Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2013) Volume 0, Issue 0
Focus on developing more effective therapeutic interventions to assuage the symptoms of mental health disorders has been a significant driving force in our search to understand the human brain. Current treatments for mental health disorders rely on modulating neurotransmitter systems such as norepinephrine, serotonin, dopamine, and GABA to achieve therapeutic relief of symptoms. While there are many medications that individually target these specific systems, treatment often leads to unwanted side effects or incomplete relief. To counter this lack of treatment efficacy, we must investigate other avenues of achieving the same or better outcomes to potentially reach those patients resistant to common treatments. One of these systems is the Endogenous Cannabinoid System (ECS), which is composed of the metabotropic CB1 and CB2 receptors. These G protein coupled receptors are responsible for mediating the effects of acute cannabis ingestion as well as modulating a number of core functions including emotion, memory, and reward. Due to the ubiquitous nature of the expression of ECS proteins, there is broad overlap between the brain regions that show high levels of receptor expression and those that are involved in mental health disorders. Consequently, CB receptors could be used as a route for modulating the levels of other modulators, such as dopamine, that are known to play crucial roles in the progression of a number of mental health disorders. Here we will review the endogenous cannabinoid system and its natural ligands, along with how this system could potentially be involved in affective disorders, anxiety disorders, and schizophrenia.
<Increasing prevalence of mental health disorders over the course of the last three decades has triggered a dramatic peak in research interest concerning all aspects of these disorders throughout the United States and other westernized countries [1]. While this work has brought about a high level of scientific awareness regarding mental health, it has been strikingly difficult to translate this knowledge into effective therapies. This problem of treatment efficacy is, at least in part, related to the highly stigmatized nature of our population’s perceptions concerning mental health disorders [1]. While there have been significant advances in the field of mental health research, less than 50 percent of those suffering from depression actually receive adequate treatment [1-3]. Many overlapping factors produce this lack of treatment efficacy, including social constructs and physiological side effects of the pharmacological tools used in the course of treatment [1-5]. Consequently, the search to find new molecules to alleviate the symptoms of mental health disorders while reducing the side effect profile of the drug(s) used is of the utmost importance. Here we will review recent evidence that points toward new treatment possibilities that rely on modulation of the endogenous cannabinoid system (ECS) to exert their therapeutic effect.
Cannabis sativa is the most commonly used illicit drug worldwide [6]. With a deep history of cannabis use among humans dating back millennia, it is only within the last century that possession of any form of the cannabis plant has become illegal [7]. The primary psychotropic compound found within cannabis is delta9-terahydrocannabinol as shown in figure 1 (Δ9 -THC). This molecule produces euphoric sensations typically cited as part of the negative consequences of cannabis ingestion [6,8]. While this one substance is responsible for the illegality of cannabis, there are hundreds of related, yet diverse compounds, known as phytocannabinoids, found within the resin of Cannabis sativa that have unknown mechanisms of action [9-11]. These various cannabinoid molecules must be individually studied for potential therapeutic effects that could be produced without the psychotropic side effects of THC. Likely, there are molecules that will prove useful and may have been known in the past, but were lost to us due to recent stigmatization of the plant [7]. Over the past decade, there has been a sizable movement to once again take advantage of the neuromodulatory effects produced by the constituent components of Cannabis sativa. This resurgence has resulted in the emergence of a new population of individuals that ingest cannabis regularly to treat a variety of medical conditions [12-14]. Without firm scientific evidence for or against the efficacy of many of these uses, these individuals are not obtaining the same level of care patients undergoing conventional therapy would obtain. To counter this disparity in the delivery of efficacious treatment, evidence regarding the medical properties of cannabis must be collected to adequately treat this new population of patients. Additionally, this plant exists as a potent source of compounds that could act on targets in the central nervous system (CNS) [8-10]. These compounds need to be isolated and fully characterized as some could provide new modulatory mechanisms that may prove to be more efficacious than current treatments for mental health disorders. Furthermore, characterization of endogenous cannabinoids and cannabinoid receptors in regards to normal physiological function and their possible roles in neuropathophysiology may provide new insight into ECS modulation-based therapies.
