Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 2
The co-transformation system using mixture of two Agrobacterium cultures, one harbouring bacterial mannitol-1- phosphate dehydrogenase (mtlD) as gene of interest, whereas the other harboring the neomycin phosphotransferase (nptII) marker gene for selection, was successfully used to develop selectable marker-free transgenics of tomato with engineered mannitol accumulation, to improve tolerance against multiple abiotic stresses. We have achieved high co-transformation frequency (up to 24%), and segregation frequency (up to 22-24%) for obtaining marker-free transgenics. Marker-free transgenic tomato plants resulted in mannitol accumulation, as evident by HPLC analysis, that leads to substantial protection of the vigour of the plants under multiple abiotic stress conditions, such as high salinity (up to 200 mM NaCl), drought (up to 15% PEG), and heavy-metal toxicity (up to 2 mM CdCl2). Under stress, transgenics performed well in comparison to wild type (WT), with high percentage of seed germination and seedling growth both in vitro and in vivo. They also performed better than WT, in terms of high chlorophyll content and other photosynthetic parameters such as Fv/Fm ratio, Electron Transport Rate (ETR), photosynthetic yield, and low Non Photochemical Quenching (NPQ). Thus, high mannitol level not only increases the nutritional value of the fruit, but also improves tolerance of transgenic tomato plants by increasing its physiological and photosynthetic efficiency, suggesting that carbohydrate alterations produced by mannitol biosynthesis may be linked to the stress response.
<Keywords: Abiotic stress, Co-transformation, Marker-free transgenics, Mannitol 1-phosphate dehydrogenase, Mannitol, Tomato
Tomato (Solanum lycopersicum) is one of the most popular and widely consumed vegetables grown worldwide [1]. It is a rich source of vitamins (A and C), nutrients, minerals and health promoting compounds, such as lycopene, whose antioxidant and anticarcinogenic nature is very well studied [2-5]. Recently, the tomato genome has been successfully sequenced and assembled, that open the areas of intensive research such as functional genomics, proteomics and metabolomics of tomato, as till now all the species of tomato are highly susceptible to all kind of abiotic stresses [6].
Abiotic stresses such as salinity, drought and heavy-metal toxicity are major factors limiting the productivity and adversely affect the vegetative and reproductive phases of tomato, and ultimately reduce the yield and fruit quality [7,8]. According to FAO [9], approximately 22% of the agricultural land is saline globally, and areas under drought and heavy-metal toxicity are already expanding and expected to expand further. Therefore, it is of significant importance to produce transgenic cultivars that can tolerate these stresses, so as to increase crop yield and thus, meet the increasing need of food production [8,10,11].
To cope up with the adverse environmental conditions, plants adapt themselves by employing various mechanisms, such as shifts in physiology and increased expression of stress-associated genes, leading to the formation of low-molecular-weight, non-toxic metabolites widely known as osmoprotectants or osmolytes [12-14]. These osmolytes include mannitol, fructans, proline, trehalose, myoinositol, glycine betaine and polyamines (PAs), which accumulate in significant amounts, and are helpful in maintaining osmotic potential, ionic balance, membrane integrity and oxygen free radical level, and in protecting chromatin under stress conditions [12,14-22].
Mannitol is a six-carbon non-cyclic sugar alcohol, widely distributed in nature, and synthesized in bacteria, fungi, algae, and lichens, and in more than 100 species of vascular plants, including many families like Brassicaceae, Cucurbitaceae, Fabaceae, etc. [23]. Mannitol plays an important role in stress tolerance [14,24]. In higher plants, mannitol, if present at all, is only present in trace amounts [25], that are inadequate to contribute to abiotic stress tolerance. For instance, the expression of bacterial mannitol-1-phosphodehydrogenase (mtlD) gene in transgenic tobacco [26-28], Arabidopsis [29], eggplant [14], rice [30], wheat [16] and pine [31] has led to the accumulation of mannitol in the cytoplasm, and increased tolerance to salt stress. Later, this gene was targeted into chloroplast of tobacco, and they have observed the accumulation of mannitol in the chloroplast and enhanced tolerance to oxidative stress [32,33].
In order to develop transgenics, selection marker genes (SMGs) are used mainly for the initial screening of the putative transgenic shoots, to identify the transformed plants from the untransformed ones [34]. SMGs are conditionally dominant genes that confer an ability to grow in the presence of applied selective agents (such as antibiotics and herbicides), that are normally toxic to plant cells or inhibitory to plant growth [35]. Subsequent to the generation of transgenic plants, the presence of these SMGs becomes no more of practical utility, and thus, arguably a matter of public euphoria, speculating the risk they can pose to the environment and health [36-38]. The development of markerfree transgenic plants could thus solve the issues of biological and biosafety in the genetically engineered crops, besides supporting multiple transformation cycles for transgene pyramiding [39,40]. Several strategies that have been used for the elimination of selectable markers include co-transformation [41-43], multi-auto-transformation system (MAT) [44,45], site-specific recombination system [46,47], transposon based marker methods, Intra-chromosomal recombination system and transplastomics [48,49].
In the present study, for the first time, marker-free transgenic tomato plants were generated via co-transformation strategy, by the introduction of the mannitol 1-phosphate dehydrogenase (mtlD) gene of E. coli, under the control of the cauliflower mosaic virus (CaMV35S), for mannitol accumulation, as previous work with model transgenic plants has demonstrated that cellular accumulation of mannitol confers abiotic stress tolerance.
Plant material and plasmid
The seeds of tomato (Solanum lycopersicum) variety “Pusa Ruby” were obtained from National Seeds Corporation, Indian Agriculture Research Institute, New Delhi, India. We have used two binary plasmids, pMVRmtlD (-HPT) having mtlD gene without plant selection marker (HPT), and pCAMBIA2300 having nptII gene as plant selection marker, for co-transformation (Figure 1a and 1b). Both plasmids were confirmed by PCR and restriction-digestion, and then mobilized into Agrobacterium tumefaciens strain LBA 4404 separately by freeze thaw method for tomato transformation.
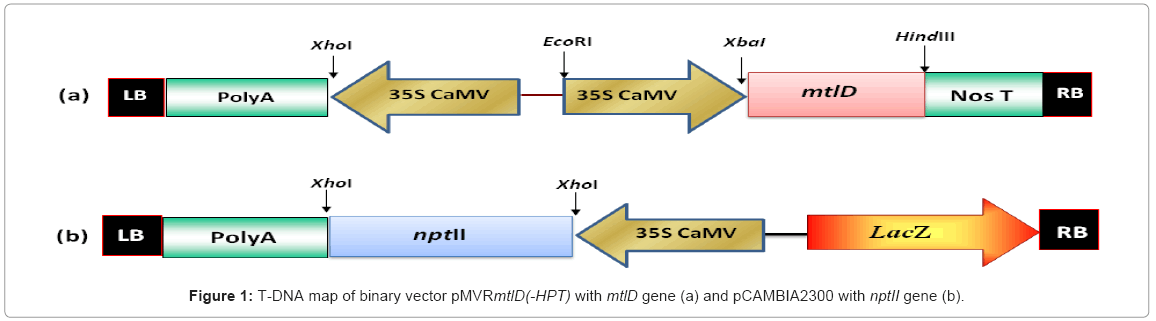
Figure 1: T-DNA map of binary vector pMVRmtlD(-HPT) with mtlD gene (a) and pCAMBIA2300 with nptII gene (b).
