Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2016) Volume 5, Issue 3
Studies on the Hypothalamo-Neurohypophysial System (HNS) have played a pivotal role in establishing the fundamental principles of neuroscience. Examples include the identification of excitation-secretion coupling, the coexistence of peptides-containing large dense core vesicles with classical neurotransmitters, neuronal-glial interactions, functions associated morphological plasticity, and domain-specific regulation of the secretory activity, etc. Moreover, mechanisms underlying the secretion by the HNS and functions of neuropeptides oxytocin and vasopressin remain the central topic of the neuroendocrine society, novel findings of which keep substantiating the model role of this system. Nevertheless, challenges are still severe in this study field, such as extrahypothalamic connections of the HNS, scaffolding and guiding roles of glial fibrillary acidic protein in astrocyte plasticity, cellular and molecular processes determining different firing patterns, the position of the HNS in neuroendocrine-immune network, and the nose-hypothalamic route that serves as a key mediator of drug effects on brain activity. Nowadays, translational studies of the HNS also become the prime time of researchers, such as vasopressin involvement in brain injury in ischemic stroke, and therapeutic potentials of oxytocin in lactation failure and its associated health issues. In this paper, we review these challenges in modern neuroscience studies on the HNS.
<Keywords: Neuropeptides; Neuroscience; Oxytocin; Supraoptic nucleus; Vasopressin
GFAP: Glial Fibrillary Acidic Protein; HNS: Hypothalamo-Neurohypophysial System; HPA Axis: Hypothalamic- Pituitary-Adrenal Axis; MNCs: Magnocellular Neurosecretory Cells; OXT: Oxytocin; OXTR: OXT Receptor; PVN: Paraventricular Nucleus; SON: Supraoptic Nucleus; VP: Vasopressin
In the hypothalamus, there are two types of peptidergic magnocellular neurosecretory cells (MNCs): oxytocin (OXT)- producing neurons and vasopressin (VP)-producing neurons. OXT and VP (also called antidiuretic hormone, ADH) are the first group of neuropeptide hormones being sequenced and synthesized biochemically by Du Vigneaud, the Nobel prize laureate in 1955. This achievement largely determines the chemical nature of the mysterious pituitrin, pitressin and pitocin factors of the posterior pituitary that had been questioned since the identification of the vasopressor, uterus contraction, milk-letdown and antidiuretic effects of pituitary extracts. In studying on the neuroendocrine cells, particularly the MNCs of the Hypothalamo-Neurohypophysial System (HNS) that was initially established by Bargmann and Scharrer, mechanisms underlying the secretion and functions of VP and OXT have been the central topic of neuroendocrine society. From functional studies of circulating nonapeptides that extensively modulate the activities of almost all organ systems, to their brain actions that are represented by the autocrine regulation and prosocial effects, and then to the hidden secretion of peripherally produced OXT and VP that subtly adjust the activities of individual cells, studies on the HNS have been playing a pivotal role in establishing the fundamental principles of the neuroscience and the working models of neuropeptides. These achievements have been excellently reviewed and will not be further presented here [1,2].
Among several hypothalamic nuclei containing the MNCs, the supraoptic nucleus (SON) has many features that are suitable for characterizing neuron activities and their underlying mechanisms. For example, these MNCs have relatively large cell bodies for their identification and electrophysiological manipulation, large amounts of secretory products for biochemical quantification, and relatively homogeneous cell populations that are easy to separate from other brain regions [3]. Thus, studies on the SON have greatly contributed to the concepts of the excitation-secretion coupling, neuronal-glial interactions, functions-associated morphological plasticity, and domain-specific regulation of the secretory activity, etc. In addition, the SON also has the following important features: 1) A clear separation of the somatodendritic zone from the axon terminals, which allows separate studies on the release profile of neuropeptides in the brain and in the circulation; 2) A distinguished spatial orientation of astrocyte processes, which is suitable for observing glial-neuronal interactions at microdomain and molecular levels; 3) Coexpression of classical neurotransmitters with neuropeptides, which provides a model to observe the interactions between different types of neurotransmitters in the autoregulation of MNC activity, and 4) Dependence of release patterns of the blood neuropeptides on their intranuclear release and on the activity of decomposing enzyme. After the initial findings, these features are also identified in many other types of neurons, particularly neuroendocrine cells [4,5]. Therefore, the SON remains a "model system" in our pursuing neuroscience research. In this review, we outline the challenges that we face in current studies on the HNS and our perspectives of future studies.
Histological features of the HNS
In the MNCs, OXT with its carrier protein neurophysin I and VP with neurophysin II are packaged in large, membrane-bound dense-core vesicles that are transported down the long axons to the nerve endings in the posterior pituitary [6]. In response to changes in extracellular environment, the MNCs are excited to generate action potentials that propagate down the axons and elicit exocytosis of some of these vesicles through excitation-secretion coupling. Released intos extracellular space, these neuropeptides diffuse into the bloodstream through fenestrated capillaries and act on corresponding receptors including V1a, V1b and V2 subtypes of VP receptors and OXT receptor (OXTR) [7,8]. Moreover, the firing activity and release profiles of the MNCs are modulated by synaptic innervations, astrocytic-neuronal interactions, blood-borne factors and auto-regulation of MNC activities [9-12]. Broadly speaking, MNC-associated presynaptic terminals and astrocytes are also the integrative parts of the HNS that is mainly composed of the MNCs and their terminals.
