Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2016) Volume 4, Issue 1
The present study focused to elucidate the effect of methyl farnesoate (MF) on ovarian development and maturation in the freshwater giant prawn, Macrobrachium rosenbergii. Control, eyestalk ablated (ESX) and MF injected prawns were maintained during the experimentation and observed significant (p<0.001) increase in the mean ovarian index, oocyte diameter and ovarian vitellogenin levels in both eyestalk ablated and methyl farnesoate administered prawns when compared with controls. All the MF injected animals are in vitellogenic stage III, whereas ESX animals were showed all the reproductive stages at 30th day of experiment. The levels of protein and lipid in the ovary were also measured. The total protein and lipid content levels were also significantly increased in eyestalk ablated (145.07% and 72.05% respectively) and MF injected animals (315.21% and 118.39% respectively) than the controls. The outcome of the present study clearly suggests that MF stimulates the reproduction in female prawns than the conventional ESX method.
<Keywords: Methyl farnesoate, Ovarian maturation, Vitellogenin levels, Total protein, Lipid
Cultivation of crustacean species plays an important role in the world aquaculture industry. In recent past, limited availability of quality seed constitutes the largest cause of destruction of aquaculture industry. In many respects, aquaculture is still in its infancy and it is necessary to improve the aquaculture by producing high quality and quantity seed from commercially important crustaceans. ESX is the technique, used by hatchery industries to induce maturation and hatching, often this technique lead to production of inferior quality seed in crustaceans [1] and shows high mortality in the brood stock due to disruption of the endocrine system, loss of hemolymph and repeated spawning which ultimately decreases the larval quality and viability. Therefore, the conventional ESX method is not in effective usage in hatchery industry. To negate this problem, researchers developed several non-surgical methods including hormonal manipulation and it has been demonstrated that supplementation and administration of hormones, opioids and biogenic amines influence ovarian maturation in edible crustacean such as prawns and crabs [2-4].
The well-known reproductive regulatory hormones in crustaceans are gonad-inhibiting hormone/Vitellogenesis inhibiting hormone (GIH/VIH) synthesized and secreted from the X-organ-sinus gland complex located in eyestalk neural tissue [5,6] gonad-stimulating hormone (GSH) synthesized and released from brain and thoracic ganglion [7-11] and a sesquiterpene hormone methyl farnesoate a secretary product of mandibular organ (MO) [12-15]. In specific MF is a positive regulator for reproduction and is negatively controlled by mandibular organ inhibiting hormone (MOIH) [2,16-19] which suppressed the synthesis and release of MF from MO. An evidence of MF on regulation of reproduction has been reported in the crab Oziothelphusa senex senex[17,19], the crayfish Procambarus clarkii [20], the prawn Machrobrachium malcamsonii [18] and the shrimp Penaeus vannameii [21]. The action of MF on reproduction in crustaceans is not yet established. However reports are suggesting that MF can induce reproduction neither directly by inducing vitellogenin synthesis in the ovary and/or in extra-ovarian sites nor by inducing ecdysteroids the reproduction inducing hormone, synthesis and secretion from Y-organs or by both [17,19]. Since no clear established mechanism of MF in crustacean reproduction, further studies are needed and hence the MF has been selected for the present study.
MF is structurally similar to the insect juvenile hormones (JH) [16] and JH play key role in the regulation of metamorphosis, reproduction and behavior in insects (see reviews [22,23]). Similar to JH, MF also regulates the crustacean reproduction and metamorphosis [24]. Reasonably good literature is available on MF regulated reproduction and metamorphosis in crustaceans [14,18,20,25-28]. Even then findings on the MF induced reproduction in many crustaceans are in its infancy.
Besides fin fish culture, Indian aquaculture industry is focusing on culturing of freshwater giant prawn Macrobrachium rosenbergii and the marine tiger prawn Penaeusmonodan. Due to public demand the major amount of prawn culture is exporting as proteinacious food and selling with a high price. The prawn culture is located throughout the Indian coastal areas. But the costal aquaculture industry is facing many problems in cultivating crustacean species, including limited availability of quality seed.
In this connection, the present study is initiated to induce reproduction in the brood stock of freshwater prawn Macrobrachium rosenbergii by endocrine manipulation with MF.