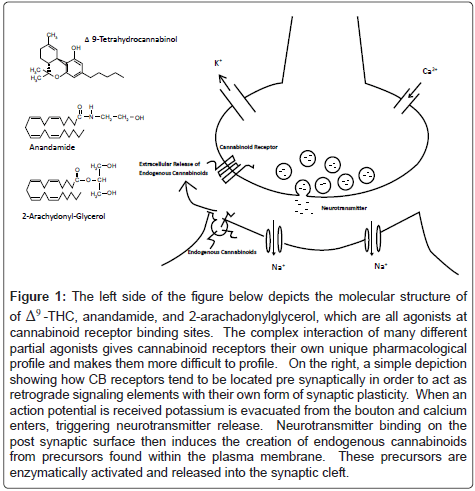
Figure 1: The left side of the figure below depicts the molecular structure of of Δ9 -THC, anandamide, and 2-arachadonylglycerol, which are all agonists at cannabinoid receptor binding sites. The complex interaction of many different partial agonists gives cannabinoid receptors their own unique pharmacological profile and makes them more difficult to profile. On the right, a simple depiction showing how CB receptors tend to be located pre synaptically in order to act as retrograde signaling elements with their own form of synaptic plasticity. When an action potential is received potassium is evacuated from the bouton and calcium enters, triggering neurotransmitter release. Neurotransmitter binding on the post synaptic surface then induces the creation of endogenous cannabinoids from precursors found within the plasma membrane. These precursors are enzymatically activated and released into the synaptic cleft.
We will begin our discussion of the involvement of the ECS in mediating mental health disorders with evidence from human and animal studies of cannabinoid (CB) receptor psychopharmacology. To facilitate understanding of this system, a short overview of ECS anatomy and physiology has been presented [15]. Each subsequent section describes a major mental health disorder including affective disorders, psychotic disorders, and anxiety disorders along with a discussion of how each disorder could have ECS involvement. These will be reviewed briefly with specific attention paid to the neural physiology underlying each of these disorders. Using this perspective of dysfunctional neural physiology, we will explore how our current knowledge of the endogenous cannabinoid system can be applied to achieve therapeutic outcomes in patients suffering from mental health disorders.
The symptoms of mental health disorders are manifestations of underlying deleterious changes in the functionality of molecular pathways mediating cellular communication [1]. When aberrant patterns of activity are present in the brain, it can lead to alterations in the behavioral profile that characterizes each of us as individuals. A variety of neurotransmitter receptor proteins are expressed in neural tissue, forming functionally nested systems. These systems interact to regulate cellular communication with acetylcholine, glutamate, and gamma amino butyric acid (GABA) representing some of the most well characterized signaling systems [15-18]. Cannabinoid receptors are another subclass of this protein signaling system that is defined by these proteins’ ability to bind molecules derived from the Cannabis sativa plant, as well as other synthetic cannabinoid molecules [15]. Unfortunately, research into the endogenous cannabinoid system has yet to be translated into widely accepted and marketable therapies. This is contrary to other receptor systems that have received large amounts of attention from pharmaceutical manufacturers, which is most likely due to the severe federal ban on herbal sources of ligands. However, with the advent of synthetic cannabinoid molecules, significant strides have been made in understanding the anatomy and physiology of this receptor class. Much of this work has revealed the cannabinoid system is similar to other neural systems we have studied and is widely expressed, thus will provide a new way of modulating neural networks expressing pathological features [15-21].
