Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2014) Volume 2, Issue 1
Sex outcome and maternal investment in progeny are important predictors of reproductive success. While environmental factors appear to influence these processes, there is little evidence to date of a direct role for gut commensals. Here we show that the reproductive outcomes (sex and survival) of mouse litters depend on signals conveyed through the microbiome. We discover that transient treatment of mouse mothers with specific microorganisms increases the absolute survival of offspring and skews offspring sex ratios via an endocrine-dependent mechanism requiring the neurophysiological hormone oxytocin. The implication of maternal oxytocin levels suggests that commensal microbes may have a broad role in modulating host endocrine and neurological pathways.
Keywords: Lactobacillus reuteri; Oxytocin; Sex ratio
Factors governing important predictors of reproductive success, such as maternal investment in progeny, remain poorly understood [1-4]. The great importance of gut microbiota in mammalian host health is only recently being recognized in full [5-8]. It is known that consumption of a Westernized diet can alter the microbiota in mice [9-12] commensurate with infanticide, leading us to postulate a microbe-dependent mechanism of action in maternal investment. To test this hypothesis, we surveyed C57BL/6 mice to determine whether there were changes in the survival of offspring and sex ratio associated with targeted consumption of a specific microorganism.
Lactobacillus reuteri (LR) is widely applied as a model beneficial microbe in humans and animal models [13-18]. During a recent study we observed that female mice consuming LR showed more frequent grooming activity, an aspect of maternal behavior regulated by neurohypophyseal hormone oxytocin [19-21]. Intriguing roles for oxytocin in diverse aspects of health have recently emerged [22-26]. Mechanisms by which gut microbiota may impart effects that expand beyond the gastrointestinal tract remain largely unknown but involve neuroendocrine and immune pathways [19,27-33]. When consuming LR, mice also display sex-specific phenotypes in skin and vaginal pH and immune response likely to have consequences on reproductive success [34]. Evidence for sex-specific effects on pH levels, oxytocin and immune response in mice fed LR gave a foundation for these studies [34].
Microbe-dependent changes in maternal care and sex ratio likely arise from modified endocrine signaling [1,32,35-41]. Here, we investigate whether microbe-dependent hormone changes alter sex ratios. In previous studies, we found significant upregulation of the neurohypophysial hormone oxytocin in female mice consuming LR [34,37]. Oxytocin plays a central role in mammalian reproductive and social behaviors, and is produced in high quantities during birth. Administration of oxytocin induces reproductive behaviors and produces neuronal activity observed during mating. Oxytocin-deficient mice exhibit social amnesia with reduced bonding and social behavior [21]. Recent evidence suggests a role for oxytocin in autism spectrum disorder [42] and post-partum depression, as well as pair bonding, anxiety, and maternal care [43-47]. Because these and other social and reproductive disorders implicate oxytocin, there is great interest in assessing whether an underlying microbial determinant may be modifying the central nervous system vis-à-vis direct or indirect influence on the activity of oxytocin in the body. Interestingly, oral therapy of B. fragilis alleviates stereotypic autism-like behaviors in a mouse model known to display features of autism, reinforcing the potential for microbial control of neurological disorders [41]. To test whether gut microbe influence over oxytocin played a role in the sex ratio outcomes observed here, we used Oxytocin-Knockout (KO) mouse mothers to assess whether sex ratios differed in offspring of these mice compared to matched controls. Our results suggest that the distribution of sex ratios in mice differs depending on maternal gut microbes that influence these outcomes through a neurophysiological and, in particular, oxytocin-dependent mechanism. The centrality of oxytocin in these outcomes suggests potential associations with autism and post-partum depression [42,43], offering promise for gut microbe therapy in these and related disorders.
Animals
Wild type C57BL/6J and oxytocin-deficient B6.129S-Oxttm1Wsy/J mice (Jackson Lab; Bar Harbor, ME) were housed and handled in Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)-accredited facilities with diets, experimental methods, and housing as specifically approved by the MIT Institutional Animal Care and Use Committee (MIT IACUC). Treatment groups included five-ten mother mice per group examining three consecutive litters of their progeny.
Experimental design: Eight-week-old C57BL/6 wild type female mice were fed special diets as above, and paired three weeks later using one male and one female per cage. Mating pairs consumed NWD (N=6), control diet (N=6), and control diet + LR (N=9). Mother mice were checked daily to record births. Survival rates and infant sex were determined upon birth and at weaning, as below. Experiments were conducted in duplicate.