Intact, pre-vitellogenic stage (identified by observing the colour beneath the carapace) adult freshwater female prawns (M. rosenbergii ) with a body weight 20 ± 2 g were collected from culture ponds in and around Nellore, P.S. Nellore District, Andhra Pradesh, India. They were acclimatized to the laboratory conditions (Temperature 27 ± 1°C; Relative Humidity 75% and a light period 12 h) for 7 days in large glass tanks containing pond water with aerators. Trans, trans-methyl farnesoate (purchased from Echelon Biosciences, USA) was dissolved in 95% ethanol (solvent vehicle).
One hundred prawns were divided into four groups of each [25]. First group served as initial controls and sacrificed on day ‘0’ of experimentation. Second group served as concurrent control injected with crustacean saline [29] of same volume with the experimental animals. Bilateral eyestalk ablated (ESX) prawns were served as third group. Fourth group was injected with an effective dosage (proven in earlier studies) i.e., 10-8 mol of MF/prawn (10 μl) on 1st, 7th, 14th, 21st and 28th day of experimentation. At 30th day, all the second, third and fourth group animals were ice anesthetized, dissected and ovaries were isolated and used for further analysis. Part of the ovaries were cleaned and stored at -20°C for measuring ovarian vitellogenin levels from all the groups.
Measurement of ovarian Index and oocyte diameter
Ovarian tissues were dissected out, blotted with paper towels and weighed wet using electronic balance and the gonad index was determined by using the following formula.
Ovarian index = (Wet weight of the ovary/Wet weight of the prawn) × 100
For measuring oocyte diameter from each freshly isolated ovary, the diameter of 25 oocytes was measured using an ocular micrometer under the compound microscope. The average diameter of 25 oocytes in each ovary was considered as oocyte diameter.
The number of oocytes in each ovary of the experimental animals was also recorded with binocular.
Ovarian histology
Based on morphological observation ovaries of various stages were dissected from the prawns and preserved in formalin and then ovarian sections were made according to the method described by Bancraft and Stevens (1982). Isolated ovaries were gently rinsed in physiological saline solution (0.9% NaCl) to remove adhesive tissues and fixed them in aqueous Bouin’s fluid. After 24 h of fixation the reproductive tissue was dehydrated through a series of alcohol. The tissues were cleaned in xylene and then embedded in paraffin wax (melting point 56.8-58.8°C).
Ovarian sections of 7 μm thick were made and stained with hematoxylin followed by eosin.
Quantification of ovarian vitellogenin levels by ELISA: Vitellogenin (Mr-Vtg) protein was isolated from the ovaries of prawn M. rosenbergii by the method described by Tsukimura et al. [30]. In brief, 10% (W/V) ovarian tissue homogenates were prepared in homogenization buffer (pH 7.8) containing 0.1 M sodium chloride (NaCl), 0.05 M Tris, 1 mM ethylenetriamine tetra acetic acid (ETTA) and 0.1% tween-20 with 10 mg/ml phenylmethylsulfonyl fluoride (PMSF), using an ice cold glass-glass homogenizer. The homogenate was centrifuged at 4000X g for 5 min at 4°C. Resultant supernatant was collected and again centrifuged at 20,000X g for 20 min at 4°C. To the supernatant, ammonium sulfate (SAS) was added to produce a 25% SAS solution. After mixing the contents for 1 h at 4°C, the homogenate was centrifuged at 20,000X g for 10 min at 4°C. The supernatant was collected and SAS was added to produce 40, 50 and 60% SAS solution sequentially. The pellets of 60% SAS solution was suspended in homogenization buffer and dialyzed three times against homogenization buffer at 4°C for 12 h each. The purified ovarian vitellogenin was stored at -20°C until further use.