The endogenous cannabinoid system is currently composed of cannabinoid receptor 1 (CB1) and cannabinoid receptor (CB2) with a possible third member, the orphan receptor GPR55 [15,18,22]. These CB receptors are widely distributed throughout mammalian tissue and early immunohistochemistry studies have revealed CB1 receptors are the most abundant receptors in neural tissue, with evidence of neural CB2 only emerging late this past decade [15,23]. Previously, CB2 had been considered the peripheral CB receptor since it is expressed at high concentrations within the spleen and is found on B cells, NK cells, and lymphocytes [23]. Neuroanatomical localization of the CB1 receptor has shown abundant protein expression in the neo-cortex, hippocampus, amygdala, striatum, and midbrain nuclei [15-20,24]. The CB1 receptor has been found primarily on presynaptic axonal terminals with some expression observed on post synaptic cell bodies [15]. All of the CB receptor proteins are members of the super family of seven transmembrane loop G-Protein Coupled Receptors (GPCR’s) bound to an inhibitory internal enzymatic complex (Gi/o) that is released upon receptor activation [15,25]. Activation of CB1 initiates a signaling cascade inhibiting adenylyl cyclase and presynaptic N p/q type calcium channels, in addition to activating MAP kinase proteins and presynaptic inwardly rectifying potassium channels [15,25,26]. This signaling cascade results in cannabinoid mediated retrograde synaptic plasticity. This occurs through synthesis of cannabinoids in the post synaptic density and is dependent upon the arrival of presynaptic signals. These cannabinoids, produced after signal arrival, diffuse away from the synapse and act on the terminal itself, ultimately influencing its long term vesicular output [15,25]. The effects of CB2 activation are very similar to that of CB1, but have not been entirely functionally characterized within brain tissue [23,27,28]. CB receptors have also been identified on astrocytes, with the demonstration that these receptors are functionally relevant on both neurons and glia [27]. This evidence regarding CB mediated neuron to glia communication speaks further to their relevance as a robust signaling system within the CNS.
There are several endogenous molecules that serve as ligands at cannabinoid receptor binding sites, all of which possess a molecular motif making them lipid soluble. The solubility of these compounds allows them to serve as long-term messengers that can perpetuate retrograde signals, which persist in the membrane far after the passage of an action potential. This longevity is part of the therapeutic potential of these compounds, as their effects tend to have significant duration due to the lipophilic structure of these ligands [15,18] (Table 1). This elongation of action creates a broader window of therapeutic action and reduces the chance of lethal overdose. Anandamide (AEA) is the most well documented endogenous cannabinoid and is locally synthesized from ehthanolamine and arachidonic acid precursors in an on-demand fashion based on recent levels of cellular activity [8,15]. This molecule and its relative compounds are produced through hydrolysis of N-arachidonyl phosphatidyl ethanolamine (NAPE) by phospholipase D. Synthesis is based on intracellular levels of calcium and cyclic AMP (cAMP). While basal anandamide concentrations are kept fairly low, the endocannabinoid 2-arachidonyl glycerol (2-AG) is found in much higher concentrations. The primary pathway for synthesis of 2-AG employs the hydrolysis of phosphatidylinositol by phospholipase C and diacylglycerol lipase (DAG). Binding of these natural ligands is differential, with anandamide serving as a low efficacy agonist at both receptors whereas 2-AG has high levels of efficacy at both receptor sites. Along with their unique synthesis pathways, anandamide and 2-AG have separate degradation processes with anandamide being broken down by Fatty Acid Amine Hydrolases (FAAH) and 2-AG by Monoacylglycerol Lipase (MAG) [8,5,18,29-32].
| CB1 | CB2 | |
|---|---|---|
| Agonist | CP 55,940, Arachydonyl- Glycerol (2-AG), | JWH-015, JWH-051, JWH-133, CP 55,940 |
| Partial Agonist | Anandamide (AEA), Δ9-Tetrahydrocannabinol (THC, Dronabinol), WIN 55,212-2, | WIN 55,212-2, Anandamide (AEA),Δ9-Tetrahydrocannabinol, Arachydonyl- Glycerol (2-AG) |
| Antagonist/Inverse Agonist | SR 141617A, LY 320315, AM 251 | SR144528, AM630 |
Table 1: This table shows the relationship between a variety of cannabinoid compounds and their selectivity for either CB1 or CB2. Most agonists have very non selective action and most knowledge regarding the effects of cannabinoid receptors must be obtained using the much more potent antagonists or inverse agonists to block the action of a particular class. These classifications were made based on the descriptions given in Pertwee et al. 2006.