Eight-week-old female oxytocin-deficient B6.129S-Oxttm1Wsy/J or littermate wild type mice received supplementary LR in their drinking water, and were then paired three weeks later using one wild type male and one experimental female per cage. Mating pairs consumed a control diet (N=10) or a control diet + LR (N=10). Mother mice were checked daily to record births and infant sex identity. Newborns from oxytocin-deficient moms were cross-fostered to OXT-het mother mice, because OXT-KO mother mice are incapable of lactation and nursing behaviors. Survival rates and sex ratio were determined upon birth and at weaning, as below.
Special diets for animals
Mice of eight-weeks-of-age were placed on experimental diets:
1) control diet AIN-76A (Harlan-Teklad, Madison WI),
2) AIN-76A plus an anti-inflammatory strain of Lactobacillus reuteri ATCC-PTA- 6475 (LR),
3) New Western diet (NWD) high in fat, and low in fiber and nutrients including vitamin D (TD.96096; Harlan-Teklad).
L. reuteri was cultivated as described previously [34,48] using a starting dosage of 3.5 × 105 organisms/mouse/day in drinking water. Live bacterial counts in water bottles were calculated to be 1.4 × 106 colony forming units (CFU) per mouse after 24 hours, 4.1 × 105 CFU at 48 hours, and 1.1×105 CFU at 72 hours, when quantified as described elsewhere [48]. Fresh drinking water was replaced twice weekly throughout the experiments. Control mice received regular drinking water. Confirmations of LR organisms in drinking water and in experimental animals were performed as described previously [48].
Mice on various diets were subsequently arranged in breeding pairs and examined as above.
Determination of survival rate
Infant survival rates were calculated using number of progeny weaned/number of progeny born. Pregnant mice were checked daily.
Determination of sex
Sex was determined upon birth or weaning (21-24 days) using standard anogenital distance measurement from anus to genital opening. Measurements were standardized using animal body weight and length.
Statistical analyses
To determine the relative effects of diet on survival, we chose to apply a binomial regression model, in which the response is one of two mutually exclusive outcomes. Outcomes are binary for both sex (male/female) and survival (live/dead), with dependency on categorical experimental variables. Each cage was considered a replicate, so the response from a single cage was the ratio of surviving offspring to non-surviving offspring (measured at weaning). The model takes the form:
log(p/(1-p))=β0+β1*X
Where p/(1-p) is the odds ratio, p the proportion of surviving offspring, (1-p) the proportion of non-surviving offspring. Each X can be a categorical variable (in this case, diet), and the β1 value gives an estimate of the differences due to these variables. Significance is determined by a Z-test on the parameters of interest using null β1=0 and alternative β1/=0. Statistical tests were performed in R (http://cran.r-project.org). The same model was applied to determine whether the fraction of females differed by diet, where response was the ratio of females to males. Samples were partitioned by litter to control for parental effects.
Maternal probiotic consumption increases the survival rate of offspring
We had previously examined age-associated weight gain in mice consuming a high-fat, low-nutrient New Western Diet (NWD) mimicking ‘fast food’ consumption [9] shown in previous work to alter the composition of the microbiota [10]. In order to test the influence of a Westernized diet on subsequent generations, we fed C57BL/6 mouse mothers NWD starting three weeks prior to mating. We then monitored the sex and physical characteristics of their offspring over three successive litters.
We noted differences in survival between groups of offspring corresponding to diet. Survival for all groups increased with successive litters. We chose to apply a Binomial Logistic Regression (BLR) model with the odds ratio of survival, given that survival is a binary outcome, as the response and diet as the predictor. We found that the offspring of NWD exhibited significantly different odds of survival from their control counterparts across all litters, even when incorporating the observation of increased survival in successive litters. The log(odds) ratio for survival of offspring from NWD-fed mice was -1.04 ± 0.41 (±s.e., d.f.=26, Z=-2.52, p=0.0012). The 95% confidence interval for the ratio of surviving offspring in NWD compared to control was (0.233,0.533), indicating a 2-5 fold decrease in survival in NWD-fed mice. Infant progeny of mothers fed control diet exhibited increased survival over mice eating NWD, indicating that the stress diet appears to influence negatively the probability of offspring surviving past weaning (Figure 1). Given that consumption of NWD is known to alter the composition of the microbiota [10], we hypothesized the changing microbial presence may have a role in differential survival between groups independent of other diet-dependent variables.