The MNCs in the SON have 1-3 large dendrites that mainly project ventrally to receive most of the presynaptic terminals from afferent neurons and participate in information processing. Moreover, the dendrites and axonal collaterals of supraoptic neurons can release OXT and VP to exert neuromodulatory effects. The axonal efferent from the SON can project into various extrahypothalamic regions including the olfactory bulb (OB), the cortex, the amygdala, and the mammillary body complex in addition to a densely labeled pathway running from the SON to the posterior pituitary through the internal median eminence [13]. Although some cells around the SON could absorb radio-labeled leucine in the aforementioned study and partially account for the neural projection from the SON to these extrahypothalamic regions, it is clear that MNCs from the SON can modulate both hypothalamic and extrahypothalamic neuron activity. This proposal is further supported by the following evidence. A small proportion of the MNCs in the SON can send axon collaterals to the suprachiasmatic nucleus, a brain center regulating circadian rhythm, the antero-ventral third ventricular region near the organum vasculosum of the lamina terminalis where osmosensory neurons are identified [14], and to the medial preoptic area where gonadotropin-releasing hormone neurons are present [15]. Furthermore, dendrites of the MNCs can reach the ventricular system and thus exert neurohormonal effects on extensive CNS areas through the CSF [16,17]. In addition, there are close interactions between the SON and the paraventricular nucleus (PVN), which could be mediated by both neural innervations and paracrine modulations [18-20]. These connections form a structural basis of the simultaneous activation of a large pool of HNS neurons in the bilateral hypothalamus, typically observed in OXT neurons during suckling [21]. Importantly, interactions of some brain functions with OXT neurons in the SON are different from those in the PVN, such as that the activity of the gonadotropin-releasing hormone neurons in the medial preoptic area has close interactions with OXT neurons in the SON but not in the PVN [15]. These facts indicate that the neuromodulatory effects of MNCs in the SON on brain activity and behaviors are largely underestimated over the years, largely because parvocellular division of the PVN has been considering as the major source of brain OXT and VP after its early findings [22]. Moreover, the SON likely exerts different effects from the PVN does on some brain functions, such as the regulation of estrous cycle [23] and of olfaction [24]. Nevertheless, further studies are required to depict the exact trajectories of these neural projections from the MNCs. Figure 1 is a schematic illustration of the histology of the SON and the targets of its neural projections.
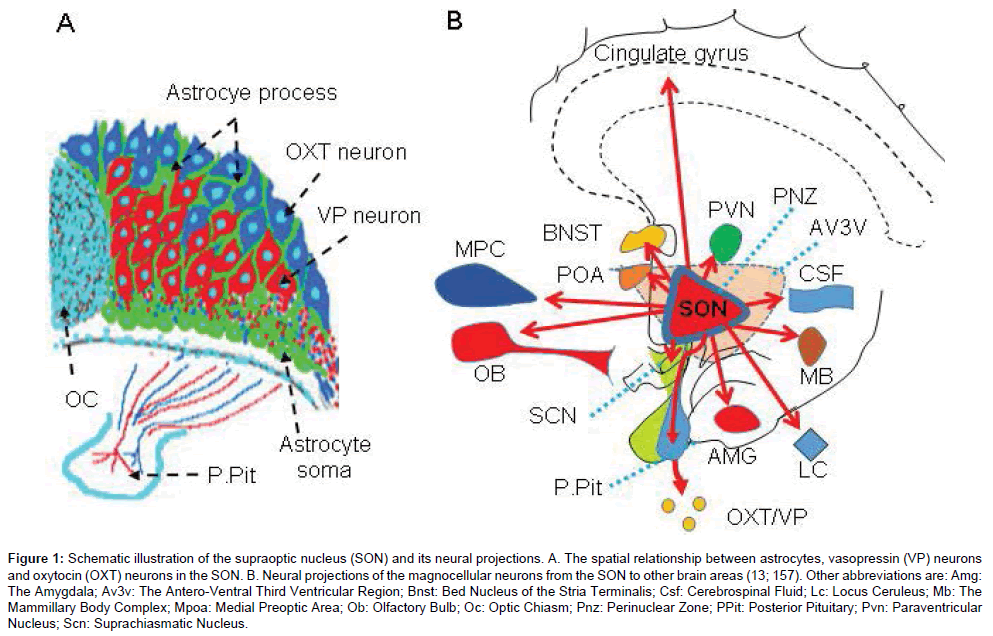
Figure 1: Schematic illustration of the supraoptic nucleus (SON) and its neural projections. A. The spatial relationship between astrocytes, vasopressin (VP) neurons and oxytocin (OXT) neurons in the SON. B. Neural projections of the magnocellular neurosecretory cells (MNCs) from the SON to other brain areas (13; 19). Other abbreviations are: Amg: The Amygdala; Av3v: The Antero-Ventral Third Ventricular Region; BNST: Bed Nucleus of the Stria Terminalis; CSF: Cerebrospinal Fluid; LC: Locus Ceruleus; MB: The Mammillary Body Complex; MPC: medial prefrontal cortex; OB: Olfactory Bulb; OC: Optic Chiasm; PNZ: Perinuclear Zone; POA: Preoptic Area; PPit: Posterior Pituitary; PVN: Paraventricular Nucleus; SCN: Suprachiasmatic Nucleus.
Morphological and functional plasticity
An important feature of the HNS is its morphological plasticity. As revealed by electron microscopic observations in rats, dehydration increases the size of the MNCs and significantly reduces glial coverage of these cells in the SON [25]; lactation increases the size of OXT neurons, the numbers of shared synapses, and the apposition between adjacent OXT neurons [26]. This plastic change was originally thought to be specific to OXT neurons [27] that are mainly located in the dorsal part of the SON [28]. However, a more detailed morphologic analysis revealed that VP-ergic dendrites become more extensive in the SON, which is likely to fill in the open space left by the reduction of dendritic branches and total dendritic length of OXT neurons during lactation [29]. This morphological plasticity can partially explain the synchronized activation of OXT neurons during suckling as well as the changes in the excitability of MNCs in response to osmotic challenges that have been previously reviewed [10,30] and thus will not be further discussed here.