Manipulation of the ECS can be performed using direct agonists or antagonists of CB receptors and concentrations of endogenous ligands can be altered by pharmacologically interfering with enzymatic degradation pathways [8,12-15,33]. Phytocannabinoids that act at CB receptors include Δ9-THC, Δ8-THC, cannabinol, and cannabidiol with the former facilitating the primary psychoactive effects of acute cannabis ingestion and the latter maintaining little psychoactive capacity. These natural ligands are not known for their high selectivity and only with the advent of synthetic compounds has localization been clearly performed [6,8-11]. An example of this abundant cross selectivity is the transient receptor potential vanilloid 1 receptors (TRPV1), which is activated by many of the same molecules that trigger CB receptor activation [34,35]. This indicates the presence of a highly complex lipid signaling system that is dependent upon the local composition of lipids in the cellular membrane, along with dependence on the local receptor expression profile. Together, these two variables interact to produce an averaged CB signal that can have dramatic effects on synaptic responses to incoming depolarization. As a result, high endocannabinoid densities can be found at synapses after periods of sustained cellular firing [36]. These lipids in the cellular membrane act on CB1 receptors and produce a physical constant that regulates potassium and calcium currents through the synapse [15,25,32,37-39]. This inhibitory regulation of synaptic current explains the effects of strong CB agonists where application can result in reduced motor coordination, mood elevation, memory storage deficits, and increased appetite [15,39-42]. With this effect on appetite in mind, blockade of the CB1 receptor has been investigated as a method for treating obesity. Models in mice have shown lean body phenotype is associated with high levels of endogenous cannabinoids. Also, weight loss among obese rats with administration of CB1 antagonists has been observed [43]. The CB1 antagonist SR141716A (Rimonabant) was developed for this purpose in humans. Once released on the market, patients experienced increases in anxiety, depression, and suicidal ideation, likely due to the distribution of CB1 receptors in the basal ganglia. After considering the effects of this antagonist, it was removed from the market even though the drug was effective at achieving reductions in weight [44].
The ECS provides an optimal target for therapeutic intervention regarding mental health disorders. The ubiquitous expression of CB proteins includes many of the key systems targeted by existing therapies, such as selective serotonin reuptake inhibitors (SSRIs) and serotoninnorepinpehrine reuptake inhibitors (SNRIs) [6,13,15-18,45-55]. This is illustrated by the fact that Δ9-THC, when applied alone and in moderate doses, produces an antidepressant effect on human subjects [41,56]. Synthetic encapsulated Δ9-THC is available as a schedule 3 drug in the United States known as Dronabinol for the treatment of nausea, vomiting and other symptoms associated with chemotherapy treatment [57]. The recent discovery of neural CB2 receptors suggests this family of receptors is still largely unexplored in the brain and further inquiry should be conducted to identify possible new therapeutic targets [27-28,58]. Neurophysiological characterization of other receptor systems has demonstrated how differences in receptor density and localization provide key insight into how that specific neurotransmitter system participates in the general process of cognition. Further exploratory studies investigating the interactions between CB1 and CB2 are required to elucidate how this system functions at a molecular and cellular level.
