Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2013) Volume 2, Issue 4
To increase detection of a complex chemical environment, vertebrates and insects express an extraordinary number of distinct olfactory receptor neuron (ORN) classes, each functionally specialized to detect a set of odorants. This is achieved as the olfactory system develops and each of these ORN classes makes developmental decisions defining the olfactory receptor genes they will express and their class-specific connections in the brain. In addition to this high level of ORN diversity, olfactory systems are also very dynamic evolutionarily, with both the number and functionality of olfactory receptor genes as well as the requirement for certain ORN circuits being under ecological constraints. In this review, we will discuss molecular and developmental strategies underlying ORN diversity and evolutionary plasticity as well as present the insect olfactory system as a model for evo-devo research in light of recent findings.
<Insects are the most diverse group of organisms on Earth, with over a million different species having been identified so far [1]. They occupy a vast variety of different niches and survive and thrive in a wide variety of different habitats, from the tropics to the arctic, and a few species are even entirely marine [2-4]. Insects are also economically important to humans, playing various roles that range from crop pollinators to agricultural pests to disease vectors. In the same way, the olfactory environment for insects is also richly diverse and species-specific, with every insect responding to a particular set of odorants in a manner that depends on the ecology and natural history of each unique species. In many cases, this can lead to different responses by different species of insects to identical odorants [5]. A good example of this would be carbon dioxide - several species such as termites and bees use carbon dioxide as an indicator of air quality [6], while certain species of moths use carbon dioxide as an indicator of floral profitability when feeding on nectar [7]. In addition, some species of Drosophila fruit flies use carbon dioxide as an repulsive alarm pheromone [8,9], while some species of blood-feeding insects such as mosquitoes and tsetse flies use carbon dioxide as their primary means of locating a host [10,11]. Thus, a single common odorant can elicit widely different behaviors from these different species of insects [11,12]. This phenomenon can even be observed between closely related species, especially for those species that have evolved to become specialist feeders on particular food sources. For example, the fruit fly Drosophila sechellia, which is a specialist feeder on the fruit of Morinda citrifolia, is highly attracted to the hexanoic and octanoic acids that form the primary odorants of the Morinda fruit, whereas its sister species, Drosophila simulans, is repelled by those same odorants and to whom the fruit itself is highly toxic [13]. This example highlights the functional plasticity of the olfactory system in insects and also the speed at which the system can evolve, as D. sechellia and D. simulans diverged only about 400,000 years ago [14]. The insect olfactory system is thus an excellent model system for learning more about the intersection between evolution and development.
The field of evolutionary developmental biology has made rapid advances over the past few decades, illuminating the extent and degree to which developmental processes are conserved (i.e. Hox genes) as well as demonstrating how these processes can change to allow the emergence of novel forms and structures. Nevertheless, much of the focus of this research has been on large dramatic differences over long evolutionary periods (i.e. the origins of major organs such as eyes and limbs) rather than relatively small changes between closely related species over short periods of time [15]. As such, there is still much to learn about how developmental processes can provide the phenotypic variability that enables short-term evolutionary phenomena such as behavioural modification, host specialization and speciation, as well as how genome-environment interactions influence these processes to modulate evolutionary change. Studying the insect olfactory system from an evo-devo perspective allows us to contribute answers to many of those questions. Given the crucial role of olfaction to most aspect of insect behaviour, from mate-seeking and finding food to complex social behaviours, an increased understanding of the differences in the developmental processes underlying these behaviours can greatly inform our understanding of the potential capacity and rate of evolutionary change.
The speed with which the olfactory system evolves is one of the major advantages of using olfactory system to investigate the modulation of developmental processes underlying evolutionary change. Comparative studies of insect genomes have shown that the genes known to be involved in the development and function of the olfactory system evolve much faster than other parts of the genome, with a continuous “birth-and-death” process causing rampant gene duplication and pseudogenization along with strong indicators of positive selection acting across the system [16-18]. This is in line with what is known about the olfactory receptor gene family in vertebrates, where that is good evidence that the olfactory sub-genome evolves quickly in response to changes in the environment as well as the natural history of the species in question. An oft-cited example of this is the massive loss of olfactory genes due to gene pseudogenization in the primate lineage as the visual system replaced the olfactory system in importance [19,20], and there is every reason to suspect that similarly dynamic genome-environment interactions underlie much of the diversity that we see in insect olfactory sub-genomes.