It is worth noting that the chronic morphological plasticity can be compensated by some other mechanisms since disabling this plasticity in the SON did not significantly influence OXT neuronal activity as shown in studying the burst discharges of OXT neurons during suckling stimulation [31]. In contrast, acutely occurred morphological plasticity in the SON is likely irreplaceable for the milk-ejection reflex and for the osmotic reaction and thus constitutes the structural basis of acute functional plasticity of the HNS [32-34].
Acute morphological plasticity of the HNS under different functional states is closely related to acute astrocytic plasticity [10] that is largely dependent upon plastic changes in GFAP molecules [35]. In the studies on supraoptic astrocytic morphological plasticity in rat models of lactation and dehydration [36-38], GFAP plasticity has been used as an indicator of gliosis or the retraction of astrocyte processes. The GFAP plasticity is closely related to astrocyte functions, such as buffering changes in extracellular K+ and glutamate levels [39], regulating extracellular volume transduction [40], adjusting physical barriers between adjacent neurons and many others [33,34,39].
The contribution of GFAP to astrocytic plasticity has been well shown in the SON during suckling [37] and in response to hypoosmotic challenges [41,42]. However, the scaffolding and guiding roles of GFAP in acute astrocytic plasticity are not well known. For example, in glutamate metabolism, GFAP has molecular associations with glutamine synthetase and serine racemase and thus, could relocate these molecules during its polymerization and depolymerization [37], thereby modulating glutamate-glutamine cycle in a microdomainspecific manner as previously discussed [32,43]. Similarly, GFAP has also molecular association with vesicular GABA transporters and thus can modulate GABA metabolism and actions [41]. These evidences suggest that GFAP can be a scaffold to provide a platform for the interactions between different molecules while serving as a transporter to guide their spatial localization of functions.
The contribution of the scaffolding and guiding roles of GFAP to astrocytic plasticity is further supported by the following studies. Aquaporin 4 (AQP4), another binding partner of GFAP, shows changes in its expression levels as the retraction or expansion of astrocyte processes around OXT neurons at different stages of suckling stimulation [37]. The membrane installation of AQP4 following the occurrence of the milk-ejection burst could allow more water to get into the cells and facilitate the expansion of astrocyte processes. This could be accompanied by the activation of Na+, K+, 2 Cl- cotransporter-1 [44] and AQP4-coupled inwardly rectifying K+ channel 4.1 [39], as a result of burst-evoked increase in extracellular K+ levels [45,46]. The activation of these ion transporters decreases extracellular K+ levels and could also lead to the post-burst inhibition [47] in OXT neurons. Conversely, the reduction of AQP4 during glial retraction exerts an opposite effect: reducing absorption of extracellular K+ and glutamate, leading to gradual increases in the inter-burst firing rate in OXT neurons during suckling [47]. Since GFAP plasticity occurs extensively in a variety of physiological and pathological conditions, such as the site-dependent GFAP plasticity in ischemic stroke [39,48], further exploration of the scaffolding and guiding roles of GFAP in astrocytic plasticity will profoundly impact our understandings of the glialneuronal interactions and in turn manifest new targets for treatment of astrocyte-associated diseases. Figure 2 is a diagrammatic drawing of the scaffolding and guiding roles of GFAP in acute astrocytic plasticity and their influences on neuronal activity.
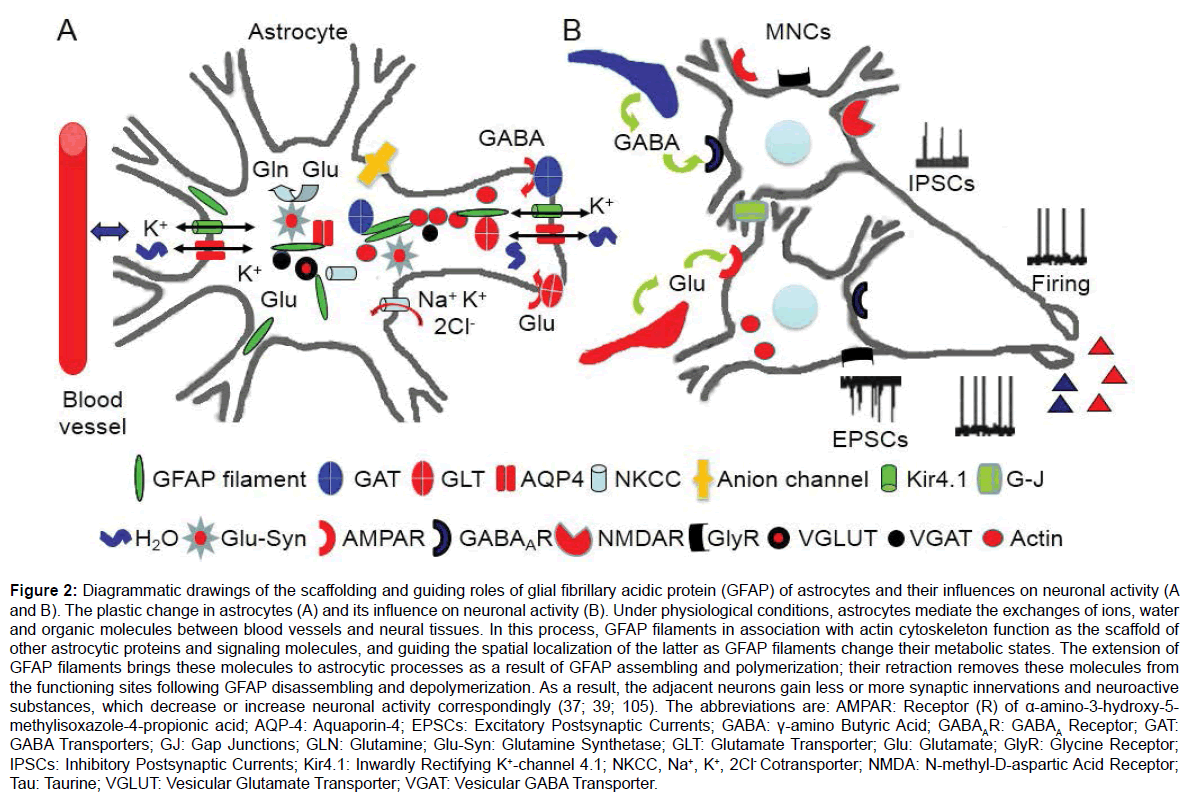
Figure 2: Diagrammatic drawings of the scaffolding and guiding roles of glial fibrillary acidic protein (GFAP) of astrocytes and their influences on neuronal activity. The plastic change in astrocytes (A) and its influence on neuronal activity (B). Under physiological conditions, astrocytes mediate the exchanges of ions, water and organic molecules between blood vessels and neural tissues. In this process, GFAP filaments in association with actin cytoskeleton function as the scaffold of other astrocytic proteins and signaling molecules, and guide the spatial localization of the latter as GFAP filaments change their metabolic states. The extension of GFAP filaments brings these molecules to astrocytic processes as a result of GFAP assembling and polymerization; their retraction removes these molecules from the functioning sites following GFAP disassembling and depolymerization. As a result, the adjacent neurons gain less or more synaptic innervations and neuroactive substances, which decrease or increase neuronal activity correspondingly (37; 39; 105). The abbreviations are: AMPAR: Receptor (R) of α-amino-3-hydroxy-5-methylisoxazole-4- propionic acid; AQP-4: Aquaporin-4; EPSCs: Excitatory Postsynaptic Currents; GABA: γ-Amino Butyric Acid; GABAAR: GABAA Receptor; GAT: GABA Transporters; GJ: Gap Junctions; Gln: Glutamine; Glu-Syn: Glutamine Synthetase; GLT: Glutamate Transporter; Glu: Glutamate; GlyR: Glycine Receptor; IPSCs: Inhibitory Postsynaptic Currents; Kir4.1: Inwardly Rectifying K+-channel 4.1; NKCC, Na+, K+, 2Cl- Cotransporter; NMDAR: N-methyl-D-aspartic Acid Receptor; Tau: Taurine; VGLUT: Vesicular Glutamate Transporter; VGAT: Vesicular GABA Transporter.