Tomato transformation and regeneration
Tomato seedlings were raised, and fully expanded cotyledons were collected from 10-12 days old seedlings. After surface sterilization, the cotyledon explants were trimmed on all sides and placed with abaxial side up for 2 days of pre-culture on shoot regeneration medium (SRM), i.e. Murashige and Skoog (MS) medium [50], fortified with 2.5 mg/l BAP+0.5 mg/l IAA+0.5 mM putrescine (Put)+3% maltose [51]. For co-transformation, two different Agrobacterium (LBA 4404): one containing binary vector pMVRmtlD (-HPT) and other containing binary vector pCAMBIA2300 were cultured overnight. The Optical Density (O.D.) of each Agrobacterium culture was 0.2 to 0.4, and was mixed in 1:1 ratio to transform the cotyledonary explants for 10 min. The infected explants were transferred to co-cultivation medium (SRM) for two days, and then to selection medium (SRM containing 30 mg/l kanamycin and 300 mg/l augmentin) for about two months, with subculture every fortnight at 26 ± 1ºC and 16 h photoperiod with irradiance of 40 μE mol m-2s-1. The regenerated small shoots were transferred to shoot proliferation medium (MS+0.5 mg/l BAP+300 mg/l augmentin), and after about 30 days, the well-grown individual shoots were excised and transferred to rooting medium (1/2 MS+augmentin 300 mg/l). The rooted plants were transferred to pots and after two weeks of hardening, they were transferred to transgenic green-house [51].
Polymerase chain reaction
The putative cotransformants and their progenies were analyzed by PCR, for the integration of the transgene to screen out marker-free transgenics. DNA was isolated from the leaves by CTAB method [52]. About 100 ng of DNA from untransformed plants, as well as putative transgenic lines, was taken and mixed with 100 mM of primer pair of mtlD and npt II genes individually, 1X PCR buffer, 2 mM MgCl2, 100 μM dNTP mix and 0.5 U of Taq polymerase (BIOTOOLS). The PCR program included denaturation at 94°C for 5 min, followed by 30 cycles of denaturation at 94°C for 1 min, annealing at appropriate temperature (59°C for npt II and 50°C for mtlD) for 1 min, and synthesis at 72°C for 2 min, and final extention for 10 min at 72°C. The primer pairs specific for the amplification of 1 kb fragment of mtlD gene is 5’-TGT TGC GGA CCT CGG AGA CAT TCT T-3’ (FP) and 5’-ACG CCG TGA TAG TCG CAC TTG AGC-3’ (RP), and 750 bp fragment of npt II gene is 5’-CGC ATG ATT GAA CAA GAT GGATTG CAC-3’ (FP) and 5’-TGA TCA GAA CTC GTC AAG AAG-3’ (RP). The PCR products were analyzed on 1% agarose gel.
Southern blot hybridization
Tomato genomic DNA (10 μg) was restricted with EcoRI to detect the copy number of the transgene. Southern blots were prepared by standard procedure [53], using Hybond-N Nylon membrane (MDI). The mtlD and npt II gene probes were prepared using 32P-labeled dCTP by Random priming kit, as per the manufactures guidelines (Takara, Japan). Hybridization was carried out for 18-24 h at 42°C. After washing, the membrane was exposed to X-ray film (Kodak) at -70ºC, and finally developed to detect signals.
RNA extraction, RT-PCR and northern blot
Total RNA was isolated using TRIzol® (Invitrogen) and iRIS kit (IHBT, India), treated with RNase-free DNase (supplied by Taurus Scientific), and was used as a template for doing one step RT-PCR reaction. The 25 μl of reaction mixture included 1X RT-PCR buffer, 10 mM of dNTP mix, 5 μM of each primer, 40 U of RNase inhibitor, 2.5 U Reverse Transcriptase and Taq Polymerase each, and 250 ng of RNA. The reaction mixture was incubated at 48ºC for 30 min. After reverse transcription by omniscript and sensiscript reverse transcriptase, reaction mixture was heated at 94ºC for 5 min, followed by 35 cycles of 30 s denaturation at 94°C, primer annealing at 59°C for 30 s, extension at 72°C for 1 min, and final extension for 10 min. The PCR products were analyzed on 1.2% agarose gel. For Northern blotting, about 25 micrograms of total RNA was electrophoresed through a denaturing formaldehyde (18% v/v)/agarose gel (1.1%), and blotted to a Nylon membrane by the capillary blot method. Prehybridization and hybridization were done at 42ºC for 8 and 18 h, respectively, using an [32P]-labeled mtlD cDNA probe. Washing of the membrane was carried out first with 2X SSC, 0.1% SDS for 5 min, and then 0.2X SSC, 0.1% SDS at 65ºC for 5 min, finally exposed to X-ray film.
Transgene segregation analysis
Transgenic tomato T1 seeds were inoculated on ½ MS medium in sterile Petriplate without any antibiotics. Once the seedlings were grown upto two leaf stages, the DNA was isolated and checked for the presence and absence of mtlD and npt II gene by PCR analysis.
Mannitol estimation
Mannitol estimation was done in all the marker-free mtlD tomato transgenics, before and after stress conditions. Mannitol was extracted from leaf tissue by grinding 1 gm (fresh weight) of the frozen tissue with 1-3 ml of methanol/ chloroform/water, 12:5:3 (v/v), at 4ºC [26]. The extract was centrifuged at 1000×g for 5 min, and supernatant was collected and vacuum dried. The resulting pellet was resuspeded in 100 μl of water, and passed through the nylon membrane to remove hydrophobic compounds. The purified samples were used for mannitol estimation. HPLC analysis was performed on Shimadzu LC-10A system using RID detector.
Abiotic stress tolerance assays
The tolerance of the marker-free tomato transgenics lines and untransformed control (UT) to multiple abiotic stress conditions, such as salinity (NaCl), drought (PEG) and heavy-metal (CdCl2) was tested by the following methods.
In vitro seed germination assay
The seeds from untransformed control and T2 transgenic lines were surface-sterilized and inoculated on MS basal medium supplemented with NaCl (150 and 200 mM), PEG (MW 8000) [10% and 15%] and CdCl2 (1 and 2 mM) and incubated at 26°C ± 1. The UT and transgenic seeds were inoculated on the half- half portion of the same Petriplate. Tolerance was considered on percent seed germination. In case of control, the seeds were germinated on MS medium without any stress agent. After one month, data was scored for percent seed germination [54].