In this paper, we discuss molecular and developmental strategies underlying both ORN diversity and evolutionary plasticity. We also aim to present the insect olfactory system as an excellent and interesting model system for future evo-devo research, and to highlight recent work that has increased our understanding of how changes in development of the olfactory system can affect the trajectory of a species’ behavioral response to a particular odorant over time. In particular, we examine recurring developmental trends in the evolution of insect specialization and the importance of changes in regulatory genes to olfactory development.
The insect olfactory system has a very specific organization that mirrors that of vertebrates: odorants in the air first come into contact with Odorant-Binding Proteins (OBPs) that are responsible for binding with and solubilizing odorants, bringing them into contact with Olfactory Receptors (ORs) that are expressed by olfactory receptor neurons (ORNs) [21]. Cell bodies of ORNs are themselves located within hair-like structures called sensilla that are found on the surface of the two olfactory appendages of the insect, the antennae and the maxillary palps [22]. In Drosophila melanogaster and other insects where the organizational and molecular details of the olfactory system are understood, each ORN expresses only one specific OR, and each sensilla houses 1-4 individual ORNs of a specific type [23,24]. Sensilla are subdivided according to their overall morphology, of which there are three basic types: trichoid (long, thick and straight, with a basal drum structure), basiconic, (short, thick, and rounded) and coeloconic (short and thin, often in a pit-like structure) [25]. Basiconic sensilla can also be further separated into distinct morphological classes, namely the large, thin, and small basiconic sensilla, respectively (Figure 1A). In addition, each morphological type of sensilla is segmented into sensilla subtypes, which are defined by the invariable combinations of OR genes expressed by the ORNs they house. These sensilla subtypes are morphologically identical, and differ only by the combination of olfactory receptors that are expressed. Stereotypical distribution of sensilla types and subtypes on the olfactory appendages also yields a stereotypical expression pattern of OR genes on distinct zones on the antenna [26]. For instance, in D. melanogaster, Or65a, Or47b, and Or88a ORNs are always found together within the same trichoid sensilla, designated as at4 (antennal trichoid 4), which is typical distributed on the anterolateral region of the antenna.