The quality of isolated ovarian Mr-Vtg was determined using discontinuous Sodium Dodecyl Sulphate-Poly Acrylamide Gel Electrophoresis (SDS-PAGE). The purified Mr-Vtg was stored at -20°C until further use. The ovarian vitellin content in each purified sample was estimated by Enzyme Linked Immuno Sorbent Assay (ELISA). ELISA was performed on certified 96-well microtiter plate. Wells were coated with 20 μl of dialyzed sample diluted in 1:10 ratio with coating buffer (carbonate buffer: 1.59 g sodium carbonate, 2.93 g sodium bicarbonate in 1000 ml distilled water, Diethyl dithio carbonate (DIECA) 56 mg per 25 ml buffer). Blank values were obtained from wells coated with buffer alone. The plate is covered with a lid and placed in a humid chamber at 37°C for 2 h. After discarding the contents, the plate was washed five times with 0.1 M Phosphate Buffer Saline containing 0.05% Tween-20, pH 7.2 (PBST). Then, 200 μl of diluted (1:1000) primary antibody (antibodies raised in rabbits against Mr-Vtg isolated from vitellogenic stage III ovary) in 0.1 M PBST with 2% polyvinyl pyrrolidine, 0.2% ovalbumin (PBST-PO) is added to each well. The plate is covered with a lid and placed in a humid chamber at 37°C for 2 h. The plate was washed 5 times with PBST after discarding contents. After that 200 μL of horseradish peroxidase (HRP) conjugated anti IgG antibody (1:1000 dilution with PBST-PO; purchased from Genei, Bangalore) was added to each plate and kept in dark for 1 h at 37°C. The plate was washed 5 times with PBST and 200 μL Tetra Methyl Benzidene (TMB in 0.015% hydrogen peroxide) was added to each plate. The plate was kept in dark for 1 h at 37°C. The reaction was stopped by adding 50 μL of 1 M phosphoric acid per well. Absorbance of each well was measured with ELISA reader (Bio-Rad Imark, USA) at 450 nm. All standards and samples measurement were performed in duplicate.
Estimation of ovarian total protein and lipid: The total soluble protein was estimated with Bovine serum albumin (BSA) as standard. For this, 25 mg wet tissue of ovary was taken in clean and dry test tubes and added 1 ml of 10% trichloro acetic acid (TCA). The mixture was homogenized and centrifuged at 3000 rpm for 15 min. The protein precipitate was dissolved in 5.0 ml of 1.0 N NaOH and estimated as per the Folin-Ciocalteu method of Lowry et al. [31].
The lipids were quantitatively determined by Sulphophosphovanillin method of Barnes and Blackstock [32]. In brief, 10 mg of ovarian tissue was homogenized in chloroform, methanol mixture (2:1 ratio) and kept overnight in tightly stoppered test tubes at 4°C for complete extraction. The contents were mixed well and centrifuged at 3000 rpm for 15 min. The supernatant containing the lipids was transformed to clean and dry test tubes. From this 0.5 ml was taken in a separate test tube and dried. To the dried samples, 0.5 ml of concentrated sulphuric acid was added and mixed well. The tubes were then plugged with non-absorbent cotton wool, incubated at 100°C in a boiling water bath for 10 min and rapidly cooled to room temperature by placing the tubes under running tap water. To 0.1 ml of this acid digest, 2.5 ml of Phosphovanillin reagent was added and mixed. The pink-red colour was developed and read after 30 min at 520 nm in UVspectrophotometer (Shimadzu, Model No. UV1800, Japan) along with the cholesterol standard against reagent blank (Phosphovanillin reagent was prepared by mixing 800 ml orthophosphoric acid, 200 ml double distilled water and 2 g vanillin).
Statistical analysis: The significance of differences between the means was determined by performing one- way ANOVA with the help of statistical software Graph Pad Prism Version 5.0.3.477. Statistical significance was considered at ‘p<0.05’.
Change in ovarian index, oocyte diameter, ovarian vitellogenin levels, ovarian total lipid and protein were observed not significant in initial control and concurrent control groups (Tables 1 and 2). The ovarian index, oocyte diameter and ovarian vitellogenin was significantly (p<0.001) increased in eyestalk ablated, MF injected animals when compared to initial control and concurrent control animals (Table 1).