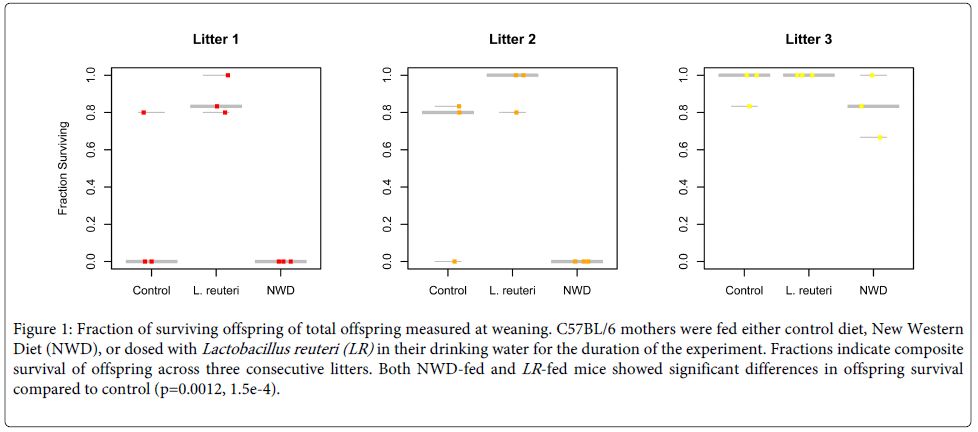
Figure 1: Fraction of surviving offspring of total offspring measured at weaning. C57BL/6 mothers were fed either control diet, New Western Diet (NWD), or dosed with Lactobacillus reuteri (LR) in their drinking water for the duration of the experiment. Fractions indicate composite survival of offspring across three consecutive litters. Both NWD-fed and LR -fed mice showed significant differences in offspring survival compared to control (p=0.0012, 1.5e-4).
To investigate the role of gut commensal microorganisms in survival of offspring in mouse litters, we made a targeted intervention through feeding mouse mothers a single probiotic microorganism. In previous work, we found that mice consuming Lactobacillus reuteri (LR) in drinking water display sex-specific phenotypes in skin and vaginal pH and immune response that are likely to have consequences on reproductive success [3]. We investigated whether transient administration of LR to drinking water could affect changes in reproductive outcomes in mouse mothers, and thus be indicative of a microbe-dependent mechanism. To test this hypothesis, we administered 3e6 CFU/mL LR into the drinking water of C57BL/6 mouse mothers (corresponding to approximately 1e7 CFU dosage/day assuming no loss in cell viability) every 7 days with feeding of a nutritionally balanced control diet. We then surveyed mouse births and rearing to determine whether there were changes in the survival of offspring with less maternal neglect or cannibalism associated with targeted consumption of this probiotic microorganism.
We found a significant increase in offspring survival for mouse mothers fed LR. In contrast to the reproductive outcomes for the mice fed NWD, the log (odds) ratio for survival of offspring from L. reuteri fed mice was 2.50±0.66 (±s.e., d.f.=26, Z=3.78, p=1.5e-4). The corresponding 95% confidence interval for the odds ratio (3.36, 44.4) indicated an increase in survival compared to the control (Figure 1). This result indicates that supplementation with LR alone increased the number of surviving offspring, supporting the hypothesis of a microbial determinant for this reproductive phenotype.
Maternal consumption of probiotic bacteria influences the sex ratio of offspring
Because maternal exposures may have a role in offspring sex ratio [1], we next tested whether there were differences in the sex outcomes of mice dependent on whether their mother had consumed LR or not. Evidence for sex-specific effects on pH levels and immune response in mice fed LR further supported this hypothesis [34]. We tested whether there were changes in sex ratio associated with consumption of LR in C57BL/6 mice. Examining the number of males and females at birth and subsequently at weaning, we tested for the effect of consuming LR on offspring sex ratios again using BLR with ratio of females to males as the response to diet. Because sex ratio outcomes varied across litters, we analyzed each litter separately in addition to assessing the overall population (Figure 2). In the first litter, there was no significant difference in sex odds in the offspring of mice fed L. reuteri (d.f.=7, Z=1.12, p=0.26), but in the second litter, the effect was significant with the offspring of L. reuteri-fed mice exhibiting a log(odds) ratio of 1.69 ± 0.61 (±s.e., d.f.=9, Z=2.61, p=0.0091), with 95% confidence interval for odds (2.75, 16.47). This effect was not as significant in the third litter (d.f.=10, Z=1.82, p=0.068). Across all three litters, the effect of L. reuteri on sex outcomes was significant, with log (odds) ratio 1.11 ± 0.35 (± s.e., d.f.=28, Z=3.16, p=0.0016) and 95% confidence interval for odds (2.14, 4.32) (Figure 2).