In vitro seedling growth assay
The seedlings of UT and T2 transgenic lines with well-developed root system were grown in test tubes containing liquid MS (1/10th strength) medium, supplemented with 200 mM NaCl (salt stress) or 15% PEG, MW 8000 (drought stress) or 2 mM CdCl2 (heavy-metal). The tolerance was measured on the surviva, and growth of the seedlings under stressed conditions. Data on seedling height, fresh weight and dry weight was scored after a period of one month for control and transgenic seedlings grown under stress conditions. The control seedlings were maintained in the same manner, but without stress agents [54].
In vivo seedling growth assay
Three-weeks-old PCR positive T1 and T2 seedlings of transgenic lines with well developed root system were grown in plastic pots containing vermiculite: soil (1:1) mix, along with UT seedlings. 15 ml of 1/10th MS solution supplemented with 200 mM NaCl (salt stress), 15% PEG (drought), and 2 mM CdCl2 (heavy-metal) was poured per pot once a day for 14 days. The tolerance was based on the survival and growth of seedlings under stressed conditions [54].
Chlorophyll determination
Tomato leaves (0.1 g) were homogenized with 80% acetone (v/v), and the total amount of chlorophyll was determined spectrophotometrically, according to the formulae given by Arnon [55]. Total amount of chlorophyll was estimated in UT and transgenic lines before and after salinity, drought and heavy-metal stress treatment.
Photosynthetic efficiency assessment
One month old tomato transgenic and UT seedlings were exposed to different abiotic stress conditions for 4-5 days. The other set containing UT, as well as transgenic seedlings without stress treatments, was considered as control. At the end of the abiotic stress treatment period, stressed and control plants were pre-darkened for 40 min, before measuring the different photosynthetic parameters, such as chlorophyll-fluorescence (Fv/Fm), photosynthetic Electron Transport Rates (ETR), yield of Photosystem II (YII) and Non-Photochemical quenching (NPQ), by using JUNIOR-PAM Chlorophyll fluorometer system (H. Walz, Effeltrich, Germany) under photosynthetic steady-state conditions, using a photon flux density of 1550 μmol m-2s-1 as actinic light and 10 mM m-2s-1 for the saturating flashes (with a duration of 0.8 s). JUNIOR-PAM fluorometer system contains WINCONTROL software (Walz, Germany) [56]. Treatments and measurements were carried out on uniform leaves.
Data analysis
All the experiments were repeated 2-3 times, and the data presented are average (mean) with the standard error from all the experiments. The data were analyzed statistically using student’s t-test.
Co-transformation, regeneration and PCR analysis of primary (T0) co-transformants
The Agrobacterium-mediated co-transformation of tomato cotyledonary explants was successfully done. The co-cultivated explants exhibited the appearance of shoot buds within 10-15 days of transfer to selection medium. Several putative transgenic tomato plants were generated, which were found to be normal like UT plants in morphology, vegetative and reproductive growth. The primary co-transformants (CoT) were analysed by PCR for the presence of transgene. About 83% (25/30) of putative CoT lines were found to be positive for nptII marker gene (Figure 2a). PCR analysis of same lines using mtlD gene-specific primers revealed an amplicon of 1 kb in about six (lane 3, 5, 6, 9, 11 and 17) of them (Figure 2b) Thus, the co-transformation frequency was about 24% (6/25). The co-transformants (CoT), containing both nptII gene and mtlD gene, were transferred to soil and maintained in transgenic net-house. These transgenic lines were designated as CoT 3, 5, 6, 9, 11 and 17. The seeds were collected from these primary transgenics and used for the segregation analysis of both the genes (mtlD and nptII) in T1 progenies.
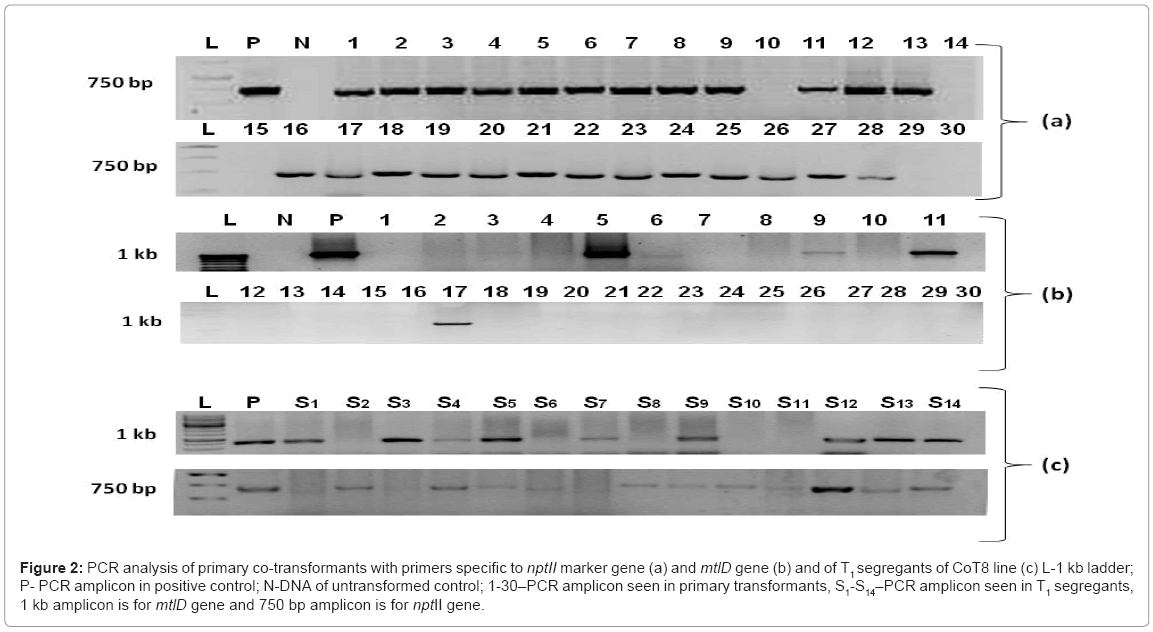
Figure 2: PCR analysis of primary co-transformants with primers specific to nptII marker gene (a) and mtlD gene (b) and of T1 segregants of CoT8 line (c) L-1 kb ladder; P- PCR amplicon in positive control; N-DNA of untransformed control; 1-30–PCR amplicon seen in primary transformants, S1-S14–PCR amplicon seen in T1 segregants, 1 kb amplicon is for mtlD gene and 750 bp amplicon is for nptII gene.