Affective disorders
One of the most prolific classifications of mental health disorders are those that disrupt processing of emotional stimuli, known as affective or mood disorders [1,2]. Within this classification, we find smaller groups with distinct symptoms including bi-polar disorder (BP), Major Depressive Disorder (MDD), dysthymic disorder, and cyclothymic disorder. Approximately 9.5% of American adults aged 18- 44 have experienced an affective disorder from one of these categories in the past 12 months; BP and MDD comprise the majority of cases and will be the focus of this discussion [1]. The neuroanatomical substrates underlying the expression of affective disorders have been the subject of great debate within the mental health community. There are large bodies of evidence pointing toward improper storage and excessive focus on negative memories as a key pathological feature of depression [1-2,59-63]. Patients suffering from affective disorders often have decreased neuronal activity in the dorsal frontal cortex as well as dysfunctional activity with cell loss observed in the hippocampus and amygdala [1,60,64]. To counter pathological activity in these brain regions, modern antidepressant drugs have been designed to target specific proteins involved in the uptake of serotonin and norepinephrine [1-5]. This eventually leads to an increase in the basal levels of these neurotransmitters found in the synapses of neurons across the brain. Patients experience elevated dopamine release due to these alterations in basal serotonin and norepinephrine levels and generally experience elevated mood as a result. Although these pharmacological interventions are effective, especially when coupled with behavioral treatments, there remains a large population that does not respond to conventional treatment alone [1,2,4,5]. This treatmentresistant group could benefit from altering the basal norepinephrine and serotonin levels using a different intermediary than conventional drugs. Manipulation of the ECS could serve as a means for providing this therapeutic modulation of the neural substrates underlying the disease pathology.
Major depressive disorder (MDD)
Diagnosis of MDD generally occurs after the administration of a variety of tests from the DSM-IV to determine whether there is a disruption in the processing of emotional cues. Tests are routinely performed by a psychologist or psychiatrist, with additional observation of symptoms such as fatigue, lowered mood, suicidal ideation, lack of appetite, and disruption of normal sleeping patterns. Broad fluctuations in symptomology with this disorder can lead to difficulties in diagnosis, as patients can sometimes be hesitant to initially consult a specialized physician concerning their condition [1]. This may be observed by the increasing number of patients diagnosed with MDD by their family or general practitioner instead of a mental health professional. Conventional treatments for MDD modulate serotonin and norepinephrine, increasing dopaminergic tone and generally resulting in relief of depressive symptoms [1,2].
Bipolar 1 and 2 disorders
Patients suffering from bipolar disorder, formerly referred to as manic-depression, experience symptoms characterized by cyclic fluctuations between periods of manic happiness and deep depression [1,61,62]. About 2.6% of American adults have been diagnosed or were bipolar within the past twelve months [1]. The large changes in mood are often disruptive to relationships and can interfere with a patient’s ability to maintain steady work. The distinction between the two forms of the disorder is primarily determined by examining the extent of the manic episodes, as bipolar 2 patients generally do not manifest with as potent manic symptoms. Those suffering from bipolar 2 disorders can often be confused with a person suffering intermittent depression and diagnosis must be performed with respect to long term symptomology. Lithium is one of the most common mood stabilizers and has gained popularity in treating bipolar disorder even though it has a very small therapeutic window [1,2,61,62].
The ECS and affective disorders
Investigating the ECS could enhance our ability to treat mental health disorders in the future by providing alternative molecular routes for modulating serotonin and norepinephrine. For example, CB1 receptors have been identified in the raphe nucleus of the midbrain, which is a major nucleus containing noradrenergic cell groups [53,54]. These noradrenergic cells project axons throughout the brain to provide modulation of glutamate and GABA at specific sites including cells of the amygdala, striatum, and forebrain. In models of depression, there are deficits in noradrenergic signaling that ultimately result in decreased dopaminergic tone and can result in negatively biased mood [60]. In addition, there are abundant CB1 receptors expressed in the amygdala, a major neural substrate of emotional processing, as well as the hippocampus, which participates in memory storage and retrieval [20,21,41,55,65-70]. This indicates that CB receptors are likely involved in the emotional coding of memories and could potentially present a method for modulating aberrant ideation on negative emotions. To this effect, mice overexpressing CB2 receptors have a phenotype that leaves them less vulnerable to negative stressors [71]. In contrast, human studies have shown that individuals with certain variations of the CB1 gene have a higher potential for being resistant to pharmacological treatment for depression and therefore could be more at risk if depression occurs [69]. Taken together, it is clear that the ECS is involved in the processing of emotional memories and could therefore provide therapies to counter these behaviors [72].