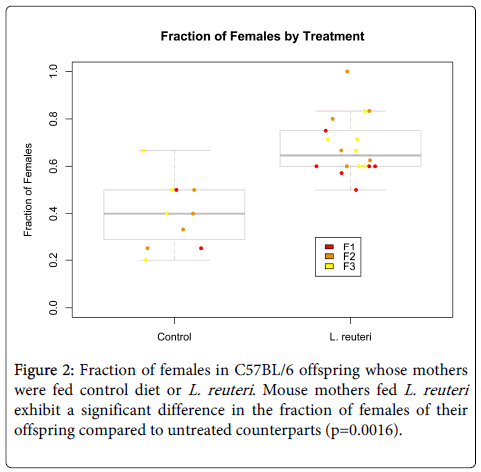
Figure 2: Fraction of females in C57BL/6 offspring whose mothers were fed control diet or L. reuteri. Mouse mothers fed L. reuteri exhibit a significant difference in the fraction of females of their offspring compared to untreated counterparts (p=0.0016).
Because the effects on sex ratio arise even after treatment with a single microbe we inferred the existence of a microbe-mediated alteration in sex outcomes.
Maternal microbial effects on infant sex ratio require the hormone oxytocin
We hypothesized that the observed changes in sex ratio and offspring survival may depend on an endocrine-mediated mechanism in the maternal host. In mammalian hosts, maternal nurturing behaviors require the production of oxytocin. In our previous studies, we found significant upregulation of oxytocin in female mice consuming LR [34,37]. We used 129 strain Oxytocin-Knockout (OXT-KO) mouse mothers and their wildtype littermate mothers to determine if sex outcomes differed because of the absence of oxytocin.
Because the offspring of OXT-KO mice must be fostered onto competent mothers, we measured sex ratio outcomes at birth rather than at weaning. To measure differences in offspring sex ratio with both genotype and diet as factors, we constructed a binomial logistic regression including these factors and their interaction. Genotype alone had a significant effect on offspring sex ratios (BLR, df=85, |Z|=3.085, p=0.002) as well as the interaction of genotype and treatment (BLR, df=85, |Z|=2.316, p=0.0205). These results indicate that oxytocin deficiency tends to decrease the overall ratio of females to males in the offspring, while treatment across groups did not result in significantly different outcomes (Figure 3).
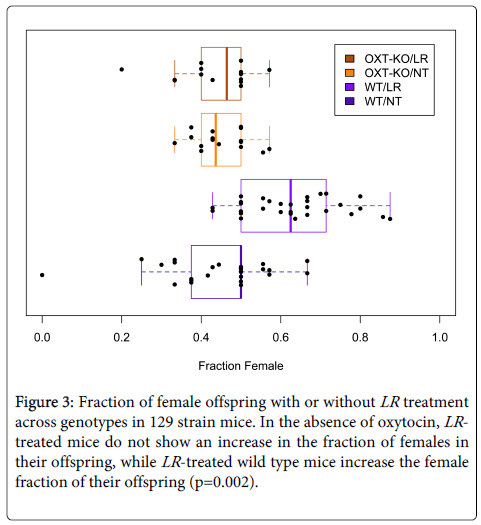
Figure 3: Fraction of female offspring with or without LR treatment across genotypes in 129 strain mice. In the absence of oxytocin, LR - treated mice do not show an increase in the fraction of females in their offspring, while LR -treated wild type mice increase the female fraction of their offspring (p=0.002).