Segregation analysis of transgenes
All the six confirmed co-transformed T0 plants were selfed; seeds were collected and germinated on half-strength MS medium. In order to get marker-free plants, we used the PCR to screen the large amount of progenies. In PCR analysis of CoT6 line, nptII marker gene was found in about 75% of segregants, while 22% of the progenies contained only mtlD gene without nptII selection marker like S1, S3 and S7 (Figure 2c). The segregation frequency of the mtlD and the nptII genes also differed among different transgenic lines (Table 1). For example, in the case of transgenic line CoT3, about 24% of the T1 seedlings were confirmed to be marker-free, but only 3% in case of CoT11 (Table 1).
| Transgenic line (T1) | Total number of T1 progenies screened by PCR analysis | Number of T1 progeny screened (PCR analysis) | % of marker-free mtlD transgenics obtained | |||
|---|---|---|---|---|---|---|
| mtlD (+) nptII (+) |
mtlD (+) nptII (-) |
mtlD (-) nptII (+) |
mtlD (-) nptII (-) |
|||
| CoT 3 | 70 | 46 | 17 | 0 | 7 | 24 |
| CoT 5 | 100 | 80 | 18 | 2 | 0 | 18 |
| CoT 6 | 64 | 34 | 14 | 13 | 3 | 22 |
| CoT 9 | 60 | 31 | 12 | 1 | 16 | 20 |
| CoT 11 | 66 | 50 | 2 | 10 | 4 | 03 |
Table 1: PCR analysis for segregation of the transgenes in T1 progeny of primary co-transformants to obtain marker-free transgenic lines.
Analysis of transgene copy number and expression
The PCR positive marker-free T1 transgenic progenies of co-transformants lines were further employed for confirmation of the integration, as well as for determination of the copy number of the transgene by Southern analysis. Only CoT6 transgenic line displayed single copy insertion, whereas CoT11 and CoT 17 displayed double copy and CoT3 and CoT5 displayed multiple copy integration (Figure 3a). The same marker-free T1 progeny of each CoT transgenic lines were also analyzed with nptII gene (marker gene) probe. As these lines are marker-free as evident by PCR results, none of them showed any signals, further substantiating the absence of marker gene in these transgenics (Figure 3b).
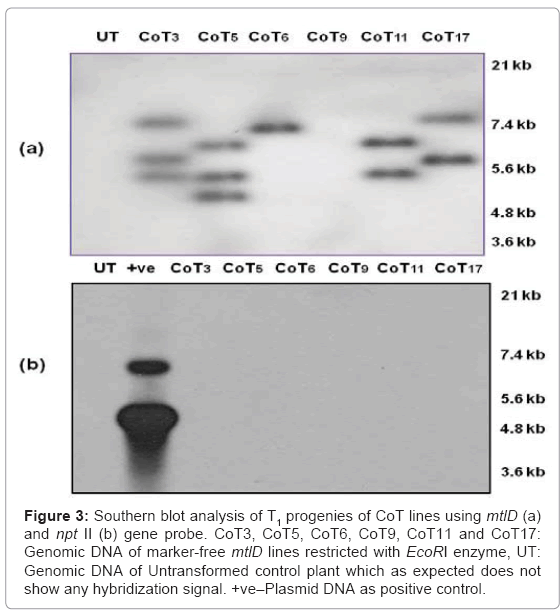
Figure 3: Southern blot analysis of T1 progenies of CoT lines using mtlD (a) and npt II (b) gene probe. CoT3, CoT5, CoT6, CoT9, CoT11 and CoT17: Genomic DNA of marker-free mtlD lines restricted with EcoRI enzyme, UT: Genomic DNA of Untransformed control plant which as expected does not show any hybridization signal. +ve–Plasmid DNA as positive control.
To assess the expression of mtlD gene, the transgenics were subjected to semi-quantitative RT-PCR and northern hybridization. In case of semi-quantitative RT-PCR, CoT3 and CoT11 transcript levels appear to be more or less similar, but higher than the CoT6, which showed low titers of transcripts (Figure 4a). Northern analysis revealed the presence of mtlD transcripts in all the CoT lines, whereas no transcripts were detected in the UT. The co-transformants showed variation in the transgene expression (Figure 4b), and more or less followed the same trend, as in case of RT-PCR analysis.
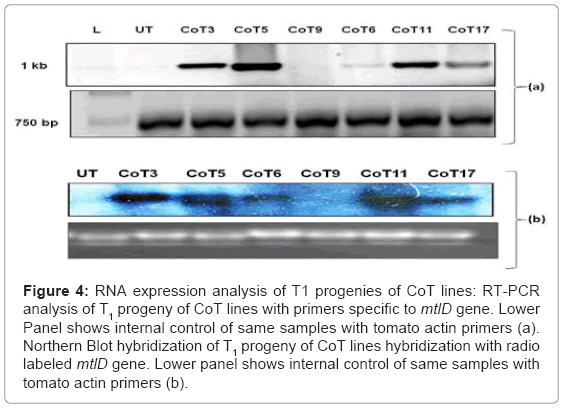
Figure 4: RNA expression analysis of T1 progenies of CoT lines: RT-PCR analysis of T1 progeny of CoT lines with primers specific to mtlD gene. Lower Panel shows internal control of same samples with tomato actin primers (a). Northern Blot hybridization of T1 progeny of CoT lines hybridization with radio labeled mtlD gene. Lower panel shows internal control of same samples with tomato actin primers (b).
Mannitol content
Higher mannitol content was found in all the CoT transgenic lines (CoT3, CoT6 and CoT11), as compared to untransformed control plants, which exhibited no mannitol under normal as well as in salt-stressed conditions. Under unstressed condition, the amount of mannitol in CoT6, CoT11 and CoT3 was about 2.0, 2.8 and 3.0 μmol gm-1 dry weight, whereas mannitol levels were not detectable in the UT plants. After 5-6 days of salt stress treatment with 1/10th MS nutrient solution containing 200 mM of NaCl, the mannitol level in the leaves of all CoT lines increased up to 29-39%. After salt stress, the mannitol levels in CoT6, CoT11 and CoT3 were found to be 2.4, 3.9 and 4.0 μmol gm-1 dry weight, respectively, and were much higher in comparison to unstressed condition, and was found to be statistically significant (Figure 5).
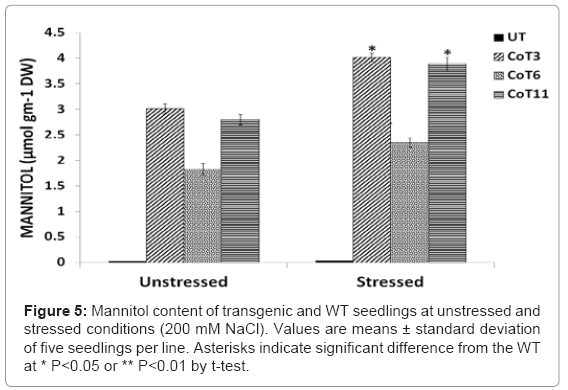
Figure 5: Mannitol content of transgenic and WT seedlings at unstressed and stressed conditions (200 mM NaCl). Values are means ± standard deviation of five seedlings per line. Asterisks indicate significant difference from the WT at * P<0.05 or ** P<0.01 by t-test.