| Group | Stage | Ovarian index | Oocyte diameter (µm) | Ovarian Vitellogenin levels |
|---|---|---|---|---|
| (µg/mg tissue) | ||||
| Control | Pre-vitellogenic stage | 0.26 ± 0.01 | 48 ± 8.1 | 0.165 ± 0.061 |
| Concurrent control | Pre-vitellogenic | 0.28 ± 0.04 | 49 ± 8.5 | 0.175 ± 0.052 |
| stage | -7.69 | -2.08 | -6.06 | |
| ESX | Vitellogenic | 0.98 ± 0.20 | 80 ± 10.3 | 0.186 ± 0.103 |
| stage I (N=7) | -276.92 | -66.66 | -12.72 | |
| Vitellogenic | 1.25 ± 0.18 | 264 ± 14 | 0.232 ± 0.120 | |
| stage II (N=7) | -380.76 | -450 | -40.6 | |
| Vitellogenic | 1.31 ± 0.19 | 410 ± 40.2 | 0.253 ± 0.109 | |
| stage III (N=6) | -403.38% | -754.16 | -53.33 | |
| MF injected | Vitellogenic | 1.58 ± 0.03 | 530 ± 36.5 | 0.392 ± 0.112 |
| stage III | -508.84% | -1004.16 | -137.57 | |
| p value | - | <0.001 | <0.001 | <0.0001 |
| F-value | - | 853.14 | 1670.3 | 22.646 |
Table 1: Effect of eyestalk ablation (ESX), injection of methyl farnesoate (MF) on the ovarian index, oocyte diameter and vitellogenin levels in the fresh water prawn, M. rosenbergii. Values are mean ± S.D. of 25 individual prawns. Significance was considered at p<0.05. Values in parenthesis are percent change from controls.
At the 30th day of experimentation vitellogenic stages I-III were observed (Figures 1A-1D) with varying ovarian index, oocyte diameter and ovarian vitellogenin levels in ESX group. The ovarian histology of all four stages of M. rosenbergii ovarian development was observed in the Figures 2A-2D.
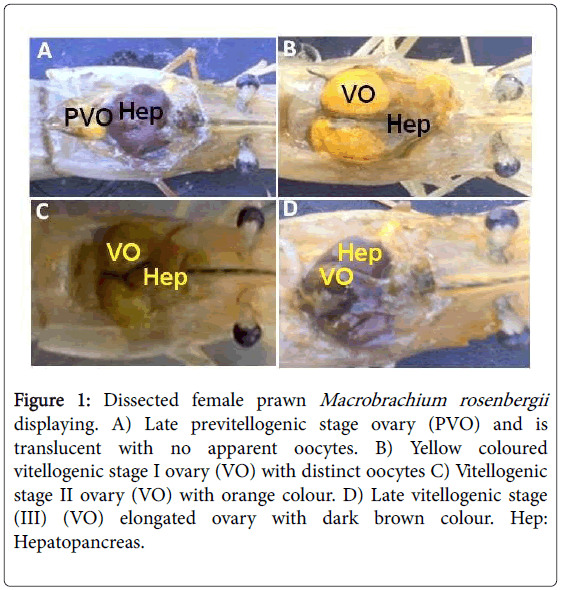
Figure 1: Dissected female prawn Macrobrachium rosenbergiidisplaying. A) Late previtellogenic stage ovary (PVO) and is translucent with no apparent oocytes. B) Yellow coloured vitellogenic stage I ovary (VO) with distinct oocytes C) Vitellogenic stage II ovary (VO) with orange colour. D) Late vitellogenic stage (III) (VO) elongated ovary with dark brown colour. Hep: Hepatopancreas.
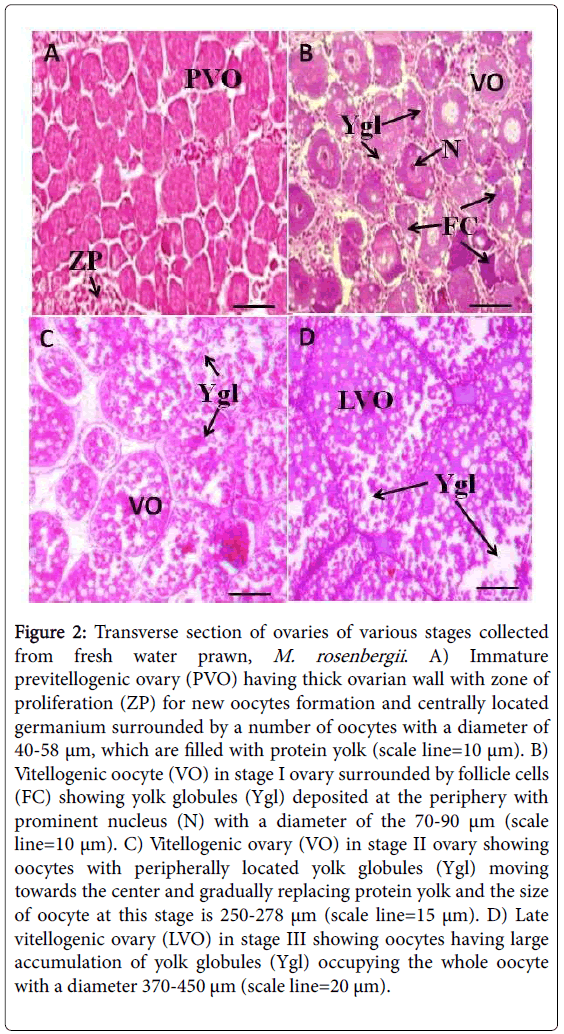
Figure 2: Transverse section of ovaries of various stages collected from fresh water prawn, M. rosenbergii. A) Immature previtellogenic ovary (PVO) having thick ovarian wall with zone of proliferation (ZP) for new oocytes formation and centrally located germanium surrounded by a number of oocytes with a diameter of 40-58 μm, which are filled with protein yolk (scale line=10 μm). B) Vitellogenic oocyte (VO) in stage I ovary surrounded by follicle cells (FC) showing yolk globules (Ygl) deposited at the periphery with prominent nucleus (N) with a diameter of the 70-90 μm (scale line=10 μm). C) Vitellogenic ovary (VO) in stage II ovary showing oocytes with peripherally located yolk globules (Ygl) moving towards the center and gradually replacing protein yolk and the size of oocyte at this stage is 250-278 μm (scale line=15 μm). D) Late vitellogenic ovary (LVO) in stage III showing oocytes having large accumulation of yolk globules (Ygl) occupying the whole oocyte with a diameter 370-450 μm (scale line=20 μm).