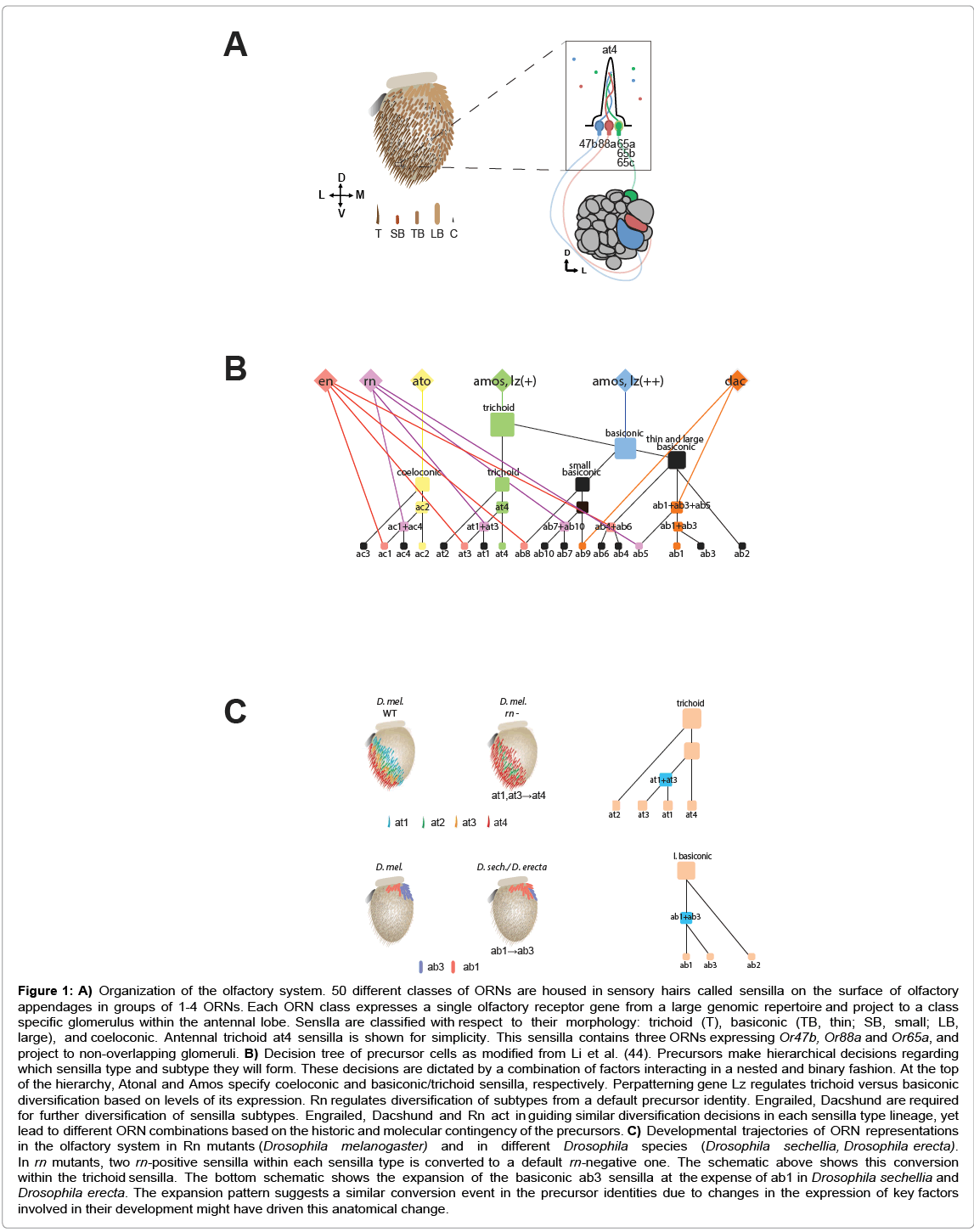
Figure 1: A) Organization of the olfactory system. 50 different classes of ORNs are housed in sensory hairs called sensilla on the surface of olfactory appendages in groups of 1-4 ORNs. Each ORN class expresses a single olfactory receptor gene from a large genomic repertoire and project to a class specific glomerulus within the antennal lobe. Senslla are classified with respect to their morphology: trichoid (T), basiconic (TB, thin; SB, small; LB, large), and coeloconic. Antennal trichoid at4 sensilla is shown for simplicity. This sensilla contains three ORNs expressing Or47b, Or88a and Or65a, and project to non-overlapping glomeruli. B) Decision tree of precursor cells as modified from Li et al. (44). Precursors make hierarchical decisions regarding which sensilla type and subtype they will form. These decisions are dictated by a combination of factors interacting in a nested and binary fashion. At the top of the hierarchy, Atonal and Amos specify coeloconic and basiconic/trichoid sensilla, respectively. Perpatterning gene Lz regulates trichoid versus basiconic diversification based on levels of its expression. Rn regulates diversification of subtypes from a default precursor identity. Engrailed, Dacshund are required for further diversification of sensilla subtypes. Engrailed, Dacshund and Rn act in guiding similar diversification decisions in each sensilla type lineage, yet lead to different ORN combinations based on the historic and molecular contingency of the precursors. C) Developmental trajectories of ORN representations in the olfactory system in Rn mutants (Drosophila melanogaster) and in different Drosophila species (Drosophila sechellia, Drosophila erecta). In rn mutants, two rn-positive sensilla within each sensilla type is converted to a default rn-negative one. The schematic above shows this conversion within the trichoid sensilla. The bottom schematic shows the expansion of the basiconic ab3 sensilla at the expense of ab1 in Drosophila sechellia and Drosophila erecta. The expansion pattern suggests a similar conversion event in the precursor identities due to changes in the expression of key factors involved in their development might have driven this anatomical change.