Comparing the LR-treated animals to untreated animals mutes the important interaction of genotype with treatment, where we see significant differences between the sex ratios of LR-treated wild type mice and LR-treated oxytocin deficient mice. Under the null hypothesis that microbe-mediated maternal effects on sex ratio do not require oxytocin, we expect that there should be no differences between the outcomes of OXT-KO and WT mice when treated with LR. However, we find that the WT mice exhibit significantly elevated ratio of female to male offspring when compared to OXT-KO mice.
As a consistency check, there are no significant differences in offspring ratio between untreated WT and KO mice (BLR, df=41, |Z|=0.017, p=0.987) nor between knockout mice treated with LR or not (BLR, df=29, |Z|=0.375, p=0.708). In line with our original findings, in these mice there are still strong differences in sex ratio outcomes between LR-treated and untreated wild type mice (BLR, df=54, |Z|=3.58, p=3.46e-4). Taken together, our results give substantive evidence for the importance of oxytocin in mediating microbial control over host offspring sex ratios. That a specific organism modulates control over maternal reproductive outcomes in an oxytocin-dependent manner suggests that these microbes play an intimate role in fitness of their mammalian hosts.
These data support a role for maternal microbes in controlling several features of maternal care for offspring and physical characteristics such as offspring sex ratios. Because the effects on sex ratio arise even after treatment with a single microbe, LR, there appeared to be a microbe-mediated alteration in host factors. We determined that: 1) offspring survival depended on treatment with the probiotic LR, 2) sex outcomes of mice depended on microbial exposures of their mother, 3) microbial effects on sex ratio and offspring survival depended on maternal oxytocin, and 4) maternal microbial exposures regulated the effects of oxytocin. To our knowledge this is the first report of microbes altering mammalian offspring sex ratio, one key factor predicting reproductive success of the microbes and their hosts. We conclude that host organisms develop and evolve both their nature and nurture dependent on their microbial commensals. Thus, the microbiome in concert with its maternal host determines the nature and nurture of her offspring.
We used each dietary treatment to infer that gut commensal microbes altered maternal state in such a way that LR improved and NWD diminished offspring survival. We found that a mouse mother’s consumption of the NWD chow negatively influenced the survival of her offspring. Given that the microbiome undergoes a drastic shift [9-11,45] after feeding of westernized diets, we postulate differences in sex ratio outcomes depended, at least in part, on the specific microbes and of their mother. Indeed, feeding of LR was previously shown to acidify vaginal pH levels in mice, indicating that vaginal pH may serve as a discriminating feature for some of these outcomes [34]. It remains to be shown whether maternal microbiota and vaginal pH levels influence sex ratio and maternal care in human subjects.
We find that gut microbes can manipulate host oxytocin levels to alter successive generations, having shown an up-regulation of oxytocin following administration of LR [34,37] and a maternal microbe-derived, oxytocin-dependent change in offspring sex and survival. Because we observed sex outcomes at birth, we were able to conclude that the sex effect of oxytocin manifested prior to parturition, removing potential biases from social and behavioral impairment related to oxytocin deficiency that would affect maternal care patterns. Implication of oxytocin in these outcomes suggests potential associations with certain neurological disorders including autism [42] and post-partum depression [43,46,47] with an underlying microbial determinant. Taken together, the present data show that a specific organism modulates control over maternal reproductive success in an oxytocin-dependent manner where microbes play an intimate role in the fitness outcomes of their hosts.
Co-evolution between host and microbe provides an integrative mechanism by which mothers may skew the sex ratios of their offspring in such a way to optimize for specific environmental conditions [32,35-41]. Under favorable conditions, probiotic bacteria may impart a bias toward female births, as they are then passed from mother to naïve offspring during vaginal birth and nursing, imparting evolutionary success to both the symbiotic bacteria and their mammalian hosts. In natural populations, there is empirical evidence for the impact of maternal environment on offspring sex ratios [2]. The transmission of environmental information by microbial commensals appears to occur via an oxytocin-dependent mechanism, consistent with findings of other groups on a stress-mediated modulation of sex ratios [2-4]. The intimate bond between a host and its microbiome provide a means by which both may influence the fitness of the other, sometimes manifesting in transgenerational effects.
We thank James Versalovic for the gift of ATCC 6475 Lactobacillus reuteri, and special thanks to James G. Fox for encouragement and support. The work was supported by National Institutes of Health grants P30-ES002109, U01 CA164337, and RO1CA108854. There are no competing financial or commercial interests.