Out of 25 from ESX group 5 animals were died during experimentation due to over loss of hemolymph. In ESX group the ovaries of 7 animals were in vitellogenic stage I with an increase of 276.92, 66.66 and 12.72% in ovarian index, oocyte diameter and ovarian vitellogenin levels respectively. The colour of the ovary was pale yellow (vitellogenic stage II) in 7 ESX animals with an increase of 380.76, 450 and 40.60% in ovarian index, oocyte diameter and ovarian vitellogenin levels respectively. A total of 6 animals from ESX group were in vitellogenic stage III with an increase of 403.38, 754.16 and 53.33% in ovarian index, oocyte diameter and ovarian vitellogenin levels respectively. Besides this, also recorded the number of oocytes in both the experimental groups and found highest number in MF injected group (36%) than ESX animals.
No mortality was observed in MF injected group and observed all of them are in late vitellogenic stage (III) with significant increase (p<0.0001) in ovarian index (508.84%), oocyte diameter (1004.16%) and ovarian vitellogenin levels (137.57%) (Table 1). MF is known to be involved in the regulation of reproduction in various crustaceans [18,25,28,33-37]. Similar to ovarian index, oocyte diameter and ovarian vitellogenin levels, observed significant increase (p<0.0001) in the ovarian total lipid and protein in both ESX and MF injected prawns (Table 2).
| Group | Stage | Total lipids | Total Proteins |
|---|---|---|---|
| (mg/g tissue) | (mg/g tissue) | ||
| Control | Pre-vitellogenic | 10.25 ± 2.05 | 25.12 ± 3.12 |
| stage | |||
| Concurrent control | Pre-vitellogenic | 10.89 ± 2.35 | 25.89 ± 3.52 |
| stage | -6.24 | -3.06 | |
| ESX | Vitellogenic stage | 14.54±1.45 | 30.44 ± 3.10 |
| I (N=7) | -41.85 | -21.17 | |
| Vitellogenic stage | 15.20 ± 3.44 | 30.99±2.78 | |
| II (N=7) | -48.29 | -23.36 | |
| Vitellogenic stage | 16.02 ± 2.66 | 32.22±4.62 | |
| III (N=6) | -56.29 | -28.26 | |
| MF injected | Vitellogenic stage | 17.56 ± 3.52 | 3.86±5.86 |
| III | -71.31 | -34.79 | |
| p value | - | <0.0001 | <0.0001 |
| F-value | - | 24.961 | 14.915 |
Table 2: Ovarian total protein and lipid in eyestalk ablated (ESX) and methyl farnesoate (MF) injected fresh water prawn M. rosenbergii. Values are mean ± S.D. of 25 individual prawns. Significance was considered at p<0.05. Values in parenthesis are percent change from controls.