The axons of each ORN class converge on the antennal lobe in the central brain to form class-specific connections with Projection Neurons (PNs) within bulbous structures called glomeruli. Again, the position of each glomeruli relative to others are conserved from one individual to the next, and the activation of different combinations of these stereotypical glomeruli create an odotopic map, in which each odorant is associated with a specific pattern of activation of the antennal lobe [27-29]. The PNs from these glomeruli then connect to higher processing and decision-making centers in the brain, causing odor-specific shifts in behavior.
The stereotypical, multi-layered structure of the olfactory system allows for many different ways in which evolution can modify or change an insect’s response to an odorant. Increased sensitivity to an odorant, such as by specialists species to their hosts, can potentially be brought about by increasing the binding affinity of OBPs, increasing the number or sensitivity of associated ORs within an OSN, an expansion of a sensilla type or a conversion from one sensilla type to another (both leading to an increased number of ORNs), a conversion from one ORN type to another within a sensilla type, an expansion of glomerular size (i.e. connections with more PNs), circuitry changes in the brain centers, or any combination of the above. A similar variety of different developmental changes can potentially bring about a decrease in sensitivity or a switch in behavioral response to any particular odorant.
One relatively well-studied example of the wide variation these structures can exhibit is the glomerular morphology of the antennal lobe in various insects. The best-studied of these is, unsurprisingly, the model organism Drosophila melanogaster, in which each individual glomerulus has been identified and labeled, and the specific ORN class corresponding to each glomerulus is known [23]. However, in moths and other species in which the males depend heavily on pheromone detection exhibit a dramatic sexually dimorphic expansion of certain glomeruli. Termed as Macroglomerular Complexes (MGC), this phenomena is thought to be involved in signal processing of female sex pheromones. An even more dramatic change in antennal lobe structure can be found in the order Orthoptera (grasshoppers and crickets), in which some species have evolved an antennal lobe consisting of thousands of microglomeruli innervated by highly branched ORNs and PNs, the purpose of which is as yet unknown [30,31].
Besides that, recent research has also introduced other ways in which the olfactory system can be modulated. For example, there is some evidence that G-protein signalling plays an important role by functioning as a presynaptic gain-control mechanism that finetunes olfactory behavior [32]. This GABA (B)-dependent system has been found in fruit flies as well as in male moths [33], suggesting a widespread and evolutionary conserved system for fine-tuning olfactory behavior. Other G-proteins are also known to be important for the normal response of CO2-sensing neurons [34]. As a whole, this suggests the presence of an additional layer of complexity by which olfactory behavioral outputs can be modified.
The complex organization of the olfactory system, along with the many different developmental mechanisms underlying it, provides a fascinating opportunity for us to study how changes in development over evolutionary time can create such a rich diversity of insect ORNs and the neuronal circuits driving odor-guided responses. The large variability in the olfactory sub-genome of sequenced insects further allows an integrated and multi-pronged approach to the speciesspecific diversification of behavioral phenotypes.
The development of the insect olfactory system has received considerable attention in the past several years, particularly by neurobiologists interested in studying the development of neural circuits. The architecture of the olfactory system provided a unique model for neurobiologist to answer questions from topics such as ORN specification, axon targeting, topographic mapping, and neuronal plasticity. However, virtually all of the research in insect olfactory development has been done in the model organism Drosophila melanogaster, and little is known about how developmental processes that set up the olfactory system might differ in other insect species that accounts for the phenotypic variability in the structure and function of olfactory circuits.