Testing of transgenic lines for abiotic stress tolerance
In vitro seed germination: Under multiple stress conditions, the seeds from marker-free CoT lines showed a higher percentage of seed germination than the untransformed control (Figure 6). On a medium without stress, there was no noticeable difference in seed germination percentage and seedling growth in UT and CoT lines. At 200 mM NaCl, CoT lines exhibited low percent seed germination with a delay in germination by about 6-8 days, when compared with the medium devoid of NaCl. The percentage of seed germination from CoT lines on salt-amended MS medium ranged from 64-88% on 150 mM and 38-56% on 200 mM NaCl, depending upon the transgenic line tested (Table 2). In drought stress, the transgenic seeds were able to germinate, but with a delay of 2-3 days and the seedlings grew well (Figure 6). The percent germination in the CoT lines varied from 44 to 66% in 15% PEG, but the untransformed control seeds failed to germinate (Figure 6) (Table 2). Under heavy-metal stress, the T2 seeds from the different CoT lines were able to germinate in MS medium containing 1 and 2 mM of cadmium (Figure 6) (Table 2). At 1 mM cadmium, the percent seed germination of the CoT lines varied from 62-86%, and it was further reduced with the increasing concentration of cadmium i.e. 48-60% at 2 mM cadmium (Table 2).
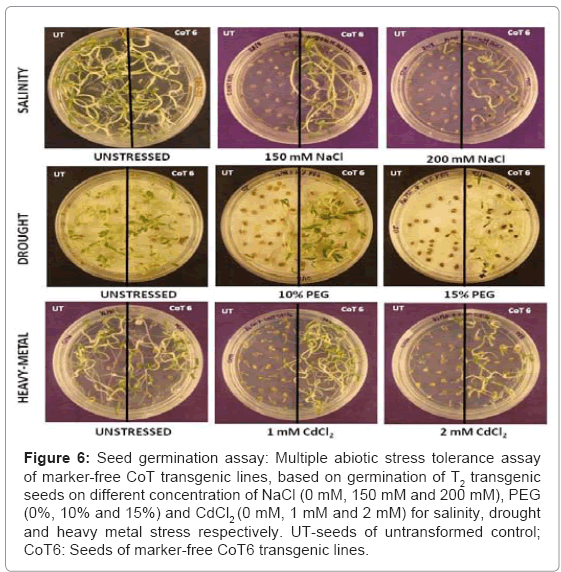
Figure 6: Seed germination assay: Multiple abiotic stress tolerance assay of marker-free CoT transgenic lines, based on germination of T2 transgenic seeds on different concentration of NaCl (0 mM, 150 mM and 200 mM), PEG (0%, 10% and 15%) and CdCl2 (0 mM, 1 mM and 2 mM) for salinity, drought and heavy metal stress respectively. UT-seeds of untransformed control; CoT6: Seeds of marker-free CoT6 transgenic lines.
| SALINITY | |||||||
|---|---|---|---|---|---|---|---|
| 0 mM NaCl | 150 Mm NaCl | 200 mM NaCl | |||||
| Co-transformants lines | % Seed germination | No. of seeds inoculated | No. of seeds germinated | % Seed germination | No. of Seeds inoculated | No. of seeds germinated | % Seed germination |
| UT | 90 | 50 | 0 | 0 | 50 | 0 | 0 |
| CoT 3 | 94 | 50 | 44 | 88 | 50 | 28 | 56 |
| CoT 6 | 92 | 50 | 32 | 64 | 50 | 19 | 38 |
| CoT 11 | 92 | 50 | 40 | 80 | 50 | 26 | 52 |
| DROUGHT | |||||||
| 0 % PEG | 10% PEG | 15% PEG | |||||
| UT | 98 | 50 | 2 | 4 | 50 | 0 | 0 |
| CoT 3 | 97 | 50 | 44 | 88 | 50 | 33 | 66 |
| CoT 6 | 97 | 50 | 32 | 64 | 50 | 22 | 44 |
| CoT 11 | 98 | 50 | 42 | 84 | 50 | 30 | 60 |
| HEAVY-METAL | |||||||
| 0 mM CdCl2 | 1 mM CdCl2 | 2 mM CdCl2 | |||||
| UT | 98 | 50 | 2 | 4 | 50 | 0 | 0 |
| CoT 3 | 98 | 50 | 43 | 86 | 50 | 30 | 60 |
| CoT 6 | 97 | 50 | 31 | 62 | 50 | 24 | 48 |
| CoT 11 | 96 | 50 | 41 | 82 | 50 | 30 | 60 |
Table 2: Percent seed germination based on the germination of T2 seeds from marker-free mtlD tomato co-transformant lines and untransformed control (UT) on MS medium containing different concentration of NaCl (0, 150 and 200 mM), PEG (0, 10 and 15%) and CdCl2 (0, 1 and 2 mM), for salinity, drought and heavy-metal stress.
In vitro seedling growth: Under multiple abiotic stresses, the marker-free CoT seedlings were green and exhibited a low degree of necrosis (Figure 7). All the tested transgenic lines survived stress conditions, though they did suffer some growth penalty. On the contrary, the UT seedlings in 200 mM NaCl showed extensive necrosis and died within 7-10 days, showing that this concentration is lethal for UT seedlings (Figure 7). Enhanced growth was reflected by a significant increase in seedling height in CoT transgenic lines (49-72%) over the UT. The co-transformed seedlings also contained higher fresh and dry weight than the UT control seedlings, which was increased by 37-68% and 47-83%, respectively (Table 3). Overall, CoT11 responded best among the marker-free CoT lines. Similarly, CoT3 performed consistently well among all the CoT lines.

Figure 7: In vitro salt tolerance assay. 1/10th MS+200 mM NaCl (a) 1/10th MS+15% PEG (b) 1/10th MS+2 mM CdCl2 (c). UT-untransformed control; CoT 3, CoT6, CoT11 and CoT12-marker-free transgenic lines generated by Co- transformation.