Cellular and molecular processes leading to different firing patterns
In response to different physiological demands, VP neurons and OXT neurons exhibit different firing patterns and secretory profiles of the neuropeptides. For example, hyperosmotic and hypovolemic stimuli can increase the release of VP that in turn causes water reabsorption in the kidney and vasoconstriction to reduce blood osmolality and elevate blood pressure. In this process, the firing rate of VP neurons increases in a phasic pattern that occurs periodically based on a plateau potential [49]. Hyperosmotic stimulation also increases the firing activity of OXT neurons and OXT release that appear in a tonic pattern but not in phasic or burst pattern [50]. During lactation [51] and parturition [52], the firing activity of OXT neurons appears in an intermittently bursting pattern on the background of a continuous or irregular firing activity. This milk-ejection burst is highly synchronized among OXT neurons in the SON and the PVN [21,53] and is followed by a bolus release of OXT [54]. In general, the phasic and burst firing patterns are more efficient for hormone release than the tonic firing pattern does with the same number of spikes [49]. Compared to the phasic firing activity of VP neurons, the burst firing in OXT neurons is much shorter and more intense [55]. This high frequency burst discharges of OXT neurons can account for the bolus release of OXT during suckling stimulation while the randomly occurred phasic firing in VP neurons can explain the intensively increased VP release in response to hyperosmotic challenges. This association between firing patterns and neurosecretion of the MNCs provides a suitable model for studying the mechanisms underlying pattern generators of neuronal activities and their association with different functions.
In investigating mechanisms underlying the differences in firing patterns between OXT and VP neurons, many questions have been explored, such as the afferent pathways transmitting different stimuli and the chemical nature of synaptic connections [56], electrophysiological features of VP and OXT neurons in vivo [51] and in vitro [57], genes associated with OXT- or VP-neurons [58], signaling molecules [59] and ion channels [57,60,61] that trigger special firing patterns. Among them, burst firing activity of OXT neurons specifically depends on suckling stimulation [62], intranuclear release of OXT [63], and a neuromodulation of the “burst synchronization center” in the mammillary body complex [64]. Relative to the regulation of OXT neuronal activity, the regulation of VP neuronal activity is “non-specific.” The type of stimulation and extracellular factors relating to the phasic firing are relatively loose although the activity of VP neurons is closely regulated by the anteroventral region of the third ventricle [65]. At the cellular level, OXT but not VP neurons possess a rebound depolarization that underlies a brief burst of action potentials while exhibiting a broader spike width and a larger Ca2+-dependent afterhyperpolarization [66]. However, the identified in vivo electrophysiological features largely reflect the extracellular environment. For example, hyperosmotic stimulation triggers phasic firing while OXT is the permissive factor of burst discharge, which does not show the link between the signaling features of these MNCs and firing patterns. In vitro studies on chemically-identified MNCs can touch the cellular and molecular processes accurately; however, previously identified electrophysiological differences between the two types of MNCs remain controversial. That is, in previous study, the presence of a sustained outward rectifying potassium current and/or an inward rectifying hyperpolarization-activated current are presumably present in OXT neurons but absent in VP neurons [67]. However, the phenotypes classified according to the electrophysiological protocol in brain slices do not match their molecular counterparts because VP and intermediate neurons also exhibit both outward and inward rectifying currents [68]. As a whole, differences in the cellular and molecular processes leading to the different firing patterns are largely unclear.