In addition to its action with pharmacological plasticity, the CB1 receptor has recently been implicated in the neurogenesis of inhibitory neurons within the hippocampus [73-76]. This finding is significant because perturbations in memory formation are observed in affective disorders along with deficits in hippocampal cell growth. Conventional therapies that targets serotonergic systems have also been shown to induce neurogenesis, which has led to the development of the neurogenesis hypothesis regarding depression [64,70,77]. This extends the role of cannabinoids in memory beyond molecular plasticity to a new level that includes developmental plasticity in response to environmental interactions. This evidence suggests that the ECS could be involved in regulating emotional memory and this may explain why we see antidepressant effects with THC administration.
Schizophrenia and psychotic disorders
Approximately 1.1% of the US adult population currently suffers from a psychotic disorder of some kind, which is relatively low when compared with other mental health disorders [1,78]. The chronic, severe nature of the symptoms associated with this disorder makes it particularly challenging to treat and can have drastic effects on a patient’s long-term health. The manifestation of psychosis varies based on the individual, but generally produces behavioral, emotional, and cognitive symptoms often characterized by somatosensory hallucinations, inabilities to execute planning, and erratic emotional patterns [78-80]. Three major classes of symptoms called positive, negative and cognitive have been developed to describe the range of symptoms in schizophrenia. The positive symptoms reported by patients include hallucinations and delusions that can affect all motor and sensory systems with varying levels of intensity. Negative symptoms incorporate those that have to do with disruption of the patient’s emotional state, similar to that observed within affective disorders and sometimes masking the diagnosis [1,78,81]. Lastly, the cognitive symptoms of schizophrenia can be subtle, with slight fluctuations that can often only be detected when specialized tests are performed. Among these categories, there is a large degree of variation in expression of the disorder that is likely due to subtle variations in underlying pathological features [1,78,80]. In the face of this variation, some progress has been made in recognizing common elements between individuals with evidence pointing toward deficits in limbic dopamine signaling and frontal cortical hypo-activity contributing to the disorder [1,78-82].
The ECS and schizophrenia
The involvement of the ECS in the emergence of schizophrenia has been highly debated and there is little controlled research involving human subjects to adequately infer therapeutic relationships. There is evidence to suggest that cannabis use can improve cognitive function in certain patients with schizophrenia [83-85]. This concept is contrary to much of the evidence in the schizophrenia research community, where many report cannabis as the culprit behind the emergence of psychosis [86-90]. At most, it has been shown that cannabis abuse can exacerbate psychotic tendencies in those that are already predisposed to schizophrenia. Most of the research that proposes a negative link between cannabis and schizophrenia relies upon correlations made by interviewing populations suffering from schizophrenia that have used cannabis recreationally either in the past or currently [83,86-90]. The wide array of patients used in these studies often confounds the links between variables of interest suggesting these studies need to be viewed with more scrutiny. Since there are few adequate mouse models that can fully replicate the symptoms of schizophrenia, our progression towards understanding the underlying neural physiology of schizophrenia has been hindered [91]. The few models that do exist in rodents have shown that agonism of the ECS can reduce the severity of the negative symptoms of schizophrenia, but has little effect on the positive symptoms of the disorder [92-93]. Black et al. [93] suggest that antagonism of CB1 with co-administration of anti-psychotics improves cognitive function and reduces the side-effect profiles of anti-psychotics. An example of this is observed in schizophrenia patients who use cannabis and also display higher levels of success in measures of psychomotor speed when compared with those who do not use cannabis [83,94,95]. As discussed prior, evidence suggests the pathophysiology of schizophrenia may be linked to deficits in limbic dopamine signaling. Thus far, CB receptors 1 and 2 have been reported in high density in limbic and para-limbic areas and are involved in dopamine signaling, suggesting a possible link between dysfunction of the ECS and schizophrenia and highlighting the need for further investigation into this system for possible therapeutic manipulation.