| SALINITY (200 mM NaCl) | ||||||
|---|---|---|---|---|---|---|
| Seedling height (cm) | Fresh weight (gm) | Dry weight (gm) | ||||
| Lines | Unstressed | Stressed | Unstressed | Stressed | Unstressed | Stressed |
| UT | 17.58 ± 1.26 | 6.97 ± 0.42 (100) | 0.155 ± 0.028 | 0.057 ± 0.005 (100) | 0.0274 ± 0.0021 | 0.0060 ± 0.0005 (100) |
| CoT 3 | 16.32 ± 0.45 | 11.18 ± 0.52* (160) | 0.151 ± 0.012 | 0.096 ± 0.004* (168) | 0.0224 ± 0.0027 | 0.0102 ± 0.0005* (170) |
| CoT 6 | 17.46 ± 0.71 | 10.36 ± 0.49* (149) | 0.150 ± 0.015 | 0.078 ± 0.001* (137) | 0.0200 ± 0.0021 | 0.0088 ± 0.0003* (147) |
| CoT 11 | 17.31 ± 0.63 | 11.99± 0.46* (172) | 0.153 ± 0.006 | 0.088± 0.003* (154) | 0.0238 ± 0.0010 | 0.0110 ± 0.0004* (183) |
| DROUGHT (15% PEG) | ||||||
| UT | 17.44 ± 1.01 | 7.09 ± 0.39 (100) | 0.160 ± 0.030 | 0.059 ± 0.003 (100) | 0.0236 ± 0.0023 | 0.0070 ± 0.0008 (100) |
| CoT 3 | 15.66 ± 0.73 | 9.24 ± 0.50* (130) | 0.158 ± 0.005 | 0.098 ± 0.020 (166) | 0.0220 ± 0.0007 | 0.0120 ± 0.0015* (171) |
| CoT 6 | 15.70 ± 0.24 | 8.80 ± 0.25* (124) | 0.150 ± 0.017 | 0.088 ± 0.013 (149) | 0.0208 ± 0.0012 | 0.0098 ± 0.0009* (140) |
| CoT 11 | 16.36 ± 0.24 | 10.22 ± 0.90* (144) | 0.158 ± 0.017 | 0.125 ± 0.008* (212) | 0.0236 ± 0.0012 | 0.0132 ± 0.0011* (186) |
| HEAVY-METAL ( 2 mM CdCl2) | ||||||
| UT | 17.12 ±1.22 | 6.38 ± 0.32 (100) | 0.162 ± 0.038 | 0.058 ± 0.004 (100) | 0.0254 ± 0.0025 | 0.0057 ± 0.0004* (100) |
| CoT 3 | 16.22 ± 0.72 | 10.3 ± 0.45 * (162) | 0.156 ± 0.004 | 0.094 ± 0.022* (162) | 0.0236 ± 0.0009 | 0.0104 ± 0.0010 (182) |
| CoT 6 | 16.50 ± 0.24 | 8.20 ± 0.85 * (129) | 0.145 ± 0.013 | 0.081 ± 0.013* (140) | 0.0230 ± 0.0011 | 0.0090 ± 0.0009* (158) |
| CoT 11 | 16.61 ± 0.23 | 11.22 ± 0.25* (176) | 0.160 ± 0.018 | 0.098 ± 0.008 * (169) | 0.0248 ± 0.0021 | 0.0113 ± 0.0012 (198) |
Values represent the mean ± SEM, and the data was recorded after a period of one month. *Significant differences from control (stressed) at 5% level. Percent control values are in parenthesis.
Table 3: Seedling growth performance of T2 seedlings of marker-free mtlD tomato co-transformants and untransformed control under salt, drought and heavy-metal stress.
Also, clear growth differences were apparent when the seedlings were subjected to drought stress. During stress treatment, the transgenic seedlings remained green and healthy, whereas there was inhibition of growth and visible damage, such as wilting, bleaching of chlorophyll and total collapse in UT seedlings, which ultimately led to their death. In the presence of PEG, the marker-free CoT seedlings displayed better growth as compared to the UT seedlings. For instance, the co-transformed lines showed a 24-44% increase in the height and 49-112% increase in the fresh weight in comparison to the respective UT. Similarly, about 40-71% increase in dry weight was observed in the CoT seedlings, in comparison to the untransformed control seedlings (Table 3). However, there was some growth penalty in co-transformed lines, as compared to the unstressed seedlings.
Under heavy-metal stress of 2 mM CdCl2, seedlings of the transgenic CoT line remained green and healthy, whereas visual symptoms of chronic cadmium phytotoxicity, including growth reduction, interveinal foliar chlorosis, wilted leaves and necrotic leaf tips were observed in UT seedlings, which resulted in major growth inhibition. The roots of UT seedlings were the most affected and turned brown after few days (Figure 7). There was also increase in height, fresh and dry weight of co-transformants than the control plants. The co-transformants showed 29-76% increase in the height, 40-69% increase in the fresh weight and 58-98% increase in dry weight, in comparison to the untransformed control seedlings (Table 3). Cadmium is considered to be one of the most harmful heavy-metal and induces the accumulation of reactive oxygen species [7,57].
In vivo seedling growth: Under salt stress (200 mM NaCl), the lower older leaves of seedlings of co-transformants became chlorotic and withered, while the younger leaves remained green and healthy. The transgenic lines showed better growth and could withstand salt stress. On the other hand, the UT control stopped growing, and the leaves turned yellow on seventh to eighth day under the same level of salt stress, and the whole untransformed control turned yellow and necrotic within two weeks and eventually died (Figure 8).

Figure 8: In vivo salt tolerance assay. UT-untransformed control; CoT6, & CoT11–Marker-free transgenic lines.
During drought stress (15% PEG), visible differences were evident between the UT control and the co-transformants (Figure 8). The transgenic seedlings exhibited delay in appearance of stress symptoms, in comparison to the UT seedlings. PEG (15%) resulted in leaf drying and wilting in the UT seedlings by fourth to fifth day, while in the transgenic seedlings, these symptoms were delayed until the eighth day. During recovery, the transgenic seedlings responded well and could recover after normal watering (rehydration). They displayed vigorous growth and survived. Conversely, the UT control seedlings could not revive (Figure 8).
The tolerance of the co-transformants to cadmium-induced heavy-metal stress was tested under in vivo growth conditions by watering with 2 mM cadmium solution, the transgenic lines showed better growth than the untransformed control, which displayed reduced growth and “burned” leaves, and subsequently they died (Figure 8).
Chlorophyll content: Under unstressed conditions the chlorophyll content in CoT lines and UT remained same. But on application of different stresses, the drastic differences of chlorophyll content were found in the UT and CoT lines. Under stress conditions, chlorophyll content of CoT was quite high and remained stable than the UT (Figure 9). This showed that CoT seedlings with higher accumulation of mannitol maintained relative steady content of chlorophyll under multiple abiotic stresses.
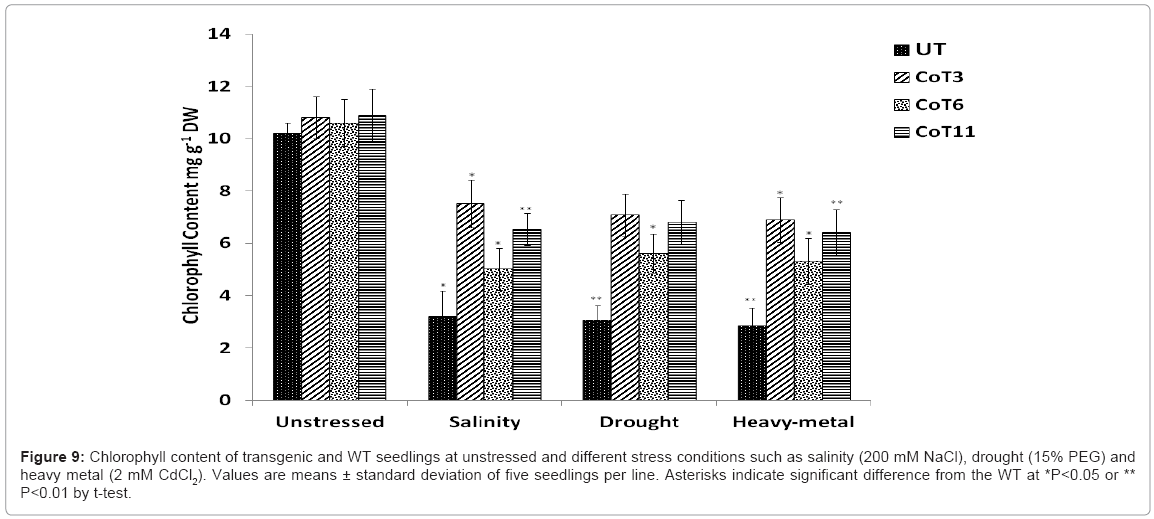
Figure 9: Chlorophyll content of transgenic and WT seedlings at unstressed and different stress conditions such as salinity (200 mM NaCl), drought (15% PEG) and heavy metal (2 mM CdCl2). Values are means ± standard deviation of five seedlings per line. Asterisks indicate significant difference from the WT at *P<0.05 or ** P<0.01 by t-test.