What we know better are the association between phasic burst in VP neurons and a series of cellular and molecular processes as excellently reviewed [57,69]. In contrast, only a few of publications using in vitro approaches adopted strict criteria that reflect the features of burst firing of OXT neurons. By simulating the neurochemical environment existing during lactation in the SON, several burst models were created in the last two decades. One was produced in a low calcium medium in brain slices from lactating rats [55] or male rats [70] by bath-application of phenylephrine, the α1 adrenoceptor agonist. Another one was established in brain slices from lactating rats [71] by extracellularly applying OXT, the permissive factor of burst occurrence in vivo [63,72]. In the third one, a spontaneous burst of OXT neurons was identified in organotypic cultures of the hypothalamic explants from neonate rats [73]. By using these models, the differences in the firing patterns between the two types of MNCs are outlined. That is, bursts in OXT neurons are shorter in duration, higher in peak firing rates, wider in the half width of action potentials and not superimposed on a plateau potential. Moreover, burst occurrence depends on the activation of OXTRs and glutamatergic synaptic transmission [73] but not on GABAergic synaptic transmission in brain slices [71]. Interestingly, OXT suppresses tonic glutamatergic and GABAergic synaptic transmission [11] but evokes short clustered glutamate and GABA release that is highly correlated with the burst features in OXT neurons [71]. One contradictory phenomenon is the lack of reports of burst evoked directly by exogenous glutamate so far while exogenous GABA in the SON could evoke burst discharge in lactating rats [74]. Thus, it is likely that the burst occurrence in OXT neurons needs an excitatory environment, based on which the mobilization of OXTRspecific signaling cascade can trigger the burst discharges as reviewed previously [75]. In addition, clustered GABAergic inputs on OXT neurons are also likely involved in the burst generation [76], which is assumed to trigger a hyperpolarization-evoked rebound depolarization [66]. However, causal relationship of these clustered EPSCs and IPSCs with the burst onset remains to be elucidated.
At molecular levels, relative to the classical α-subunit signaling in OXT actions, Gαq/11-associated Gβγ subunit signaling plays a dominant role in the burst firing of OXT neurons at both pre- and postsynaptic sites [71]. In the downstream of the Gβγ subunits, phosphorylated extracellular signal-regulated protein kinase 1/2 and actin reorganization at the membrane subcortical areas [77] are essential to the burst generation. However, the dependence of phosphorylated extracellular signal-regulated protein kinase 1/2 [78] and actin reorganization [79] are also essential for the phasic firing in VP neurons. The common involvement of these signaling molecules in different types of the MNCs does not mean that the signaling processes are the same for both the phasic and burst firing patterns. It is possible that the two types of MNCs express these molecules in different spatial and temporal orders in response to their adequate stimuli. That is, the actin cytoskeletal reorganization occurs before the burst in OXT neurons following a cascade of cytosolic signaling molecules during suckling [75] while osmotic challenges in VP neurons elicit actin reorganization following a series of membrane events and triggering the phasic firing [79]. Another line of evidence explaining the differences in the firing patterns is from the studying on cell-specific expressions of calcium binding proteins. Immunohistological studies revealed that OXT neurons have more calbindin-D28k and calretinin than VP neurons [80], which allows Ca2+ oscillation in OXT neurons to be buffered quicker than that in VP neurons. This feature determines that OXT neurons discharge action potentials in either continuous or burst pattern while VP neurons adopt the phasic firing pattern [81]. Nevertheless, it remains to clarify how these different signaling processes are linked to changes in the extracellular milieu and ion channel activity that underlies the different firing patterns.
As more ion channels and their functions are identified in the MNCs, such as ATP-activated transient receptor potential vanilloid channels [82], the approaches of their contributions to the burst firing are also questioned. Further study should clarify if the burst onset is associated with a group of ion channels or a single channel, based on appropriate burst-firing models. Overall, burst model-based experiments are required to identify the expression, distribution, and functions of individual channels as well as their associations with burst-evoking signaling molecules, which should fully consider factors at levels of system, local neural circuit, MNCs, glial-neuronal interaction, receptors and intracellular signaling as well as ion channels of an organism (Figure 3).
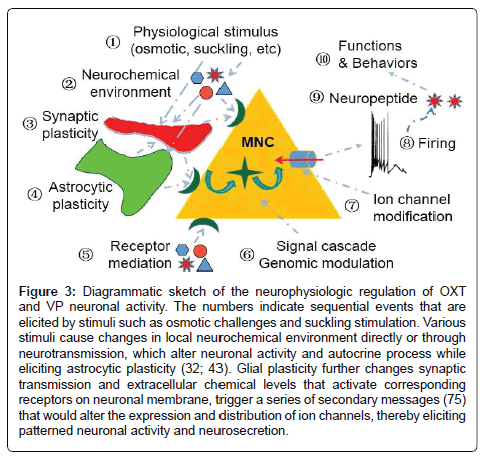
Figure 3: Diagrammatic sketch of the neurophysiologic regulation of OXT and VP neuronal activity. The numbers indicate sequential events that are elicited by stimuli such as osmotic challenges and suckling stimulation. Various stimuli cause changes in local neurochemical environment directly or through neurotransmission, which alter neuronal activity and autocrine process while eliciting astrocytic plasticity (32; 43). Glial plasticity further changes synaptic transmission and extracellular chemical levels that activate corresponding receptors on neuronal membrane, trigger a series of secondary messages (75) that would alter the expression and distribution of ion channels, thereby eliciting patterned neuronal activity and neurosecretion.
The HNS-immune network
The first and a major hypothalamic peptidergic system involving in neuroimmunologic regulation is considered to be the hypothalamic pituitary- adrenal (HPA) axis [83]. The HPA axis refers to a complex set of direct influences and feedback interactions between the hypothalamus, the pituitary gland, and the adrenal glands. Under the regulation and coordination of hypothalamic corticotrophin-releasing hormone, the HPA axis can prime the immune system and potentiate acute defensive responses while suppressing immunologic activities by antagonizing adrenergic pro-inflammatory actions [84].