Anxiety/fear disorder
Anxiety is a psychophysiological state characterized by somatic, emotional, cognitive, and behavioral components, which affects roughly 40 million American adults above 18 years of age [1,2,96]. In a clinical setting, humans are diagnosed with anxiety disorders through the professional administration of tests from the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV) [97]. Based upon certain criteria, clinicians can separate the observed behaviors into a variety of subtypes of anxiety including Generalized Anxiety Disorder (GAD), Panic Disorder, Post Traumatic Stress Disorder (PTSD), Obsessive Compulsive Disorder (OCD), or a Phobia of some kind. While anxiety to some degree can be seen as relevant for survival, its persistence beyond situational arousal is generally considered pathological [98].
The etiology of anxiety disorders is unclear. It is known changes occur to the primary modulators of synaptic transmission including shifts in serotonin, nor-epinephrine, and dopamine release [99-101]. These same neuromodulators are associated with depression and many hypothesize these disorders are intertwined, with treatments for both disorders overlapping heavily [67,102]. For example, studies of mood in humans and rodents have shown severe or long-lasting stress can change the anatomical distribution of neuromodulators in the brain and therefore affect behavior [66,67]. Other investigations have shown individuals with certain anxiety disorders possess variations in neuroanatomical structures identified to play a crucial role in controlling memory and mood [103,104]. Moreover, environmental factors, such as trauma or a major life-altering event, could trigger the emergence of anxious behavior in those who have an inherited susceptibility [105]. Treatment for anxiety usually consists of combined behavioral therapy with pharmacological manipulation using classical drugs including benzodiazepines, buspirone, selective serotonin or norepinephrine reuptake inhibitors, and older tricyclic antidepressants [102]. These options have had limited success as their efficacy is highly patient dependant and the side effects are often intolerable, leading to discontinuation of therapy. This suggests pharmacotherapies with better tolerability and broader efficacy must be developed, which is where the ECS is poised to play a crucial role [106-108].
The ECS and anxiety
Cannabinoid receptors have been identified in many regions that are associated with anxiety, such as the amygdala, hippocampus, ventro-medial forebrain, and periaqueductal grey [109,110]. However, interpretation of this localization and its effects on anxiety in rodents has been difficult. Many studies of the effects of cannabinoids on anxiety in mammals show varied outcomes, which can often be attributed to differences in dose, drug, or experimental design. There have been major attempts to parallel classifications of behavior in humans to models of anxiety in rodents with limited success based upon the paradigm and behavior of interest. Measures of time spent in open versus enclosed environments, such as the elevated plus maze, have been used to great effect in the study of anxiety in rodents [111]. This method is based on the logic that rodents have evolved in an environment where enclosed spaces are relatively safe and anxiety free, and the open spaces represent the opposite. Therefore, when an animal is placed in such a maze, the time spent out in the open and time spent in enclosed spaces can be reported as a ratio representing the anxiety level of the animal. Even though great attempts to standardize these methods have been made, interpretation of rodent behavior is still somewhat subjective. Clearly this situation requires remediation and further study should be dedicated to focusing the lens of experimentation on standardized models of anxiety in rodents.