Photosynthetic efficiency: The Fv/Fm ratios in UT were significantly reduced during stress, while the ratios were not dramatically decreased and remained stable in marker-free CoT lines (Figure 10a). On 5th day of stress, Fv/Fm ratio was reduced up to 43-57% in UT, but only 10-20% in CoT lines. Moreover, the Fv/Fm ratios in CoT3 and CoT11 remained high than CoT6 on stress treatment (Figure 10a). Under different stresses, ETR was also reduced drastically in the UT, in comparison to CoT lines on 5th day. The reduction was up to 42-50% in UT, but only 8-10% in CoT lines (Figure 10b). Photosynthetic yield, Y(II) of CoT lines was also increased up to 50-60% in comparison to UT control (Figure 10c). Interestingly, there was increase in the value of NPQ of UT in comparison to CoT lines. An increase in NPQ suggested that abiotic stress treatment induced dissipation of damaging excess energy (Figure 10d).
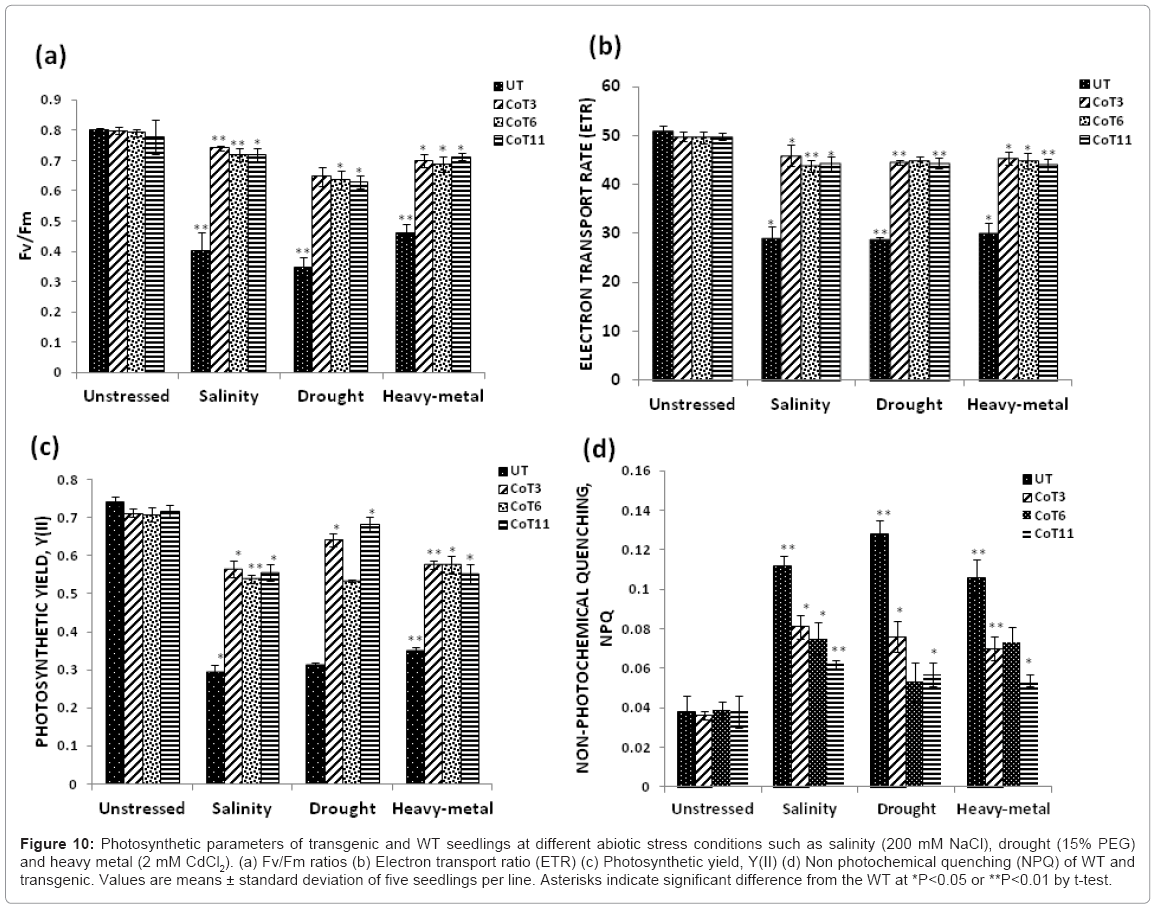
Figure 10: Photosynthetic parameters of transgenic and WT seedlings at different abiotic stress conditions such as salinity (200 mM NaCl), drought (15% PEG) and heavy metal (2 mM CdCl2). (a) Fv/Fm ratios (b) Electron transport ratio (ETR) (c) Photosynthetic yield, Y(II) (d) Non photochemical quenching (NPQ) of WT and transgenic. Values are means ± standard deviation of five seedlings per line. Asterisks indicate significant difference from the WT at *P<0.05 or **P<0.01 by t-test.
Our main objective was to generate marker-free transgenic tomato plants against multiple abiotic stresses via co-transformation strategy, by the introduction of mtlD gene for mannitol accumulation. There are few reports in which different scientific groups had demonstrated that cellular accumulation of mannitol can confer abiotic stress tolerance [14,16,26-28]. We have successfully used co-transformation strategy using mixed Agrobacterium cultures to obtain marker free transgenics with high co-transformation and segregation frequency. Our co-transformation frequency is relatively high (up to 24%), compared to the previous reports, where only 2-4% of co-transformation frequency was reported [41,58]. The segregation frequency to obtain marker-free transgenic is also quite high, ranging from 20-24% in CoT3, CoT6 and CoT9. The lower segregation frequency in some lines such as CoT11 was observed, that may contribute to the reason that the two separate T-DNAs integrate with a higher frequency at linked loci. Also, multi-copies of transgene in the transgenic plants may be one of the other reasons for the low segregation frequency of the target and marker genes. Komari et al. [58] reported that the frequency of co-transformation of the target gene and SMG was 2%, while the frequency of segregation of the two genes was 100% when two binary vectors were used separately (1:1 mixture). In another study, a higher frequency of co-transformation (82-90%, and lower efficiency of segregation (14.3-17.4%) of two genes was also observed when the twin T-DNA binary vector system was employed [59]. It was found to be technically simple and an effective approach for segregating the transgene and the marker gene, and for subsequent marker removal. This strategy was successfully used by other groups with low co-transformation frequency, but high segregation frequency [41-43,58,59].