In Drosophila, adult ORNs develop from multipotent Sensory Organ Precursors (SOPs) in the larval antennal imaginal disc. Developmental decisions in these cell lineages are determined by the hierarchical interactions of different transcription factors [35-37]. Each SOP has to make decisions regarding which type and subtype of sensilla it will become, which determines the possible combinations of OR gene to be expressed in each sensillum. The decisions regarding sensilla type a given precursor cell will give rise to are determined by proneural basic helix-loop-helix transcription factors, Amos and Atonal [38,39], which are required for the development of basiconic/ trichoid and coeloconic sensilla SOPs, respectively. Sensilla subtype identity decisions of SOPs on the other hand are dictated by the combinations of transcription factors such as Rotund, Engrailed and Daschund and ultimately restrict the particular ORN classes that can be formed from a given precursor cell (Figure 1B) [36,40]. Once the SOPs have restricted their differentiation potentials, cell-cell interactions mediated by Notch-delta signaling during asymmetric SOP divisions are also known to be important in the segregation of cell-fate decisions [41,42] Besides that, there is also good evidence that ORN specification in Drosophila depends on epigenetic mechanisms involving the modification of chromatin states in the developing neuron [42,43], but the relationship between transcription factor expression and these epigenetic mechanisms remains to be elucidated.
Recent research has provided further insight into the transcription factory-mediated precursor cell decisions responsible for the diversity of olfactory neurons [44]. Under this model, sensory precursor cells undergo nested, bifurcating cell fate decisions, with the concurrent use of the same transcription factors in parallel lineages to give rise to the different sets of ORNs that underlie each sensilla subtype. For example, the transcription factor Rotund operates in multiple parallel lineages within each sensilla type to specify a new subset of ORNs that are distinct from a default fate in a binary ON/OFF fashion. When Rotund is lost, ORNs that normally develop from rn-positive SOPs revert to a ‘default’ rn-negative SOP identity. This leads to expansion of “default” rn-negative ORNs within each antennal sensilla type zone at the expense of rn-positive ORNs (Figure 1B and C). The hierarchical and nested structure of the transcription factor interactions driving each of these decisions allows the easy addition of new regulatory nodes at different stages of precursor cell development to economically regulate neuronal diversity in the peripheral olfactory system over an evolutionary time frame. Thus, this system allows for the rapid modification of ORN specification programs in response to changes in the environment and highlights the plasticity of the olfactory system as a whole.
The wiring of each ORN class to a specific glomerulus, suggests that connectivity of each ORN is tightly linked to the developmental programs regulating sensory identity. For example, perturbations in Notch signaling during asymmetric SOP divisions, results in the duplication of both sensory and wiring identities of one ORN class at the expense of its neighbor in the same sensillum [41]. Axonal targeting of the different classes of adult ORNs to their appropriate glomerulus is less well-understood but seems to be a stepwise process starting with the pre-patterning of the antennal lobe by projection neurons [45,46], and subsequently by the correct contact of these PNs to their corresponding ORNs [46,47]. Several different factors including teneurins, acj6 and drifter are known to be involved in PN targeting [48-51], while guidance molecules such as teneurins, Dscam, semaphorins and robo have also been identified as important players in ORN axon guidance process [51-54]. This process is also influenced by the degenerating larval olfactory system [47,55], but the mechanisms responsible for patterning the larval olfactory system remains to be elucidated.
Although these developmental processes have been fairly wellcharacterized in Drosophila melanogaster, almost nothing is known about whether or not any of these processes are conserved in other insect species. Given the large differences in the olfactory sub-genome of even closely-related species, as well as rampant gene duplication and pseudogenization across the board, how these developmental processes change over time to accommodate each species’ unique ORN repertoire is an interesting question and one that we hope to be able to answer in the future. Another potentially interesting area of further inquiry is into the ecological constraints that keep OR gene loci unstable and plastic in number and sequence. Changes in gene expression in response to the changes in the quantity, quality, and context of the olfactory environment can add new regulatory modules to pre-existing lineage-specific combinations to facilitate the coordination of novel ORN fates with the evolution of receptor genes or lead to elimination of specific ORN classes.