In parallel with the role of the HPA axis in immune regulation, the HNS is also implicated in neuroendocrine regulation of immunologic activities. On the one hand, the HNS is the target of immunologic activities. For instance, thymectomy causes general atrophy of the SON, PVN and the median eminence [85]; immune cytokines [86] and pathogens [87] can change the activity of neuroendocrine neurons and neuropeptide secretion. On the other hand, the HNS can profoundly modulate the development, differentiation and functions of the immune system. For example, removal of the intermediate and posterior lobes of the pituitary reduces hormone- and cell-mediated immune responses to pathological challenges in rats as recently reviewed [88]. In rat tissue cultures, VP inhibits while OXT promotes the growth of thymic glands [89]. OXT reduces stress-induced psychological illness by strengthening social bonding while VP and corticotrophin-releasing hormone enhance autonomic reactivity and increase susceptibility to adverse mental and physical impacts through its facilitating arousal [90]. In addition, OXT can extensively modulate the activity of immunologic system, such as increasing the production of hematopoietic progenitor cells, reducing inflammatory cytokines, inhibiting the invasion of neutrophils [88]. Consistently, blocking OXTRs inhibits T-cell differentiation in the thymus [91] and activates the excretion of the inflammatory cytokines, interleukin-6 and Chemokine (C-C motif) ligand 5 [92,93] while neutralization of VP specifically and reversibly blocks interferon-γ production in mouse splenic lymphocytes [94]. Moreover, intracerebroventricular administration of VP suppresses the proliferative response of splenic T-cells and natural killer cell cytotoxicity. Consistent with the actions of these two neuropeptides, OXTRs are found in monocytes and macrophages [95], thymic T-cells [96], and mesenchymal stromal cells of adult bone marrow [97] while VP receptors are found on splenic membrane, splenic lymphocytes [96] and CD4+ and CD8+ thymic cells [98]. Clearly, there are bidirectional communications between the HNS and the immune system, in which the HNS could function as a higher neuroendocrine regulatory center of the immune system and modulate immunologic responses differentially and coordinately through OXT and VP, respectively. This proposal expands the view that the OXT-secreting system is a major part of the neuroendocrine center regulating immunologic activity [88].
To claim that the HNS is a higher neuroendocrine regulatory center of the immune system, it is essential to clarify the relationship between the HNS and the HPA axis. On the one hand, glucocorticoids secreted in response to stress activation of the HPA axis can exert rapid negative feedback influences onto the HNS and thus, reduce the secretion of OXT and VP [99,100]. On the other hand, the activity of HPA axis is heavily modulated by VP and OXT. For example, VP can promote the secretion of adrenocorticotropin hormone in the pituitary, particularly in males [101] while OXT inhibits the activity of the HPA axis [102]. Thus, the HPA axis and the HNS form a complex feedback loop while functioning collaboratively in responses to immunologic challenges. Consistently, the central inhibition of VP on the proliferative response of splenic T-cells could be due to VP activation of HPA axis in the brain that directly inhibits splenic responses to pathologic challenges [103]. Nevertheless, it remains to explore the subtle interactions between these immune-associated neuropeptide systems.
Together with the proposal by Pittman [104], we propose that in the neuroendocrine-immune network, the HNS can integrate immunologic, neural signal, and hormonal messages and regulate immunologic activities directly as well as indirectly through the HPA axis (Figure 4). By this way, the HNS can tailor the immunologic responses to meet body’s defense requirement accurately. Further study should differentiate the contributions of the VP-secreting system from that of the OXT-secreting system to the hypothalamus-immune network.
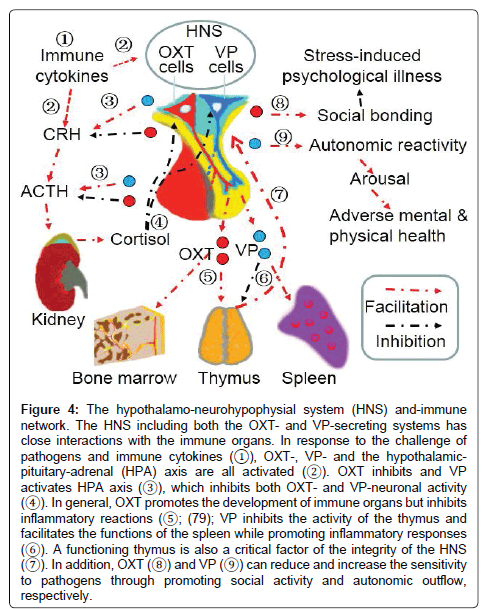
Figure 4: The hypothalamo-neurohypophysial system (HNS) and-immune network. The HNS including both the OXT- and VP-secreting systems has close interactions with the immune organs. In response to the challenge of pathogens and immune cytokines (?), OXT-, VP- and the hypothalamicpituitary- adrenal (HPA) axis are all activated (?). OXT inhibits and VP activates HPA axis (?), which inhibits both OXT- and VP-neuronal activity (?). In general, OXT promotes the development of immune organs but inhibits inflammatory reactions (?; (79); VP inhibits the activity of the thymus and facilitates the functions of the spleen while promoting inflammatory responses (?). A functioning thymus is also a critical factor of the integrity of the HNS (?). In addition, OXT (?) and VP (?) can reduce and increase the sensitivity to pathogens through promoting social activity and autonomic outflow, respectively.
Nose-HNS route mediating nasal drug effects on brain activities
As the accumulation of knowledge of the NHS, particularly their involvement in diseases such as acute hyponatremia [43] and ischemic stroke [105], exploring non-invasive approaches modulating HNS activity has become a strong challenge to neuroscience researchers. Recently, an OB-HNS route has emerged as a potential approach mediating intranasal drug effects on brain activities. In general, intranasally applied drugs can get into the brain bypassing the BBB, through diffusion via the olfactory cells and neural sheathes, through circulation or by activating trigeminal neurons [106]. Nevertheless, drugs diffusing from the olfactory epithelium into the brain get into and accumulate in the OB first [107]. Moreover, OB cells also express OXTRs [108] and VP receptors [109] while circulating neuropeptides are less likely acting on the HNS [110]. Thus, the OB could be the first brain target of nasally-applied neuropeptides and can be the only direct target if these neuropeptide doses are lower enough [111-112]. Relative to the extensively identified brain effects of nasally-applied OXT, the effects of nasal VP are relatively weak and limited [113] and thus, exploring OXT-associated OB-SON pathway likely provides a useful model to study the mechanism underlying the actions of intranasal neuropeptides.