We have successfully raised many mtlD gene overexpressing marker-free transgenics of tomato by Agrobacterium-mediated transformation protocol standardized in our lab [51]. Transgene integration and copy number was confirmed by PCR and Southern blot analysis, respectively, whereas its expression was confirmed by RT-PCR and Northern blotting. In case of semi-quantitative RT-PCR, there is a variation in the transcript level of transgenics. CoT3 and CoT11 transcript levels appear to be more or less similar, but higher than the CoT6 which showed low titers of transcripts (Figure 4a). This may be due to the single copy of transgene in CoT6, as compared to CoT3 and CoT11 lines which have multiple transgene insertions. CoT5 showed highest level of transcripts, and it has multiple copies of T-DNA insertions (Figure 4a). In Northern blotting, co-transformants also showed variation in the transgene expression (Figure 4b), and more or less followed the same trend, as in case of RT-PCR analysis. The disparity in the transcript levels among the double copy lines (CoT11 and CoT17), as well as with the multiple copy lines (CoT3 and CoT5), may be attributed to the positional effect. It is well known that the transgene expression can be strongly influenced by site of integration, which can lead to low or high level of transgene expression [60,61]. Moreover, variabilities in the level of gene expression from independent transformants are very common in plant transformation systems [62]. Certain other factors, such as transgene copy number and construct fidelity of the transgene, also influence transgene expression [60,63]. Under normal condition, high expressing line CoT3 and CoT11 accumulated higher amounts of mannitol than CoT6 line, which was low expressing line, indicating a positive correlation between mtlD transcript level and mannitol accumulation. Although mtlD gene is driven by constitutive promoter (CaMV 35S), higher mannitol accumulation was seen under salt stress condition, suggesting that CaMV 35S promoter might be responding to salt stress.
Osmotic stress mediated by salinity, drought and heavy-metal conditions in tomato primarily causes ionic and metabolic imbalance, thus disrupting cell homeostasis [57,64,65]. Hence, the integrity of cellular membranes, the activities of various enzymes and function of photosynthetic apparatus are all known to be prone to the toxic effects of stress causing molecular damage, growth arrest and even cell death [13,57,66]. Marker-free CoT lines performed better in all the stresses, but showed variation in tolerance level. Under abiotic stress conditions, CoT3 and CoT11 showed higher tolerance than the CoT6 line. This could be due to higher accumulation of mannitol in these transgenics, as they are high expressing lines, whereas CoT6 was a low expressing line. Comparison between the tolerance levels of CoT lines suggested that these differences could be due to the positional effects mediated variations are more critical for the transgene expression than the transgene copy number [67]. In addition, multiple transgene insertions are often associated with gene silencing, which could also possibly have affected its mannitol levels and consequently, the tolerance pattern.
Abiotic stress indirectly inhibits the photosynthetic activity of plants and thus, reduces its productivity [68,69]. The decrease in photosynthesis induced by abiotic stress can be associated with partial stomatal closure and reduced chlorophyll content. The survival of the marker-free CoT seedlings under stress may be due to the retention of photosynthetic capacity, as a result of increased levels of mannitol. Under stress conditions, chlorophyll content of CoT was quite high and remained stable than the UT (Figure 9). This meant that CoT seedlings with higher accumulation of mannitol maintained relative steady content of chlorophyll under multiple abiotic stresses, and thus, increases the photosynthetic capacity of transgenics [14,16,29,30]. The Fv/Fm ratio is the most frequently used parameter to indicate injury to the PSII complexes due to abiotic stress factors, including salinity, drought and heavy-metal [70]. The Fv/Fm ratios in UT were significantly reduced during stress, while the ratios remained stable in marker-free CoT lines (Figure 10a). On 5th day of stress, Fv/Fm ratio was reduced up to 43-57% in UT, but only 10-20% in CoT transgenics. Moreover, the Fv/Fm ratios in CoT3 and CoT11 remained high than CoT6 on stress treatment, indicating that the PSII complexes had not suffered serious injury due to high accumulation of mannitol during stress treatment (Figure 10a). This suggests the protective effect of mannitol on Photosystem II activity, and cell membrane integrity. Under different stresses, ETR was also reduced drastically in the UT, in comparison to CoT lines on 5th day. The reduction was up to 42-50% in UT, but only 8-10% in CoT lines (Figure 10b). ETR provides a good way to evaluate the photosynthetic performance in stressed plants [71], and to gain insight into the behavior of the photosynthetic machinery under stress [70]. Photosynthetic yield, Y(II) of CoT lines were also increased up to 50-60%, in comparison to UT control (Figure 10c). Interestingly, there was increase in the value of NPQ of UT, in comparison to CoT lines. An increase in NPQ suggested that abiotic stress treatment induced dissipation of damaging excess energy (Figure 10d). This energy could be dissipated at the antennae through heat emission, although other mechanisms involved in energy dissipation related to a transmembrane H+ gradient generated by ATPase activity and uncoupling of electron transport, that may lead to oxidative stress, could also be involved [70,72].
However, having potential to act as free-radical scavengers, mannitol can protect the membranes and other macro-molecules from oxidative damages, and thus, can stabilize biological membranes under stressful conditions [16,54]. Due to its free-radical scavenging properties, mannitol accumulation also provide tolerance against heavy metal, such as cadmium, which is considered to be one of the most harmful heavy-metal and induces the accumulation of reactive oxygen species [7,57].
In conclusion, expression of the mtlD gene in marker-free transgenic tomato plants resulted in higher levels of mannitol accumulation and substantial protection of the vigour of the plants under multiple abiotic stress conditions. Mannitol not only improved tolerance, but also increased the photosynthetic efficiency, suggesting that carbohydrate alterations produced by mannitol biosynthesis be linked to the stress response. Our results would provide a helpful tool for understanding the physiological function of mannitol under abiotic stress in tomato plant.
This work is supported by Defense Research Development Organization (DRDO), New Delhi, India. We thank Prof. Hans J. Bohnert, University of Arizona, USA for providing E. coli mtlD gene. The pCAMBIA 1380 was kindly provided by Dr. R. Jefferson, Centre for Application of Molecular Biology in Agriculture (CAMBIA), Australia. The Agrobacterium strain LBA 4404 and the binary plasmid pBinAR were obtained from Dr. P. Anand Kumar, Indian Agricultural Research Institute (IARI), New Delhi. Brijesh Gupta is thankful to the University Grant Commission (UGC), Government of India for the award of senior research fellowship. We also thank UGC, New Delhi for special assistance programme and Department of Science and Technology, New Delhi for FIST programme and Ms. Sneha Yogindran for her help.