One tantalizing example of developmental and molecular mechanisms underlying species-specific differences in neuronal circuit structure is the discovery of microRNA mutant that affects CO2 olfactory circuit structure in D. melanogaster [56]. The CO2 receptors in D. melanogaster and other insects belong instead to a class of receptors known as the Gustatory Receptors (GRs), but similarly to ORs, the CO2 receptors are primarily expressed in the peripheral olfactory organs and converge onto a single glomerulus in the brain. In D. melanogaster CO2 avoidance is mediated by the antennal ORNs that express CO2 receptors and connect to a ventrally-located glomerulus in the antennal lobe. On the other hand, in the mosquito Anopheles gambiae that is highly attracted to CO2, these receptors (that share a common origin with those of D. melanogaster) are instead expressed in the maxillary palps and connect to a medially-located glomerulus in the antennal lobe. These opposing behavioral responses to CO2 must have evolved through multiple steps including elimination of CO2 neurons from one olfactory appendage and their generation in another, as well as changes in connectivity. Interestingly, miR-279 mutants generate ectopic CO2 ORNs in the maxillary palps, and these ORNs exhibit the connectivity of both fly and mosquito CO2 ORNs by projecting to both medial and ventral glomeruli in the antennal lobe (Figure 2). The microRNA miR- 279 is expressed in maxillary palp SOPs undergoing asymmetric cell divisions and here it suppress the formation of CO2 neurons by downregulating certain transcription factors (e.g. nerfin-1).
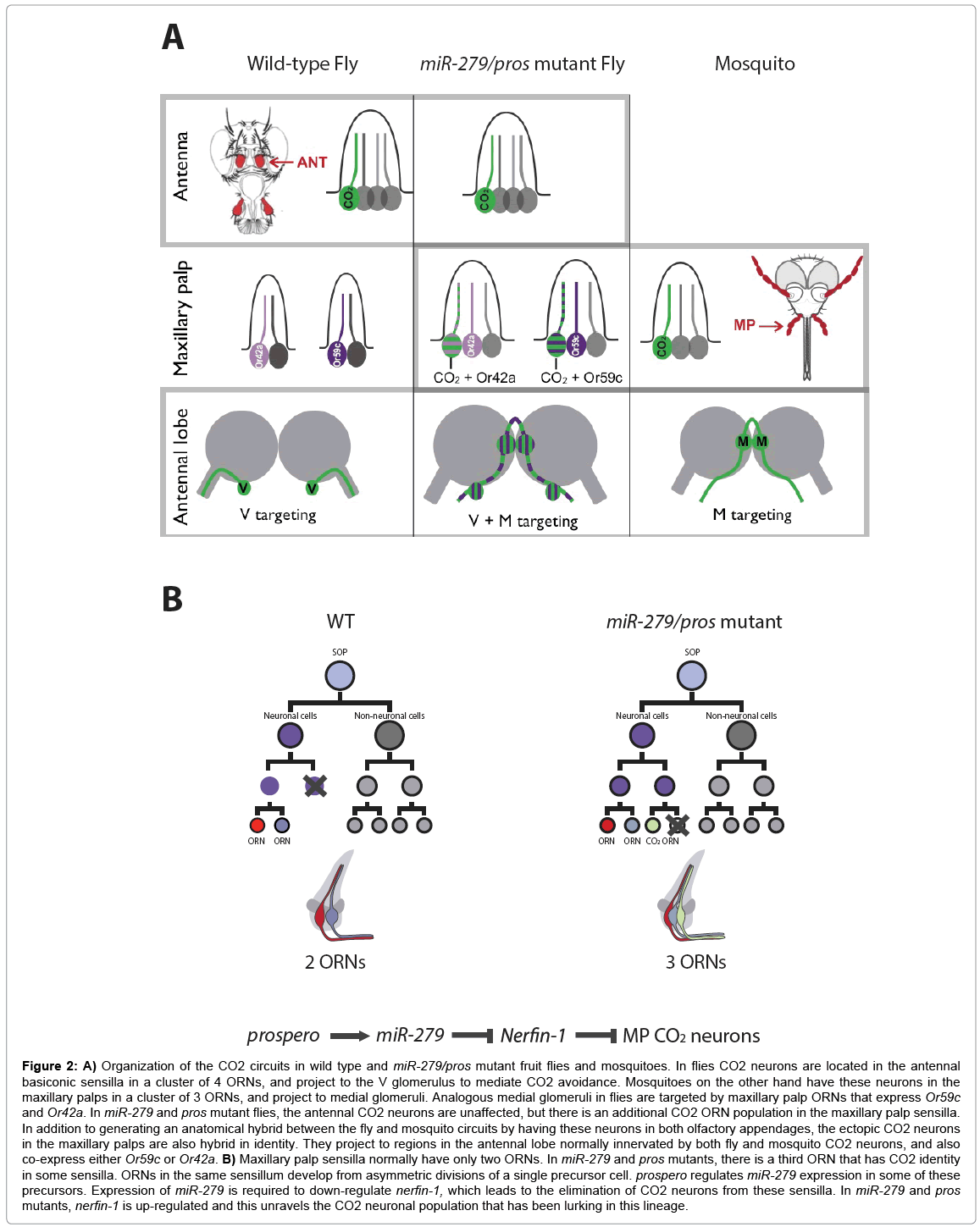
Figure 2: A) Organization of the CO2 circuits in wild type and miR-279/pros mutant fruit flies and mosquitoes. In flies CO2 neurons are located in the antennal basiconic sensilla in a cluster of 4 ORNs, and project to the V glomerulus to mediate CO2 avoidance. Mosquitoes on the other hand have these neurons in the maxillary palps in a cluster of 3 ORNs, and project to medial glomeruli. Analogous medial glomeruli in flies are targeted by maxillary palp ORNs that express Or59c and Or42a. In miR-279 and pros mutant flies, the antennal CO2 neurons are unaffected, but there is an additional CO2 ORN population in the maxillary palp sensilla. In addition to generating an anatomical hybrid between the fly and mosquito circuits by having these neurons in both olfactory appendages, the ectopic CO2 neurons in the maxillary palps are also hybrid in identity. They project to regions in the antennal lobe normally innervated by both fly and mosquito CO2 neurons, and