In fact, an OB-SON neural circuit has been well established based on close interactions of OBs with the SON. Stimulation of the OB increases activity of supraoptic neurons [114,115] via the lateral olfactory tract [116]. Conversely, increased OXT neuronal activity causes OXT release in the OB via neural terminals from the SON or its associated interneurons [13]. By activation of OXTRs in the OB, OXT can change the electrical activity of mitral cells [24], the major output neurons in the OB. Thus, there are bidirectional interactions between the OB and the SON, which allows nasal OXT to modulate the activity of OB neurons and subsequently change the activity of MNCs in the SON [117]. Since OXTR has very high sensitivity to low levels of OXT [118], if nasally-applied OXT is controlled at appropriate levels, it is possible to elicit a “specific” effect on SON neuronal activity. This is particularly meaningful to avoid unwanted effects when OXTRs are known to express in extensive brain areas and peripheral tissues [8].
Nasal therapy is a simple yet effective method for medical modulation of brain activity. For instance, OXT administration can greatly attenuate the cortisol response to laboratory tasks that strongly activate the HPA-axis, which is more robust among clinical populations concomitant with social difficulties [119], thereby exerting an anxiolytic effect. These actions could be mediated by the SON. As stated above, axonal collaterals and dendrites of the SON neurons can innervate extensive brain areas directly or through its perinuclear zone neurons or the cerebroventricular system [16,120]. Thus, brain effects of intranasal neuropeptides could be mediated by a nose-OB-SON-brain approach. To achieve a specific effect, future studies are required to identify the distribution of different neuropeptide receptors, the unique or major pathway from the OB to a brain target area, and the relative power of the main OB versus vomeronasal organ [121]. Figure 5 shows a putative nose-brain approach mediated by the OB-HNS route.
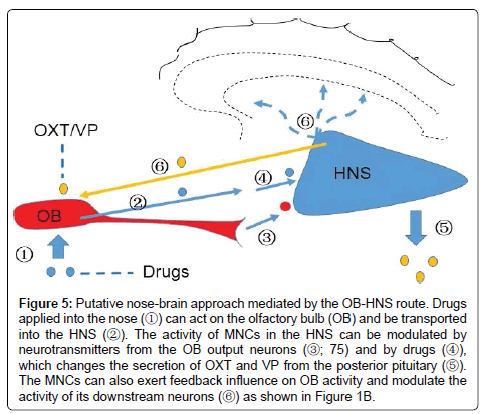
Figure 5: Putative nose-brain approach mediated by the OB-HNS route. Drugs applied into the nose (?) can act on the olfactory bulb (OB) and be transported into the HNS (?). The activity of MNCs in the HNS can be modulated by neurotransmitters from the OB output neurons (?; 75) and by drugs (?), which changes the secretion of OXT and VP from the posterior pituitary (?). The MNCs can also exert feedback influence on OB activity and modulate the activity of its downstream neurons (?) as shown in Figure 1B.
The HNS is extensively involved in a variety of physiological functions while its pathological implications are also explored [122,123]. For example, VP is involved in stress, the hypophysial-adrenocortical axis, neuroimmune responses, renal function and corticotroph pituitary tumors [124]. Similarly, OXT is extensively involved in many body functions such as sexual activity, penile erection, ejaculation, pregnancy, uterus contraction, milk ejection, maternal behavior, osteoporosis, diabetes, cancer, social bonding, and stress among many others [125,126]. Thus, OXT and its receptor have been considered as potential targets for drug therapy of diseases in the brain and peripheral organs [127]. Here, we discuss the clinical relevance of OXT and VP in association with their classical functions.
Abnormality in the VP-secreting system can result in multiple disorders in body functions, typically seen in diabetes insipidus (DI) and the syndrome of inappropriate antidiuretic hormone secretion (SIADH) [128]. DI is characterized by hypotonic polyuria greater than 3 liters/24 hours in adults and persisting even during water deprivation due to a defect in VP synthesis (central DI), VP resistance (nephrogenic DI), or early decomposition of VP by placental enzymes (gestational DI). For the central DI, desmopressin, a synthetic analog of vasopressin, is the treatment option [129]. Conversely, overly produced VP can cause SIADH. SIADH is a condition that the body releases excessive VP from the posterior pituitary or another source, which induces too much water to remain inside the body and results in hypervolemia along with dilutional hyponatremia. In SIADH, physiological mechanisms regulating VP secretion are disrupted, either because of the activation of neuroendocrine tumors or non-osmotic regulatory processes, and thus, the synthesis and release of VP are not inhibited by a reduction in plasma osmolality [43]. Thus, tolvaptan, a VP antagonist can be used along with fluid restriction and diuretics to resolve the fluid retention in the SIADH [130].
Recently, the involvement of VP hypersecretion in ischemic stroke has also been highlighted. In ischemic stroke, brain edema formation is one of the most important pathologic processes [131], in which VP exerts a pivotal role [132,133]. VP release and expression of its receptors in the brain increase significantly following ischemia, trauma or subarachnoid hemorrhage in patients and in animal models of these diseases [134]. VP can increase ion transport across the cell membrane and the blood-brain barrier [135] while increasing AQP4 expression in astrocytes and thus promoting cytotoxic edema [136]. In parallel with the direct effect on hydromineral balance, excessive VP also causes brain arteriole constriction and worsens tissue hypoxia [137], thereby worsening brain edema through oxygen and glucose deprivation. These and other associated factors [138-140] can lead to disruption of the blood-brain barrier and increase hydromineral transfer into the brain, leading to further increases in intracranial pressure and neuron damages. By increasing water retention, cytotoxic edema, disruption of the blood-brain barrier and worsening ischemic injury, VP hypersecretion becomes a major cause of brain injury and disability following stroke. However, mechanisms underlying VP hypersecretion remain unclear [105] although the SON and PVN are known to be involved in the infarct zone in rats [141]. Clarification the mechanisms underlying VP hypersecretion during stroke and suppression of its hypersecretion or the effect of VP receptor signaling processes remain a serious challenge in stroke treatment.