There are several means of altering tone within the ECS, including direct agonism, direct antagonism, and blocking enzymatic pathways of cannabinoid degradation to increase endogenous levels. Studies using direct agonists of the ECS including Delta -9- THC, WIN55-212- 2, anandamide, and others have shown that CB1 is responsible for the primary psychoactive effects associated with cannabis ingestion. The nature of the behavioral effects of these drugs on anxiety is biphasic, with low doses producing anxiolytic effects and anxiogenic effects emerging when high doses are administered [112-115]. In a mouse model of anxiety, the behavioral traits associated with anxiety can be ameliorated by injection of CB1 agonist into the dorsal hippocampus. Additionally, blockade of CB1 results in an anxiogenic response when CB1 antagonist AM251 is directly injected into the core of the amygdala or when it is administered systemically. The synthetic compound URB597 is an inhibitor of FAAH and when administered with anandamide can produce anxiety, but if either is given alone they produce anxiolytic effects [113]. Since CB1 is psychoactive, its use to treat anxiety is clouded due to its side effects, such as euphoria during the initial stages of blood plasma saturation. To this end, CB2 has been investigated as a potential anxiolytic target since it does not possess the psychoactive properties associated with CB1 activation. Studies of CB2 and behavior have shown rodents with higher CB2 expression are resistant to anxiety and have differential responses to benzodiazepines, likely due to changes in GABAergic tone. Studies of this receptor and its relationship to anxious behavior are still limited and must be further explored before any strong conclusions can be drawn. Since there is evidence the ECS interacts with many different modulatory systems, including current anxiety treatments, exploration of this receptor family could open a new taxonomy of lipid-like drugs [116].
The ubiquitous expression of CB receptors on cells in the brain presents a wide array of possibilities for altering neuronal activity and potentially treating pathological activity [8,24,54,71]. Given the complexity of the ECS, further studies must be conducted to establish a baseline for comparison to models of disease. Genetic knock-out mice have been one of the most supportive tools in recent efforts to understand the cannabinoid system as they lack CB receptor expression. Behavioral assessment of the mice under application of various cannabinoids will allow the assessment of specific pharmacological activity for various ligands. These results can then be translated into human trials used to formally test the potential of the compound. Currently, no clinical trials can be performed using herbal derivatives of cannabis even though the material likely contains potent modulators of psychological state. Future research aimed at isolating specific herbal ligands for behavioral and pharmacological evaluation in animal models is the next step. In addition, combinations of cannabinoids should be studied together due to the moderate selectivity of many ligands. It is likely that what is observed with cannabis is due to the mixture of cannabinoids found within the plant itself and not due to specific ligands such a THC alone [8]. The phytocannabinoids within cannabis have already undergone evolutionary development through coexistence with mammals and are likely an important reservoir of information for the understanding and modulation of our endogenous neurological systems.
The lipophilic nature of cannabinoid ligands confers them an inherent advantage over other molecules because they are able to traverse the blood brain barrier (BBB) and incorporate themselves into the cell membrane. Embedded within and free to move along the plane of the plasma membrane, cannabinoid ligands probabilistically interact with CB receptors. The probability of CB receptor activation scales with the density of ligands in the membrane is a complex value due to the partial activity of many molecules at these receptors [42]. This suggests that drugs designed to target cannabinoid receptors will have a longer half-life and also have a high therapeutic index as the hydrophobic nature of the molecule reduces the chances of overdose. Properties such as these represent key advantages of cannabinoids that could be beneficial in scenarios where current treatments carry risks of abuse and non-compliance, such as in bipolar disorder or schizophrenia.
The emergence of incredibly powerful genetic and molecular tools, including fluorescent probes, has provided the tools to study the very foundations of mental health in our own cells. To truly understand the physiological mechanisms of the ECS, we must look beyond cultural stigmas to envision new medical frontiers wherever they may present. From the modest research conducted thus far, it is apparent the ECS plays a crucial role in normal neurophysiology and in many functions that we consider core elements of our human existence, such as memory and emotion. It is also probable that the ECS is involved in mediating symptoms of mental health disorders, and dysfunction of the ECS may even be involved in the pathogenesis of mental health disorders. With current research so focused on determining the underlying mechanisms and potential therapeutic opportunities of mental health disorders, it is enormously short-sighted to ignore such an integral and robust signaling system. Investigating the ECS, especially in conjunction with other neural signaling systems will likely provide a more comprehensive understanding of normal neurophysiology and pathophysiology, as well as present novel techniques for modulating the symptoms of mental health disorders.
We would like to thank the Gaines Family Foundation for their generous support of this work and continuing efforts to support medical cannabis research.