. Photomicroscopic image taken in 2011.
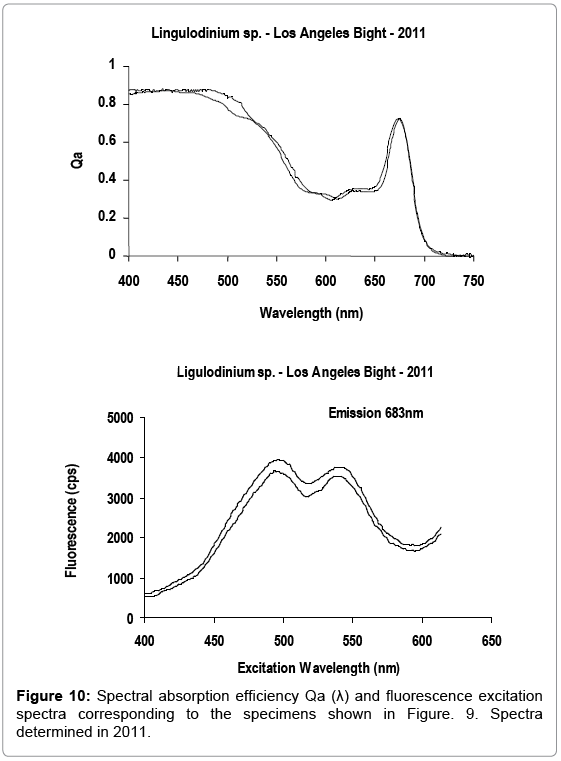
Figure 10: Spectral absorption efficiency Qa (λ) and fluorescence excitation spectra corresponding to the specimens shown in Figure. 9. Spectra determined in 2011.
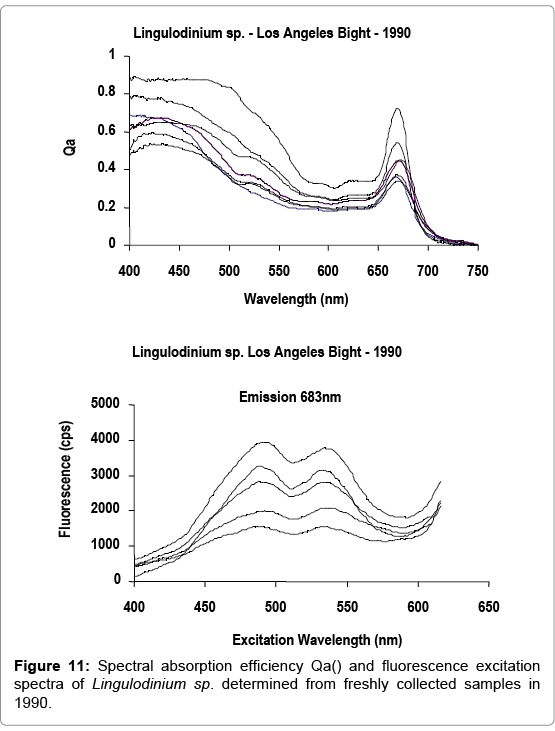
Figure 11: Spectral absorption efficiency Qa() and fluorescence spectra of Lingulodinium sp. determined from freshly collected samples in 1990.
Preservation of other microorganisms
Protozoans and small zooplankton observed in some of the samples collected in the upper sediments at the Cinder Cone station, indicated that the preservation capabilities of the gelatin-based technique extends to other cell types besides microalgae. An example of possible applications of this technique is shown in Figure 12.
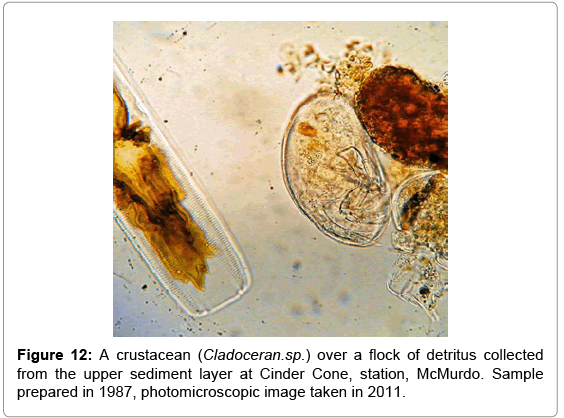
Figure 12: A crustacean (Cladoceran.sp.) over a flock of detritus collected from the upper sediment layer at Cinder Cone, station, McMurdo. Sample prepared in 1987, photomicroscopic image taken in 2011.
The gelatin-embedding technique enables the preservation of natural microalgae cells and their optical properties when samples are kept frozen at -20°C for extended periods of time. Cell damage on transferred samples is mainly caused by filtration at high vacuum pressure. However, the cells’ conditions at the time the samples were taken can be another cause; senescent cells at different levels of integrity are present in most field samples, as observed in hundreds of individually analyzed cells [10,12]. Given that the refractive index of the embedding medium approximates the refractive index of the cells, and only a cross-section in two dimensions is observed on transferred cells, it is difficult to observe the cell wall with higher resolution under light transmission. The focal depth (determined by the length of the optic axis of the microscope) is adjusted to the plane position of the chloroplasts, which are used as a reference before running the spectra. This may also affect the resolution needed to observe the cell wall. This technique coupled to microphotometric analyses enables the study of the cells pigments’ concentration per cell through different time periods, as well as their photoacclimation state, which are distinctively expressed by the shape of the absorption and fluorescence excitation spectra. These types of analyses also provide information about the intrasample variability within cells of the same species, or specific taxonomic groups, which cannot be determined through bulk optical analyses.
During this study, a thorough statistical analysis was not performed due to the small number of samples that could be revisited. However, a visual examination of the cells structure and comparison of the values and shape of the Qa(λ) and the fluorescence excitation spectra, indicated that the optical properties of the revisited samples remained well preserved without any detectable changes for over 24 years of storage. The strength and validity of our observations focused on the spectral absorption and fluorescence excitation spectra of the cells’ pigments. The efficiency factor Qa (λ) for chlorophyll a, with peaks at 435-440 nm and 675 nm and the fluorescence excitation spectra (emission at 683 nm) represent undisputable internal markers of the cells photosynthetic pigments preservation. Furthermore, the photoacclimation state observed on the platelet layer and benthic cells remained unaltered after storage. Preservation of fluorescence on stored field samples for such a long period has not been reported.
New molecular techniques may soon facilitate the identification of microalgae taxonomic groups offering new tools to incorporate the phytoplankton species composition. However, the species specific optical properties, including their photoacclimation response, as a function of the prevailing light field and other environmental conditions, have yet to be analyzed separately in order to understand their response to changing conditions. Taking into account the excellent preservation of the photosynthetic pigments, the cells’ DNA may well remain unaltered in the embedded cells. A combined effort utilizing molecular and optical analyses of individual species could provide a better understanding of the photoacclimation response of microalgae to environmental changes.
We wish to thank Lisa Campbell, John Marra, Bridgett Segers and two anonimous reviewers for their helpful comments and suggestions.