also co-express either Or59c or Or42a. B) Maxillary palp sensilla normally have only two ORNs. In miR-279 and pros mutants, there is a third ORN that has CO2 identity in some sensilla. ORNs in the same sensillum develop from asymmetric divisions of a single precursor cell. prospero regulates miR-279 expression in some of these precursors. Expression of miR-279 is required to down-regulate nerfin-1, which leads to the elimination of CO2 neurons from these sensilla. In miR-279 and pros mutants, nerfin-1 is up-regulated and this unravels the CO2 neuronal population that has been lurking in this lineage.
It is plausible that miR-279 mutants reveal a putative evolutionary intermediate state between Drosophila and mosquito CO2 circuitry. The phenotype can be classified as an anatomical intermediate with the simultaneous presence of CO2 neurons in both olfactory appendages, as well as a cellular intermediate in the form of a hybrid CO2 neuronal population that exhibits connectivity to medial and ventral glomeruli, simultaneously. Natural selection can work on such a hybrid neuron to generate different combinations of odorant receptors, and wiring patterns depending on the selective pressures that the insect is exposed to in its environment. For example, it can match up a CO2 receptor program with a ventral or medial targeting program; ventral targeting may drive attraction as is seen in Drosophila while medial targeting may promote the CO2 attraction characteristic of mosquitoes. These suggest that regulatory mechanisms presumably have evolved, which have allowed such differential coupling of different receptors to different targeting programs that in turn allow for evolutionary flexibility. These mechanisms may aid generation of different olfactory responses to the same odorant, such as attraction versus repulsion, or to different odorants in different species. miR-279 exists in the mosquito genome, and miR-279 binding sites in mosquito nerfin-1 3’UTR are conserved [56], yet it is unclear whether mosquitoes express miR-279 in their maxillary palps. In the Drosophila maxillary palps, miR-279 expression is regulated by the transcription factor prospero [57]. Thus, it is possible that acquisition of Prospero binding sites upstream of Drosophila miR-279, might have contributed to the expression of miR- 279 and elimination of CO2 neurons from this olfactory appendage in Drosophila (Figure 2). Despite the lack of cellular mechanisms associated with this circuit restructuring, these findings provide us with an insight into developmental modifications that can occur in this fastevolving system.