In parallel with the progress in studying the pathological roles of VP, OXT-associated diseases are also explored. Parturition and lactation are the two classical functions of OXT. OXT knockout does not influence parturition; however lactation or breastfeeding is clearly disrupted as shown in animal experiments [142]. Thus, roles of OXT in lactation failure and its associated diseases are discussed here. Breastfeeding has many beneficial effects on the health of mothers and the babies. However, insufficient breastfeeding and lactation failure are common socioeconomic and biomedical issues among the mothers. Insufficient breastfeeding is associated with the high incidence of postpartum depression, premenopausal breast cancer, diabetes and obesity in the mothers and autism, sudden death, and deficiency in maternal behaviors in their offspring [143]. Thus, clarification of the mechanisms underlying lactation failure and exploration of its potential therapeutic approaches are important for the health of both the mothers and the babies.
In general, cesarean section, maternal illness during pregnancy, and preterm birth are considered as the major factors that affect early breastfeeding status [144]. These factors can disrupt breastfeeding through the following approaches. 1) Pain and stress. Factors leading lactation failure are commonly associated with pain and stress, physically or mentally. For example, cesarean section increases the incidence of postnatal depression, psychological birth trauma, birth related post-traumatic stress disorder and postoperative pain [145], which can cause breastfeeding failure through inhibition of OXT neuronal activity by the activated HPA axis. Consistently, maternal separation causes postpartum depression, leading to maternal loss of interests toward her offspring and lactation failure [146]. 2) Immaturity of the OXT-secreting system for lactation. Around the term, the SON experiences a significant morphological and functional remodeling [25,26] that facilitates milk-ejection reflex [147]. However, maternal separation days after the parturition can reverse the facilitatory morphologic changes in the hypothalamus [148] and results in reduction of the secretion of prolactin [149] and OXT [150] along with increased corticosteroid levels [151]. Thus, mothers with preterm babies or separated from the baby could delay the proceeding of this maturation, particularly in the mother with preterm baby. 3) Failure in maintenance of the functional state of the structural basis for the milk-ejection reflex. Suckling stimulates the release of OXT and milk removal [152] while increasing the secretion of prolactin as well [153]. At cellular and molecular levels, without sufficient suckling, OXT synthesis in OXT neurons decreases and OXTRs uncouple with their downstream signals as observed in a maternal separation model of lactating rats [146]. Failure of suckling also causes accumulation of milk in breast ductal system that increases intramammary pressure, blocks milk secretion, elicits oxidative stress and results in the involution of breast tissues [148]. Thus, lacking suckling stimulation not only induces the involution of the OXT-secreting system and the mammary glands, but also disrupts the structural basis for the milk-ejection reflex.
Importantly, breastfeeding failure seeds the risk of breast cancer. A cohort study of parous premenopausal women indicated that breastfeeding is inversely associated with the incidence of breast cancer among women with a family history of breast cancer [154]. However, the biological basis of this anticancer effect of breastfeeding has not gained support of experimental evidence in spite of the presence of many hypotheses. Recently, an animal experiment using phosphorylated extracellular signal-regulated protein kinase 1/2 and cyclooxygenase 2 as indicators of hydrogen peroxide-evoked proliferative activity revealed that lactation failure resulting from lack of pulsatile OXT release during lactation is causally related to the precancerous lesions of the mammary glands [155]. This finding, although requiring observation of long-term effects on the incidence of mammary gland tumorigenesis, suggests that intermittently pulsatile action of OXT during lactation underlies the anti-pre-menopausal breast cancer effects of breastfeeding.
Studies on the MNCs have greatly enriched our knowledge of the peptidergic neurons in modern neuroscience. These include not only the progress in the studies on the classical roles of the HNS, such as neuronal-glial interactions and function-associated morphological plasticity, but also exploration of the scaffolding and guiding roles of glial fibrillary acidic protein in astrocytic plasticity, cellular and molecular mechanisms underlying different firing patterns, the pivotal position of the HNS-immune network in neuroendocrine regulation of immune activity, and the OB-HNS route that serves as a key mediator of nasal drug effects on brain activity. The studies also highlight the translational potentials of modulating HNS activity, such as suppression of VP signaling process to control the pathogenesis of ischemic stroke and utilizing the pulsatile feature of OXT actions in prevention of premenopausal breast cancer. Nevertheless, a number of major questions remain to be addressed, which include but not limited to: 1) The details of neural connections between the HNS and other brain structures and between different nuclei in the HNS; 2) The compensatory mechanisms to reserve the “lost” functions in gene deficient animals, particularly in OXT and OXTR gene knockout mice [156]; and 3) Exploration of the optimal approaches and therapies to treat the HNS-associated diseases. To achieve these goals, it is necessary to apply integrative research approach in the studies on the HNS to build an intact picture from individual molecular events to whole body’s functions/behaviors. It is also important to present “more comparable” data by adopting uniformed and optimized experimental conditions in individual laboratories. By clarifying these questions and resolving emerging challenges, we can better understand the functions of the HNS and their translational potentials, and in turn provide important insights into common disease states.
We thank Dr. Stephani C. Wang for critical reading. The project is sponsored by the National Natural Science Foundation of China (grant No. 31471113, YFW), and the higher education talents funds of Heilongjiang province (grant No. 002000154). All authors declare no conflict of interest.