In MF injected animals, the lipid and protein levels were significantly increased when compared with control and eyestalk ablated animals. In eyestalk ablated animals, the highest ovarian lipid and protein level was 56.59 and 28.26% respectively. Whereas MF injected animals showed increase in ovarian lipid and protein levels (71.31 and 34.79% respectively) from that of initial controls.
The role of ESX in the regulation of ovarian maturation has been documented in many crustaceans. Bilateral extirpation of eyestalk results in removal of X-organ sinus gland complex, which eliminates an array of hormones, including GIH/VIH regulates maturation negatively [28,38,39] and leads to precocious ovarian maturation [40,41]. Induced ovarian development and increased ovarian weight along with hemolymph vitellogenin levels and ovarian vitellogenin mRNA levels in the kuruma prawn Marsupenaeus japonicas by ESX was reported [42]. It is also reported that bilateral ESX increases the size of MO in Libinia emarginata [43] and Palaemonetes varians [44] and increased circulatory MF levels which ultimately precocious ovarian maturation. Similar to the ESX group of present study, several authors reported the increase of protein content and lipid level in the ovary during maturation [45-47].
In the present study the role of MF in the induction of ovarian maturation in prawn M. rosenbergii was elucidated. In support of this, accelerated ovarian maturation by the administration of MF in Procambarus clarkii was reported by Laufer et al. [20]. Similarly by the injection of MF ovarion maturation was initiated in crab O. senex senex [17,19,28]. In vitro stimulation of ovarian growth by MF has also been reported for both penaeid shrimp Penaeus vannamei [21] and crayfish Procambarus clarkii [26]. Incubated hepatopancreas explants of crab Charybdis feriatus with farnesoic acid, MF and JH III and observed up regulation of vitellogenin gene expression in hepatopancreas of farnesoic acid incubated explants [48]. Whereas in crab O. senex senex observed vitellogenin expression in hepatopancreas incubated with MF in vitro [19].
Significant increase (p<0.001) in ovarian index, oocyte diameter, ovarian vitellogenin levels, ovarian total protein and total lipid is observed in prawns injected with MF than control groups in this study. MF induced maturation in prawn M. rosenbergii is much higher than the conventional eyestalk ablation method. At the end of experiment, found only vitellogenic stage III with more number of oocytes in MF injected animals, whereas in ESX group it is vitellogenic stage I-III. The results of the present study are supported by many studies. Nagaraju et al. [49], reported fourfold increase in ovarian index and oocyte diameter in Penaeus indicus by the administration of MF. In the same study they also observed that all the MF injected animals were in late vitellogenic stage (III). Increased ovarian index, oocyte diameter and ovarian vitellogenin levels in MF injected crab Oziothelphusa Senex senex was reported [17,19,25,]. From the present study results, it is clearly indicating that MF induces ovarian maturation better than eyestalk ablation technique in freshwater prawn M. rosenbergii along with increased number of oocytes.
Furthermore it is predicted that MF may increases the deposition of high amounts of vitellogenin, total protein and total lipid in ovary thereby increases in size of the egg. Due to its activity, MF may facilitate the progress of conversion of pre-vitellogenic oocytes into vitellogenic oocytes thereby increase in number. Being a natural hormone of crustaceans, MF induces reproduction without modifying the genetic pool. The present study is providing a way to focus on the usage of MF to produce high quality, more susceptible and viable seed which is badly required for withstand of World aquaculture industry thereby produced quality protein to meet the protein scarcity demand of ever growing human population. However, further metabolic and molecular level studies have to be established on MF induced reproduction in crustaceans. So that the usage of MF directly by the hatchery industry or by the aqua formers be established in expectations which ultimately produces quality crustacean protein with cost effective.
The authors thanks the University Grants Commission (UGC), New Delhi for financial assistance in the form of research grants to Dr. B. Kishori, Assistant Professor, Department of Biotechnology, Sri Padmavati Mahila Viswavidyalayam, Tirupati [F.No. 39-273/2010(SR)] and partly from Dr. P. Ramachandra Reddy, Assistant Professor, Department of Biochemistry, Yogi Vemana University, Kadapa [F.No. 41-582/2012(SR)]. The authors are highly thankful to Prof. P. Sreenivasula Reddy, Department of Zoology, Sri Venkateswara University for valuable guidance in executing this research work.