Recent research into host specialization within the Drosophila genus has also highlighted how parallel ecological adaptations can be generated by changes in olfactory development. It has been known for some time now that the specialization of D. sechellia to the Morinda fruit correlates with an expansion of one particular class of sensilla, ab3, which houses the Or22a ORN, in the antennae. Or22a is thought to be the main detector of Morinda volatiles. The expansion of ab3 sensilla corresponds with the reduction of another sensilla class, the ab1 sensilla, and as one would expect D. sechellia also has corresponding increases in the volume of the Or22a glomerulus on the antennal lobe [13,58]. Interestingly enough, a recent study has suggested that the specialization of the closely related Drosophila erecta to its own seasonal host, Pandanus sp., may be driven by largely the same developmental changes [59]. D. erecta has an increased sensitivity towards the characteristic Pandanus volatile 3-methyl-2-butenyl acetate (3M2BA). Surprisingly, this adaptation is again thought to be modulated by the Or22a sensilla, albeit to a different odorant than in D. sechellia. Similarly, it also has an increased number of ab3 sensilla coupled with a reduction of ab1 sensilla, along with an increased Or22a glomerular volume. In light of research into the parallel and nested nature of the regulatory factors that underlie sensilla development, this shared phenotype could be due to the loss or change in expression of a factor that functions similar to that of Rotund to drive specification of the large basiconic sensilla subtypes (Figure 1C). Together, these studies suggest the exciting possibility that the adaptation of an olfactory circuit to a specific host follows a developmentally-specific trajectory, at least in some cases. Whether or not species-specific differences arise largely due to changes in amino-acid sequences for OR genes or transcription factors, changes in cis-regulatory elements, or changes in other RNA regulatory mechanisms remains an open question.
In the past, comparative studies in insect olfactory development have been limited primarily by the difficulties that come with using nonmodel organisms. Even with insect species that are relatively easy to grow and maintain in the lab such as Drosophilid flies, definitive answers to evo-devo questions have been elusive due to difficulties in creating transgenic individuals, limiting the questions that can be answered to mainly those of an anatomical, genomic, or electrophysiological nature. However, due to recent advances in methods for targeted mutagenesis this may no longer be a problem in the near future. Novel genetic engineering methods of particular note include the use of Zinc-Finger Nucleases (ZFNs) [60,61] and Transcription Activator-Like Effector Nucleases (TALENs) [62], in conjunction with methods such as the CRISPR-Cas mechanism [63,64] as cheaper and more efficient ways to create transgenic individuals from non-model species. These methods have already been shown to work in Drosophila [61] and species such as the mosquito Anopheles gambiae [65], and the increased use of such methods in the future on a more diverse range of insect species has very good potential for enabling finer-scaled examinations of how the developmental process differs in these different species.
The insect olfactory system presents a unique opportunity for study in multiple areas. The multilayered complexity of its organization provides a fascinating opportunity for research into interacting and overlapping developmental processes. Furthermore, the modular plasticity of these developmental processes allowed for by the use of parallel, nested regulatory factors lends itself well to a study of how a complex system can be rapidly modified over evolutionary time. Coupled with the rapid evolution of the large olfactory sub-genome and the incredible diversity of the insect olfactory environment, the insect olfactory system allows us to study the intersection between evolution, ecology and development on a much finer scale. As such, continued research into the evolutionary development of the insect olfactory system should prove to be an interesting and fruitful endeavor.